遠期恐懼記憶再鞏固更新機制的線索選擇性特點*
2019-03-05 02:13:16胡靜初張蔚欣陳小婷王文清王子潔莊楚群鄭希付
心理學報 2019年3期
胡靜初 張蔚欣 陳小婷 王文清 王子潔 莊楚群 馮 彪 鄭希付
?
遠期恐懼記憶再鞏固更新機制的線索選擇性特點
胡靜初張蔚欣陳小婷王文清王子潔莊楚群馮 彪鄭希付
(華南師范大學心理學院;華南師范大學生命科學學院, 廣州 510631) (中山紀念中學, 廣東 中山 528400)
已有動物和人類研究均表明, 通過記憶的再鞏固更新機制能有效削弱新形成的條件性恐懼記憶(1天), 并且存在線索選擇性特點。然而創傷后應激障礙(PTSD)往往在形成相當一段時間后才能得到治療, 且現實生活中人們通常一次習得對多個線索的恐懼。因此找到針對多線索創傷記憶的有效治療方法顯得尤為重要。目前未有人研究遠期恐懼記憶的再鞏固更新機制是否存在線索選擇性特點。為探究遠期恐懼記憶(>7天)的再鞏固更新機制是否同樣存在線索選擇性特點, 本研究采用被試內實驗設計, 以皮膚電作為恐懼反應指標, 多個線索作為條件刺激進行恐懼習得, 習得14天后給被試單獨呈現一個線索進行恐懼記憶提取, 10分鐘后進行消退訓練, 24小時后對不同線索進行自發恢復測試。結果顯示:未提取線索的自發恢復程度顯著高于提取線索。說明遠期記憶(14天)的再鞏固更新機制同樣存在線索選擇性特點, 并確認了提取消退作為一種行為手段對遠期恐懼記憶再鞏固進行干預的有效性, 對臨床干預具有一定指導意義。
遠期記憶再鞏固; 線索選擇性; 條件性恐懼; 提取消退
1 前言
恐懼記憶的形成通常以經典巴甫洛夫恐懼條件化模型為原理, 該模型認為, 一個原本不具備引發個體恐懼情緒的中性刺激CS (條件刺激, Conditioned Stimulus, 如一個聲音), 多次伴隨一個恐怖或者厭惡刺激US (無條件刺激, Unconditioned Stimulus, 如電擊)出現之后, 個體會逐漸習得對這個中性刺激的條件恐懼反應。當這個中性刺激CS單獨出現時, 個體會對其產生恐懼反應(Mineka & Zinbarg, 2006)。同時, 條件性恐懼也可以消退, 當條件刺激繼續出現而恐懼刺激不再出現時, 已經習得的條件恐懼反應則會逐漸消退。在臨床實踐中, 往往使用基于消退訓練原理的暴露療法治療與恐懼有關的情緒障礙(Barlow, Allen, & Choate, 2016)。然而, 消退訓練一段時間后, 恐懼常常會以各種形式返回, 如:自發恢復、續新、重建, 訓練效果不佳。這是因為消退訓練使個體產生了一種新的抑制記憶, 與原來的CS-US恐懼記憶進行競爭, 并未減弱原來的恐懼記憶, 而是產生了一種新的CS-no US的聯結(Bouton, 2002)。
傳統記憶鞏固理論認為, 記憶一旦鞏固形成, 則無法改變。近年來這個觀點受到挑戰, 研究者們認為已鞏固的記憶可以經過提取重新進入不穩定狀態, 經過一段時間后再次鞏固穩定下來, 這個過程稱為記憶的再鞏固(Alberini, 2005; Dudai, 2004; Nader, 2003)。通過記憶的再鞏固更新機制, 可削弱恐懼記憶, 有效阻止恐懼的返回。動物研究中, 在提取激活新形成的恐懼記憶后立刻注射蛋白質合成抑制劑(茴香霉素, anisomycin), 大鼠在之后的測試中表現出恐懼反應顯著減少(Nader, Schafe, & Le, 2000)。隨后Doyère, Debiec, Monfils, Schafe和Ledoux等人(2007)發現, 大鼠新形成的恐懼記憶再鞏固存在線索選擇性特點, 即:運用再鞏固更新機制只能選擇性地削弱提取了的特定線索的恐懼記憶, 并且這種線索選擇性特點是通過提取以選擇性地改變個體杏仁核的突觸可塑性來達成的。由于藥物對人類存在副作用, 研究者們隨后提出了行為干預恐懼記憶再鞏固的提取消退范式, 即在記憶提取后, 在記憶的再鞏固時間窗內進行消退訓練以削弱原始恐懼記憶。Monfils, Cowansage, Klann和Ledoux (2009)首次驗證提取消退范式的有效性, 記憶提取后在再鞏固時間窗內進行消退訓練, 有效地削弱了大鼠的新形成的恐懼記憶, 而Schiller等人(2010)則首次使用提取消退范式成功干預了人類新形成的恐懼記憶再鞏固, 并且證實了人類恐懼記憶再鞏固同樣存在線索選擇性特點。
如果運用記憶再鞏固更新機制能夠削弱恐懼記憶, 這對創傷后應激障礙的治療將有重大意義。然而, 創傷后應激障礙患者很難在創傷記憶形成后立即得到治療, 臨床中需要干預的大多是遠期記憶。遠期恐懼記憶被證明與新形成的恐懼記憶有著不同的特點。一些大鼠的研究表明, 相較于近期記憶, 遠期記憶更不容易受到提取的干擾(Milekic & Alberini, 2002; Suzuki et al., 2004)。Suzuki等人(2004)的結果表明在小鼠習得條件性恐懼反應56天后, 使用茴香霉素干擾記憶再鞏固, 發現未出現藥物對遠期記憶再鞏固的干擾效果。這是由于遠期恐懼記憶與新形成的恐懼記憶有著不同的神經機制, 新的恐懼記憶的編碼和存儲更依賴于海馬, 而遠期記憶的存儲更依賴于新皮層(Anagnostaras, Maren, & Fanselow, 1999; Eichenbaum, Otto, & Cohen, 1994; Frankland, O'Brien, Ohno, Kirkwood, & Silva, 2001; Kim & Fanselow, 1992; LeDoux, 1999; Mcclelland, Mcnaughton, & O'Reilly, 1995; McGaugh, 2000; Quevedo et al., 1999; Squire & Alvarez, 1995; Rie, Hotaka, Frankland, & Satoshi, 2016), 因此, 提取激活遠期記憶需要激活皮層區域, 而不僅是海馬。由于大腦皮層區域廣泛, 所以更難達到激活(Bontempi, Laurent-Demir, Destrade, & Jaffard, 1999; McClelland et al., 1995)。
目前, 大部分人類恐懼記憶再鞏固研究都集中在新形成(1天)的恐懼記憶, 沒有人研究過大于7天的恐懼記憶。然而真實生活中的創傷記憶往往是遠期恐懼記憶, 而且常伴隨多個習得線索。因此, 多線索遠期恐懼記憶再鞏固是否可以被成功干預, 以及是否同樣存在線索選擇性特點, 成為了關鍵問題。
為探究遠期恐懼記憶再鞏固更新機制是否存在線索選擇性特點, 以及再鞏固更新機制是否對人類遠期恐懼記憶同樣有效, 我們使用Schiller等人(2010)的提取消退范式, 被試在第一天習得恐懼記憶, 14天后進行提取, 提取后在再鞏固時間窗內進行消退訓練, 后一天進行自發恢復測試, 探究14天的條件性恐懼記憶是否能被干預, 以及干預是否存在線索選擇性特點, 以推進記憶再鞏固更新機制在心理治療中的臨床應用。
2 對象與方法
2.1 被試
被試為28名在校大學生(其中男生15名, 女生13名), 通過自愿報名的方式參加, 年齡范圍為18~23歲(= 23.34,= 1.18)。對于本研究所涉及到的被試采用如下選擇方法:在大學里發布招募通知, 在報名的大學生中先通過電話詢問其是否有精神障礙史、最近是否有服用過精神類藥品等問題, 選擇右利手、無軀體疾病和精神障礙、視力或矯正視力正常、無色盲及色弱且之前沒有參加過類似實驗的被試。
正式實驗前首先向被試說明:本實驗是一個恐懼情緒實驗, 實驗過程中將在左手食指和無名指的末端指腹上連接電極以記錄皮膚電, 在右手手腕處施加一個輕微電擊, 由于每個人的感覺閾限有所不同, 正式實驗前首先會進行電擊強度的評定, 電擊儀器的使用是經過科學評定的, 絕不會對您造成任何傷害, 如果在實驗過程中有任何不適, 可隨時提出終止實驗, 三天實驗結束后給您支付相應的報酬。接著, 被試需簽署實驗知情同意書, 并填寫《狀態?特質焦慮量表》。被試狀態焦慮(= 38.21,= 1.32)和特質焦慮(= 38.90,= 2.29)均在正常范圍之內。
在實驗設備準備好后, 進一步確認被試是否能看清電腦屏幕, 能否聽清實驗員的指導語等, 在得到確認答復后才開始正式實驗。
一個樂于“嚼舌根”的人,常常是一個成功度不高、渴望友誼但又難獲真正友誼的人。人活著,總需要一點存在感。有些人為了證明存在的價值,完全不顧他人感受,像鐘女士就是這樣。
2.2 實驗刺激
條件刺激(CS):采用圖片作為條件刺激, 圖片分別為黃色正方形、藍色正方形和綠色的正方形。其中兩種顏色的正方形會跟隨US, 作為CSa+、CSb+, 另一種顏色的正方形始終不會跟隨US, 作為CS? (如圖1所示)。為避免被試對不同顏色的偏好和順序效應, 刺激顏色和呈現順序已進行平衡。三個圖片均為標準色, 具有相同的大小和亮度, 呈現時間為4 s, 背景為標準白色。為平衡不同顏色的正方形在不同被試間的影響, 三種顏色的圖形有相同的概率充當CS+。

圖1 遠期恐懼記憶再鞏固實驗的刺激材料
非條件刺激(US):參考Schiller等人(2010)的研究, 本實驗采用腕部電擊作為非條件刺激(US)來誘發被試的恐懼情緒, 電擊持續時間為200 ms。考慮到個體之間感覺閾限會存在差異, 在正式實驗前會讓被試對電擊強度進行評定, 被試需要自行決定讓其感到極度不舒服但可以忍受的電擊強度。
2.3 實驗流程
電擊強度評定:在正式實驗之前先對每個被試所能接受的電擊強度進行評定, 這個強度能讓被試感到極度不舒服但可以忍受。評定先從較微弱的10 V開始, 之后每次增大10 V, 并在每次增大電擊強度之后詢問被試的感受, 直到被試報告強度已達到讓其感到極度不舒服但可以忍受的程度(電擊強度最高不超50 V)。每次電擊持續時間為200 ms, 電流為每秒50脈沖。在接下來的各個實驗環節中被試的電擊強度不再改變。
實驗分為3天, 分別為第1天的習得階段、第15天的提取?消退階段、第16天的自發恢復測試階段(如圖2所示)。正式實驗開始前, 先向被試詳細解釋指導語, 確保被試明白指導語的意思后, 進入第一天的習得階段。
第1天習得階段:CSa+、CSb+分別呈現13次(每個CS不會超過兩次連續出現), 每次呈現4 s, 試次之間的時間間隔(ITI)為12~18 s, 其中有38% (5次) CSa+、38% (5次) CSb+伴隨電擊。CS?呈現8次, 均不伴隨電擊。
第15天提取消退階段:先給被試單獨呈現一次CSa+, 呈現時間為4 s, 以提取激活習得的恐懼記憶。接著被試休息10 min, 休息之后進行消退訓練。消退過程中CSa+、CSb+和CS?各隨機呈現11次, 共呈現33次, 每次呈現4 s, ITI為12~18 s, 均不跟隨電擊。
第16天自發恢復測試階段:自發恢復測試過程CSa+、CSb+、CS?各隨機呈現11次, 共呈現33次, 每次呈現4 s, 均不跟隨電擊, ITI為12~18 s。
所有實驗刺激都通過Eprime 2.0 編程呈現。首先在屏幕中央呈現紅色注視點“+”, 呈現時間為1000 ms, 注視點后呈現條件刺激CS, 呈現時間為4 s, 隨后跟隨非條件刺激US或者白屏, 呈現時間均為200 ms (如圖3所示)。實驗過程中被試不需要做任何按鍵反應。
2.4 測量指標
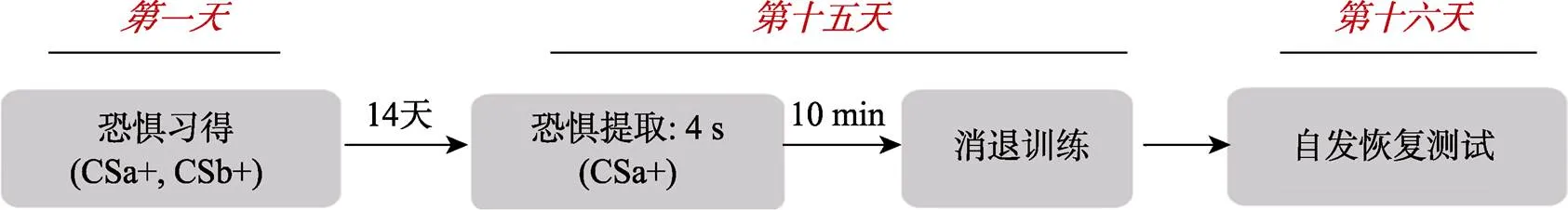
圖2 實驗流程
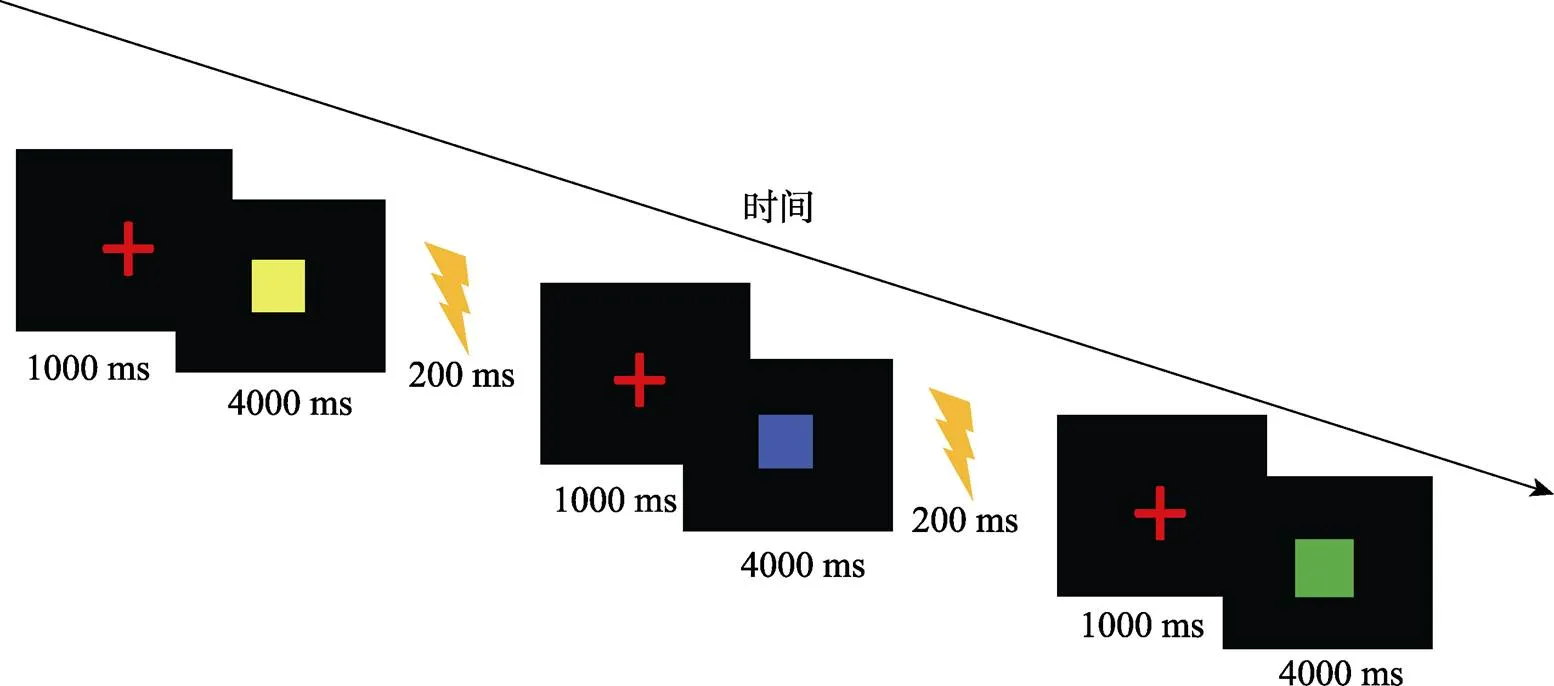
圖3 實驗刺激呈現示意圖
本實驗的數據均使用SPSS 17.0進行統計分析。分別對各個階段(恐懼學習階段, 恐懼消退階段, 恐懼自發恢復階段)的皮膚電進行刺激類型(CSa+, CSb+, CS?) × 階段(早期, 晚期)的重復方差測量分析, 并采用Bonferroni法進行事后檢驗或配對樣本t檢驗對不同階段的不同刺激類型的差異進行比較。
3 實驗結果
3.1 習得階段
以刺激類型(CSa+, CSb+, CS?)和實驗階段(習得早期:習得階段前4個試次; 習得晚期:習得階段后4個試次)為被試內因素做重復測量方差分析, 結果顯示:刺激類型的主效應顯著,(2, 56) = 9.79,< 0.001, η2 p= 0.26; 實驗階段的主效應顯著,(1, 28) = 18.70,< 0.001, η2 p= 0.40; 刺激類型和實驗階段的交互作用顯著,(2, 56) = 3.89,< 0.05, η2 p= 0.12。
采用Bonferroni法進行事后檢驗發現CSa+, CSb+, CS?在習得早期均不存在顯著差異(1.00,0.45,0.37), 而到了習得晚期, CSa+與CS?, CSb+與CS?出現了顯著差異(0.001,< 0.01), 而CSa+與CSb+仍然不存在顯著差異(0.30)。
分別對習得晚期的皮電均值進行刺激類型(CSa+, CS?)與(CSb+, CS?)的配對樣本t檢驗, 結果發現在恐懼習得晚期被試對CSa+, CSb+的皮膚電反應均顯著高于CS? [CSa+,(28) = 6.07,< 0.001, Cohen’s= 1.25; CSb+,(28) = 3.95,< 0.001, Cohen’s= 0.83], 而對CSa+和CSb+的皮膚電反應卻不存在顯著差異,(28) = 1.70,= 0.10, Cohen’s= 0.32。
總體來看, 在習得的早期階段, 被試對CSa+, CSb+與CS?的差異并不顯著, 意味著被試在習得早期對三種刺激類型的恐懼程度一致, 但隨著實驗階段進行到習得晚期, 被試對CSa+, CSb+的反應顯著的高于了CS?, 而對CSa+和CSb+的差異仍然不顯著。這意味著隨著實驗的進行, 被試在習得晚期對CSa+和CSb+產生了顯著高于CS?的恐懼, 并且對CSa+和CSb+的恐懼程度仍然趨向一致(如圖4、圖5所示)。
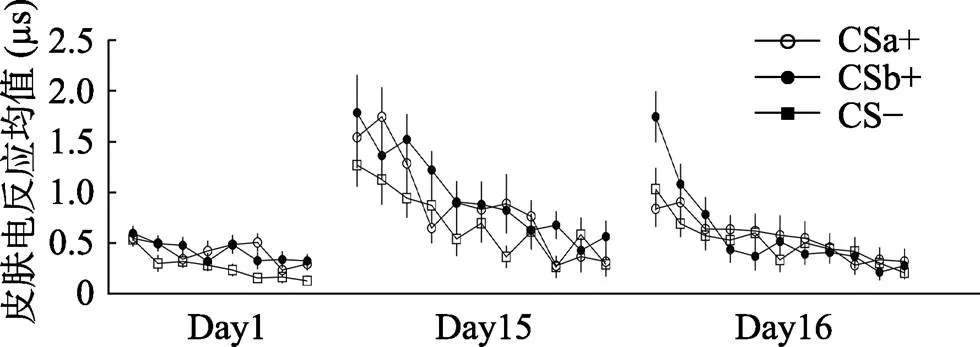
圖4 各階段不同刺激類型的皮電反應值變化
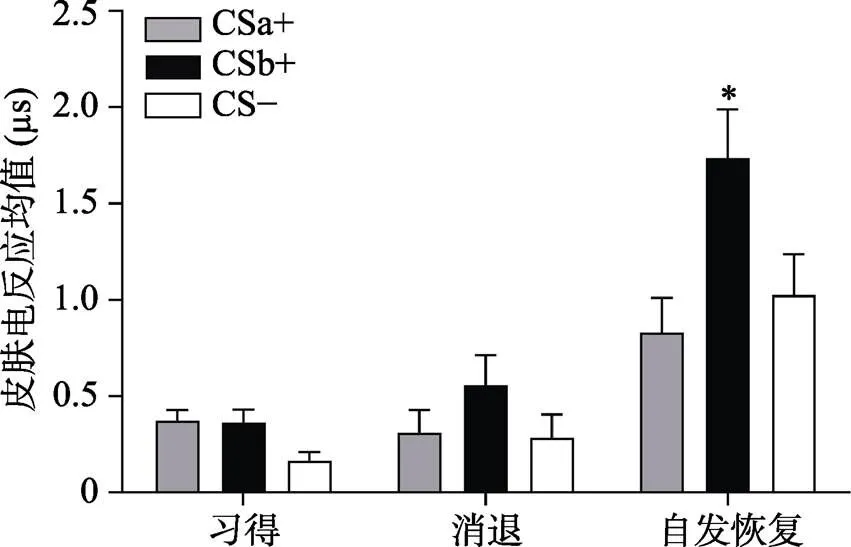
圖5 各階段不同刺激類型的皮電反應均值
注:習得階段(Acquisition)為習得后半段均值(后5個試次的均值); 消退(Extinction)為消退最后一個試次均值, 自發恢復(Recovery)為自發恢復第一個試次均值。
3.2 消退階段
以刺激類型(CSa+, CSb+, CS?)和實驗階段(消退早期:消退階段前4個試次; 消退晚期:消退階段后4個試次)為被試內因素做重復測量方差分析, 結果顯示:刺激類型的主效應顯著,(2, 56) = 6.50,< 0.01, η2 p= 0.19; 實驗階段主效應顯著,(1, 28) = 25.56,< 0.001, η2 p= 0.48; 刺激類型與實驗階段的交互作用顯著,(2, 56) = 4.16,< 0.05, η2 p= 0.13。
為了檢測消退訓練的效果, 我們分別計算了消退前三個和最后三個試次中呈現不同刺激類型(CSa+, CSb+, CS?)時的皮電均值, 并對它們進行了刺激類型(CSa+, CS?; CSb+, CS?)的配對樣本t檢驗, 結果發現被試在消退的前三個試次對CSa+與CSb+的皮電反應顯著高于CS? [CSa+,(28) = 2.94,< 0.01, Cohen’s= ?0.05; CSb+,(28) = 3.48,< 0.01, Cohen’s= 0.78], 而到了最后三個試次對CSa+, CSb+的反應與CS?無顯著差異[CSa+,(28) = ?0.69,= 0.50, Cohen’s= ?0.13; CSb+,(28) = 1.63,= 0.12, Cohen’s= 0.31]。
對消退階段最后一個試次的皮電均值進行刺激類型(CSa+, CS?; CSb+, CS?)的配對樣本t檢驗, 結果發現被試在消退的最后一個試次對CSa+, CSb+的反應與CS?無顯著差異[CSa+,(28) = 0.17,= 0.87, Cohen’s= 0.03; CSb+,(28) = 0.99,= 0.33, Cohen’s= 0.19]。這表示在消退的末尾階段, CSa+、CSb+的皮電值與CS?沒有顯著差異, 表明被試完成了消退訓練。并且配對t檢驗顯示被試在消退的最后一個試次對CSa+與CSb+沒有顯著差異,(28) = ?0.89,= 0.38, Cohen’s= 0.17, 這意味著被試對CSa+與CSb+的恐懼消退程度一致(如圖4、圖5所示)。
3.3 自發恢復測試階段
以刺激類型(CSa+, CSb+, CS?)和實驗階段(自發恢復早期:自發恢復階段前4個試次; 自發恢復晚期:自發恢復階段后4個試次)為被試內因素做重復測量方差分析, 結果顯示:刺激類型的主效應顯著,(2, 56) = 3.17,< 0.05, η2 p= 0.10; 實驗階段的主效應顯著,(1, 28) = 26.43,< 0.001, η2 p= 0.49; 刺激類型和實驗階段的交互作用顯著,(2, 56) = 4.69,< 0.05, η2 p= 0.14。
采用Bonferroni法進行事后檢驗發現, 在自發恢復早期階段, CSb+的皮電值顯著大于CSa+ (< 0.05), 并且顯著大于CS? (< 0.01), 而CSa+與CS?的皮電值不存在顯著差異(= 1.00), 而到了自發恢復晚期, CSa+與CS?, CSb+與CS?, CSa+與CSb+的差異均不顯著(s = 1.00)。這表明, 在自發恢復的早期階段, 被試對沒有經過提取后消退的CSb+出現了明顯的自發恢復反應, 卻沒有對經過提取消退訓練的CSa+表現出恐懼的自發恢復。而隨著實驗進行到后半段, 由于一直沒有電擊US的出現, 被試一開始對CSb+出現的恐懼自發恢復逐漸被消退, 對CSa+, CSb+, CS?的反應逐漸趨于一致(見圖4)。
為了進一步探究不同刺激類型從消退階段到自發恢復階段的變化情況, 我們對CSa+與CS?的差值, CSb+與CS?的差值進行了兩因素重復測量方差分析, 以與CS?差值的刺激類型(CSa+, CSb+)和實驗階段(消退最后一個試次; 自發恢復第一個試次)為因素, 結果顯示:刺激類型的主效應顯著,(1, 28) = 10.15,< 0.01, η2 p= 0.27; 實驗階段的主效應不顯著,(1, 28) = 0.52,= 0.48; 刺激類型和實驗階段的交互作用顯著,(1, 28) = 5.63,< 0.05, η2 p= 0.17。
通過配對樣本t檢驗對CSa+/CSb+與CS?在自發恢復測試階段第一個試次的皮電差值和在消退階段最后一個試次的皮電差值進行比較, 結果顯示:即CSb+與CS?在自發恢復測試階段第一個試次的皮電差值顯著高于在消退階段最后一個試次的皮電差值:(28) = ?2.07,< 0.05, Cohen’s= 0.15, 而CSa+與CS?沒有差異:(28) = 0.78,= 0.44, Cohen’s= ?0.39 (如圖6所示), 說明提取了的CSa+沒有出現自發恢復, 而沒有提取的CSb+出現了顯著的自發恢復。
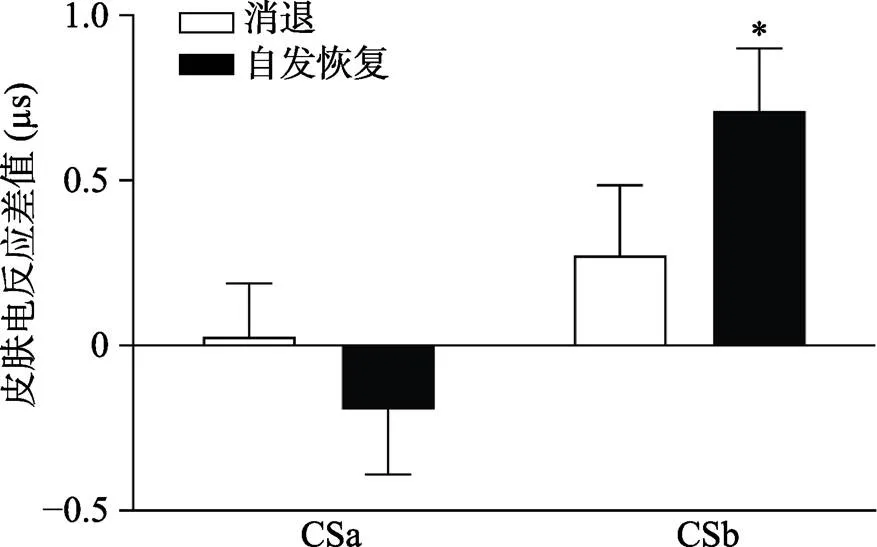
圖6 CSa+/CSb+與CS?在自發恢復測試階段第一個試次和消退階段最后一個試次的皮電差值
注:*< 0.05 CSb+與CS?在自發恢復測試階段第一個試次的皮電差值與在消退階段最后一個試次的皮電差值存在顯著差異。
4 討論
本研究探究了提取消退訓練是否能夠干預人類遠期恐懼記憶再鞏固, 以及是否存在線索選擇性特點。被試在第1天習得恐懼, 第15天進行恐懼記憶的提取及消退訓練, 第16天進行自發恢復測試。實驗結果表明, 提取消退訓練成功干預了遠期恐懼記憶再鞏固, 阻止了遠期恐懼的自發恢復, 而且同樣存在線索選擇性特點。
遠期恐懼記憶的再鞏固更新機制出現線索選擇性特點, 可能是由于提取單一線索這一操作, 只能選擇性地再激活大腦皮層中對于特定線索的遠期恐懼記憶, 讓特定記憶進入不穩定狀態, 導致了只有特定線索的記憶得以被干預和破壞。這一結果也意味著不同線索的恐懼記憶可能是以不同路徑編碼儲存在特定大腦皮層, 印證了人類記憶編碼儲存方式的復雜度。
本研究的實驗結果與部分使用藥物干預遠期恐懼記憶再鞏固的動物實驗結果一致(Debiec, Ledoux, & Nader, 2002; Nader et al., 2000), 也與Steinfurth等人(2014)的人類實驗結果一致, 他們嘗試使用提取消退范式對7天的恐懼記憶再鞏固進行干預, 并成功阻止了人類遠期恐懼的返回。我們的結論驗證了記憶的不穩定性:無論是剛形成的新記憶還是遠期記憶, 一旦有新的安全信息進入再鞏固時間窗, 記憶就非常容易被改變。
值得注意的是, 本研究與部分使用提取消退范式干預遠期恐懼記憶的動物實驗結果不一致。例如, Clem和Huganir (2010)使用提取消退的手段干預小鼠的遠期記憶(7天), 結果發現無法阻止小鼠的遠期記憶返回, 這可能由于恐懼記憶的強度不同, 小鼠在實驗室習得的恐懼記憶相對更強。出于倫理考慮, 我們無法在實驗室創造過強的人類恐懼記憶, 這就決定了實驗室中造成的恐懼記憶可能更加容易被更改。其次, 雖然記憶鞏固周期的分子機制在不同物種間也非常相似, 但他們的記憶時間長度卻可能因為壽命的長短不同而不一致, 如:人類壽命約為70歲左右, 而小鼠壽命約為2年左右, 那么對于小鼠而言, 14天的遠期恐懼記憶可能相當于它們490天的恐懼記憶(Dutta & Sengupta, 2016; Quinn, 2005)。
本研究首次在人類被試上使用提取消退范式干預14天的遠期記憶再鞏固, 并驗證了遠期恐懼記憶再鞏固更新機制存在線索選擇性特點。對于人類而言, 即使是14天的遠期記憶, 也可以通過提取激活進行干預改變。本研究有助于我們進一步了解人類恐懼記憶的特點, 從而有助于我們更好地將基礎研究中的結論推廣到臨床應用中去, 例如根據新形成的創傷記憶與形成很久的創傷記憶的特點及兩者差異, 有針對性地對其進行干預。通過本研究我們也可以了解到人類與動物物種的差異, 如對于人類來說, 可以通過短時間對記憶線索的暴露迅速提取特定恐懼記憶, 哪怕是遠期恐懼記憶也存在同樣規律。本研究還為使用提取消退干預創傷記憶提供了指導, 具有一定的臨床應用價值, 因為大多數實驗室創造的條件性恐懼記憶多是在一天后進行干預, 而真實生活中的創傷記憶往往不可能在剛形成時就能得到干預。
然而, 本研究同樣存在幾個限制:第一, 在最后的實驗結果中, 由于習得和消退恐懼是本實驗得以進行的先決條件, 我們排除了無法習得或消退恐懼的被試, 因此無法測量無法習得和消退恐懼的被試的自發恢復(Kindt, Soeter, & Vervliet, 2009; Sotres-Bayon, Diaz-Mataix, Bush, & Ledoux, 2009; Yang, Chao, & Lu, 2006); 第二, 我們使用能體現自主神經反應的恐懼指標(皮膚電)來測試人類的條件化恐懼, 然而皮膚電經常隨著室溫和天氣而變化, 被試學習恐懼的第一天和提取消退的第15天的氣溫非常不同, 這也解釋了為什么我們實驗中第一天習得的數據范圍相對于第15天要低的多; 第三, 我們只使用了皮膚電作為測量恐懼的指標, 并沒有使用任何認知外顯指標(如US預期, 表示被試對于每個試次出現電擊的概率估計, 可以代表被試對于條件化恐懼學習的外顯學習程度, 同樣也是恐懼學習的一個有力指標)。然而, 研究表明使用認知指標會導致被試習得更強, 讓被試更能外顯地學習到CS-US之間的知識聯結, 以致于記憶無法消退(Warren et al., 2014), 同時容易與皮膚電指標相互影響。基于這些原因, 我們更傾向于使用一種非侵入性、能反應自主神經活動的指標來測量恐懼程度, 但這也在某種程度上減少了數據的獲得。
本研究證實, 遠期恐懼記憶再鞏固更新機制存在線索選擇性特點, 以及14天的遠期恐懼記憶也是可以通過在提取后的再鞏固時間窗內進行消退訓練而成功削弱的, 再次確認了提取消退作為一種記憶再鞏固的行為干預手段的有效性。
Alberini, C. M. (2005). Mechanisms of memory stabilization: Are consolidation and reconsolidation similar or distinct processes?.(1), 51–56.
Anagnostaras, S. G., Maren, S., & Fanselow, M. S. (1999). Temporally graded retrograde amnesia of contextual fear after hippocampal damage in rats: Within-subjects examination.(3), 1106–1114.
Barlow, D. H., Allen, L. B., & Choate, M. L. (2016). Toward a unified treatment for emotional disorders.(2), 205–230.
Bontempi, B., Laurent-Demir, C., Destrade, C., & Jaffard, R. (1999). Time-dependent reorganization of brain circuitry underlying long-term memory storage.(6745), 671–675.
Bouton, M. E. (2002). Context, ambiguity, and unlearning: Sources of relapse after behavioral extinction.(10), 976–986.
Clem, R. L., & Huganir, R. L. (2010). Calcium-permeable ampa receptor dynamics mediate fear memory erasure.(6007), 1108–1112.
Debiec, J., Ledoux, J. E., & Nader, K. (2002). Cellular and systems reconsolidation in the hippocampus.(3), 527–538.
Dudai, Y. (2004). The neurobiology of consolidations, or, how stable is the engram?(1), 51–86.
Doyère, V., Debiec, J., Monfils, M-H., Schafe, G. E., & Ledoux, J. E. (2007). Synapse-specific reconsolidation of distinct fear memories in the lateral amygdala.(4), 414–416.
Dutta, S., & Sengupta, P. (2016). Men and mice: relating their ages., 244–248.
Eichenbaum, H., Otto, T., & Cohen, N. J. (1994). Two functional components of the hippocampal memory system.(3), 449–581.
Frankland, P. W., O'Brien, C., Ohno, M., Kirkwood, A., & Silva, A. J. (2001). α-camkii-dependent plasticity in the cortex is required for permanent memory.(6835), 309–313.
Kim, J. J., & Fanselow, M. S. (1992). Modality-specific retrograde amnesia of fear.(5057), 675–677.
Kindt, M., Soeter, M., & Vervliet, B. (2009). Beyond extinction: Erasing human fear responses and preventing the return of fear.(3), 256–258.
Ledoux, J. E. (1999). Emotion circuits in the brain.(23), 155–184.
Mcclelland, J. L., Mcnaughton, B. L., & O'Reilly, R. C. (1995). Why there are complementary learning systems in the hippocampus and neocortex: Insights from the successes and failures of connectionist models of learning and memory.,(3), 419–457.
Mcgaugh, J. L. (2000). Memory--A century of consolidation.,(5451), 248–251.
Milekic, M. H., & Alberini, C. M. (2002). Temporally graded requirement for protein synthesis following memory reactivation.(3), 521–525.
Mineka, S., & Zinbarg, R. (2006). A contemporary learning theory perspective on the etiology of anxiety disorders: It's not what you thought it was.(1), 10–26.
Monfils, M. H., Cowansage, K. K., Klann, E., & Ledoux, J. E. (2009). Extinction-reconsolidation boundaries: Key to persistent attenuation of fear memories.(5929), 951–955.
Nader, K. (2003). Memory traces unbound.(2)65–72.
Nader, K., Schafe, G. E., & Le, D. J. (2000). Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval.(6797), 722– 726.
Quevedo, J., Vianna, M. R. M., Roesler, R., De-Paris, F., Izquierdo, I., & Rose, S. P. R. (1999). Two time windows of anisomycin-induced amnesia for inhibitory avoidance training in rats: Protection from amnesia by pretraining but not pre-exposure to the task apparatus.(6), 600–607.
Quinn, R. (2005). Comparing rat's to human's age: How old is my rat in people years?(6), 775–777.
Rie, I., Hotaka, F., Frankland, P. W., & Satoshi, K. (2016). Hippocampal neurogenesis enhancers promote forgetting of remote fear memory after hippocampal reactivation by retrieval.:e17464.
Schiller, D., Monfils, M-H., Raio, C. M., Johnson, D. C., Ledoux, J. E., & Phelps, E. A. (2010). Preventing the return of fear in humans using reconsolidation update mechanisms.(7277), 49–53.
Sotres-Bayon, F., Diaz-Mataix, L., Bush, D. E. A., & Ledoux, J. E. (2009). Dissociable roles for the ventromedial prefrontal cortex and amygdala in fear extinction: Nr2b contribution.(2), 474–482.
Squire, L. R., & Alvarez, P. (1995). Retrograde amnesia and memory consolidation: A neurobiological perspective.(2), 169–177.
Steinfurth, E. C. K., Kanen, J. W., Raio, C. M., Clem, R. L., Huganir, R. L., & Phelps, E. A. (2014). Young and old pavlovian fear memories can be modified with extinction training during reconsolidation in humans.(7), 338–341.
Suzuki, A., Josselyn, S. A., Frankland, P. W., Masushige, S., Silva, A. J., & Kida, S. (2004). Memory reconsolidation and extinction have distinct temporal and biochemical signatures.(20), 4787–4795.
Warren, V. T., Anderson, K. M., Kwon, C., Bosshardt, L., Jovanovic, T., & Bradley, B., Norrholm, S. D. (2014). Human fear extinction and return of fear using reconsolidation update mechanisms: The contribution of on-line expectancy ratings.(3), 165–173.
Yang, Y. L., Chao, P. K., & Lu, K. T. (2006). Systemic and intra-amygdala administration of glucocorticoid agonist and antagonist modulate extinction of conditioned fear.(5), 912–924.
Cue specificity of reconsolidation update mechanism in remote fear memories
HU Jingchu; ZHANG Weixin; CHEN Xiaoting; WANG Wenqing; WANG Zijie; ZHUANG Chuqun; FENG Biao; ZHENG Xifu
(School of Psychology, South China Normal University, Guangzhou, 510631, China) (School of Life Sciences, South China Normal University, Guangzhou, 510631, China) (Sun Yat-Sen Memorial Secondary School, Zhongshan, 528400, China)
Recent studies of fear memories conducted in both humans and animals have suggested that new fear memories (1-day old) can be attenuated using a reconsolidation update mechanism, which is selective to the reactivated cue. In real life, patients with post-traumatic stress disorder (PTSD) usually receive treatment much after the traumatic memories form, and a traumatic event can be associated with multiple cues. However, the cue specificity of reconsolidation update mechanism in remote fear memories (> 7 days) remains largely unknown.
To assess the cue specificity of remote fear memories (14 days) reconsolidation, we explored whether retrieval-extinction during the reconsolidation time window of remote fear memories is selective to the reactivated cue. We used a within-subject design, and skin-conductance response (SCR) served as the measurement. All subjects underwent fear conditioning by three coloured squares on day 1. Two squares (CSa+ and CSb+) were paired with the shock on 38% of the trials. The third square (CS?) was never paired with the shock. Fourteen days later, subjects received a single presentation of CSa+ (reactivated CS+) but not CSb+ (non-reactivated CS+). Ten minutes after the reminder trial, extinction training was conducted (within the reconsolidation window). Twenty-four hours later, all subjects returned to the experiment room and received spontaneous recovery test of the remote fear memories.
Results showed that there is no recovery for the reminded CS+, but significant recovery for non-reminded CS+ during spontaneous recovery testing. The recovery index (which was calculated as the first trial on day 16 minus the last trial on day 15 by differential SCR) of non-reminded CS+ was found to be significantly higher than that of the reminded CS+ (< 0.05). Thus, retrieval-extinction during reconsolidation window only attenuates the fear memory of the reactivated cue.
Our findings demonstrated that the reconsolidation update mechanism is effective for attenuating remote fear memories, and that this mechanism is selective to the reactivated cue of remote memories. We provide evidence to support the ongoing efforts in the development of novel strategies to combat remote pathogenic memories, which we think could lead to a more effective application of the reconsolidation update mechanism.
remote memory reconsolidation; cue specificity; conditioned fear; retrieval-extinction
10.3724/SP.J.1041.2019.00316
2017-12-05
* 國家自然科學基金(31771218), 廣東省自然科學基金博士科研啟動基金資助(2018A030310661), 中國博士后科學基金第64批面上項目資助(2018M640791)。
胡靜初和張蔚欣同為第一作者
鄭希付, E-mail: zhengxf@scnu.edu.cn
B845
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
學苑創造·A版(2020年10期)2020-11-06 05:21:26
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
作文周刊·小學一年級版(2016年27期)2017-06-03 23:21:17
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
絲綢之路(2016年9期)2016-05-14 14:36:33
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
