飼料脂肪含量對兩種規格的軍曹魚生長、體組成和血清生化指標的影響
2019-03-11 09:03:52劉迎隆麥康森張彥嬌艾慶輝
水生生物學報 2019年2期
劉迎隆 麥康森 徐 瑋 張彥嬌 王 震 艾慶輝
(中國海洋大學農業部水產動物營養與飼料重點實驗室, 教育部海水養殖重點實驗室, 青島 266003)
隨著養殖產業的不斷發展, 優質蛋白源, 尤其是魚粉的供需不平衡勢必會成為阻礙產業健康可持續發展的障礙。蛋白質、脂肪和糖類都可作為能量源為生物體供能, 脂肪和糖類均具有一定的蛋白節約效應[1]。相較于價格較高的蛋白質類和利用效率有限地糖類, 單位能值更高的脂肪越來越受到人們的關注[2, 3]。另外, 除了供能之外, 被吸收的脂肪還可為魚體正常生長代謝提供必需脂肪酸[4]。當飼料中脂肪含量不足時, 可使養殖魚類出現脂溶性維生素和必需脂肪酸缺乏癥, 同時還會出現飼料蛋白利用率低下等癥狀[5]。但是, 近年來隨著高脂飼料的使用,脂肪在魚體的異常沉積正越來越受到人們的關注[6]。飼料中過量的脂肪會導致脂肪沉積在腹腔和組織中[7—9]。脂肪的異常過度沉積不僅會造成脂肪原料的浪費進而提高生產成本, 而且會影響到魚類的代謝與免疫能力, 還會直接影響養殖魚的品質[2, 6, 10]。另外, 隨著魚類的生長發育, 魚類在不同生長階段其消化[5, 11]和代謝能力[5, 12]會有所不同, 因此魚類在不同生長階段對營養物質的需求存在著顯著的差異[13, 14], 尤其是作為主要能量源的脂肪[3, 5, 15]。通過生長實驗確定出不同生長階段養殖魚類的脂肪需求量, 無疑可以通過最大程度地利用蛋白原料, 降低生產成本來保證水產養殖產業的健康可持續發展。
軍曹魚(Rachycentron canadumL.)隸屬于鱸形目(Perciformes)中的鱸亞目(Perxoidei)、軍曹魚科(Rachycentridae)、軍曹魚屬(Rachycentron), 體細長、頭扁平、近圓筒狀[16], 體披細小圓鱗, 屬于暖水廣鹽性魚中底層肉食性海水魚類; 無鰾, 需要靠不斷游動保持身體平衡; 其不耐饑餓具有殘食習性[17]。生長快、抗病力強、產量高的軍曹魚無細棘骨, 同時肉厚質細、肉質鮮美是制作生魚片的上好材料[18]。隨著人工繁殖以及海水網箱養殖技術的發展, 軍曹魚已成為我國東南沿海地區近海網箱養殖的重要品種[19]。目前, 對于軍曹魚脂肪需求的研究主要集中在幼魚階段[8], 而對于大規格的軍曹魚, 尤其是不同規格的軍曹魚的脂肪營養需求的研究尚未見報道。本研究通過配制不同脂肪水平的飼料投喂2種規格的軍曹魚, 以FER和SGR為評定指標, 構建二次回歸曲線來估算不同生長階段的軍曹魚的脂肪需求量。同時, 探討飼料脂肪水平對軍曹魚魚體、肝臟和肌肉組成、血清生化指標的影響,為不同規格的軍曹魚高效配合飼料的開發提供理論依據。
1 材料與方法
1.1 飼料配方與制作
原料(主要蛋白源: 魚粉、酪蛋白和豆粕; 糖源:小麥粉和小麥淀粉)粉碎后過40目篩, 后按逐級放大的原則手工配合高效混合機(0.5-V型, 上海天祥健臺制藥機械有限公司)均勻混合原料。之后, 手工混入魚油和磷脂油并加入適宜的水, 擠條并加工為6種脂肪含量的等氮的(2種規格: 4和6 mm)硬顆粒飼料(表 1)。冷氣房中空氣晾干至水分10%左右后儲存備用(-20℃)。
1.2 飼養管理
將從廣東湛江雷州購來的同一批孵化的軍曹魚苗置于廣東湛江廣東粵海飼料公司南三島實驗基地1.5 m×1.5 m×2.5 m淺海網箱中進行實驗, 每個處理3個重復。在實驗開始前, 投喂商業飼料至實驗魚達到相應規格。稱重分魚[40 g規格組(38.24±0.30) g, 25尾/箱; 500 g規格組(529.17±5.67) g, 10尾/箱]前禁食24h。實驗分別持續10周和8周, 每天08:00和17:00進行2次飽食投喂(緩慢投喂至實驗魚不再搶食為止), 記錄飼料每個網箱每日的投喂量。實驗期間環境如下: 水溫27—32℃, 鹽度21‰—23‰, pH 7.6—7.8, 氨氮水平<0.05 mg/L, 溶解氧≥6.0 mg/L。

表 1 飼料配方及主要營養成分組成(%, 干重)Tab. 1 The formulation and proximate composition of the experimental diets (%, dry matter)
1.3 樣品收集與分析
樣品收集分魚時各預留30尾魚進行體成分分析。在養殖實驗結束后, 禁食24h, 利用丁香酚(1∶10000)麻醉后, 迅速計數并稱重, 每個網箱隨機取3尾魚迅速測量體重和體長并儲存于-20℃用于全魚成分分析。再從每個網箱隨機取3尾實驗魚,使用抗凝劑(肝素鈉)處理過的無菌注射器取血后,取下針頭將血液注入離心管中冰上靜置。4℃條件離心(3500×g)10min, 取上清液, 將血清于-80℃保存。采完血后迅速測定體重及體長, 解剖并對內臟和肝臟稱重, 收集肝臟和肌肉于-20℃保存用作成分分析的樣品。
飼料、全魚及組織成分分析飼料、全魚及組織水分、粗蛋白及灰分分別使用烘箱(105℃)、全自動凱氏定氮儀(KjeltecTM 8400, FOSS, Tecator,Sweden)和馬弗爐(550℃, 12h)參照AOAC的方法測定[20]。飼料及全魚粗脂肪采用AOAC中描述的索氏抽提法測定[20]。肝臟及肌肉脂肪采用氯仿甲醇(v∶v=2∶1)聯合抽提法測定[21], 糖原參照Hassid和Abraham描述的方法測定[22]。
血清生化指標分析使用Mindray公司提供的試劑盒, 配合LX-20型Beckman全自動生化分析儀, 血糖、血清總蛋白、血清總膽固醇、血清總甘油三酯、高密度脂蛋白-膽固醇和低密度脂蛋白-膽固醇分別用Barham等[23]、Peters[24]、Richmond等[25]、Schettler和Nussel[26]、Gordon等[27]和Okada等[28]所描述的方法進行測定。
1.4 公式及統計方法
存活率(%)=終末魚數目/初始魚數目×100%
特定生長率(SGR, %/d)=(Ln末重-Ln初重)/天數×100%
飼料效率(FER)= 魚體增重(濕重)/飼喂飼料(干重)
肝體比(HSI, %)=肝重/體重×100%
臟體比(VSI, %)=內臟團重/體重×100%
肥滿度(CF)=體重(g)/[體長(cm)]3×100%
使用SPSS 17.0對所得數據進行單因素方差分析(ANOVA), 若結果顯示差異顯著(P<0.05), 再用Tukey’s檢驗進行多重比較分析。所有數值用平均值±標準誤表示。
2 結果
2.1 生長指標
隨著脂肪含量的升高, 40 g規格和500 g規格軍曹魚特定生長率(SGR)先分別由1.76和0.21升高到2.13和0.46, 之后均呈現出降低趨勢(P<0.05); 40 g規格和500 g規格軍曹魚飼料效率(FER)先分別從0.48和0.09升高到0.70和0.20, 之后均呈現出降低趨勢(P<0.05)。以SGR為評定指標, 利用二次曲線模型為擬合模型, 40 g和500 g規格的軍曹魚的脂肪需求量分別為: 13.79%和13.49%; 以FER為評定指標,利用二次曲線模型為擬合模型, 40 g和500 g規格的軍曹魚的脂肪需求量分別為14.56%和13.10% (圖 1、圖 2)。綜上所述, 40 g規格軍曹魚的脂肪需求量為13.79%—14.56%, 500 g規格軍曹魚的脂肪需求量為13.10%—13.49%。對于2種不同規格的軍曹魚,相較于飼喂低脂(高糖脂比)飼料的實驗魚, 飼喂高脂(低糖脂比)飼料的處理魚均呈現出更高的臟體比(VSI)和肥滿度(CF)。而魚體肝體比(HSI)只在40 g規格的軍曹魚處理組中發現顯著性差異, 500 g規格處理組中卻未發現顯著差異(表 2)。
2.2 全魚及組織組成

圖 1 脂肪含量對2種規格的軍曹魚特定生長率(SGR)的影響Fig. 1 Relationship between different lipid levels and SGR of cobia at two body sizes
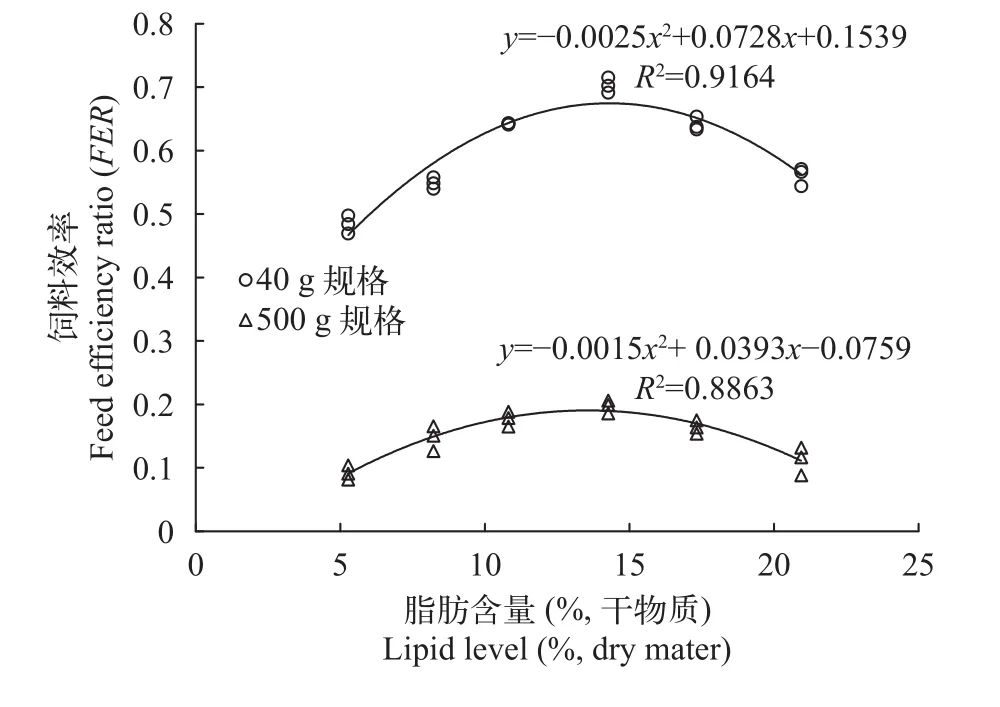
圖 2 脂肪含量對2種規格的軍曹魚飼料效率(FER)的影響Fig. 2 Relationship between different lipid levels and FER of cobia at two body sizes
隨著飼料中脂肪含量的增加, 40 g規格軍曹魚其體水分和蛋白含量顯著降低, 而體脂含量顯著升高(P<0.05), 而在500 g規格組卻未發現顯著差異(P>0.05)。在肝臟和肌肉中, 40 g規格軍曹魚組織脂肪含量均隨飼料中脂肪的增加顯著升高, 而組織水分則呈現出相反的趨勢(P<0.05), 而500 g規格軍曹魚各處理之間卻未發現顯著性差異(P>0.05)(表 3—5)。
2.3 血清生化指標
隨著飼料脂肪水平的升高, 2個不同生長階段的軍曹魚其血清總甘油三酯(TG)水平均顯著升高(P<0.05), 而血清總膽固醇(TC)、低密度脂蛋白-膽固醇(LDL-C)、高密度脂蛋白-膽固醇(HDL-C)和血糖(GLU)含量卻只在40 g規格處理組間發現顯著差異(P<0.05), 在500 g規格處理組中則未發現顯著差異(P>0.05) (表 6)。
3 討論
3.1 生長指標
本實驗發現, 2種規格的軍曹魚的存活率都不受攝入飼料中脂肪含量變化的影響。這與黃尾 (Seriola dumerilii)、黃顙魚(Pseudobagrus fulvidraco)、斜帶石斑魚(Epinephelus coioides)、大黃魚(Pseudosciaena croceaR.)等的研究結果一致[29—32]。作為重要的能量源, 隨著飼料中脂肪含量的增加,2種規格的軍曹魚SGR與FER均呈現出先上升再下降的趨勢。這與之前對大黃魚[32]、斜帶石斑魚[5]、團頭魴(Megalobrama amblycephala)[15]等研究的結果一致。這主要是因為, 當飼料中脂肪水平由不足提高到適宜水平時, 飼料中必須脂肪酸的含量及總能水平提高, 進而起到促進魚體生長和節約蛋白質的作用[33, 34]。而當飼料中脂肪的含量超過最適需求量時, 脂肪可能會在肝臟等組織中出現異常沉積,進而影響魚體的正常生長代謝, 起到抑制生長的作用[2, 6, 9, 32, 35—37]。另外, 本研究還發現, 隨著魚體規格的增大,SGR呈降低趨勢, 這與之前的研究結果是一致的[12, 13, 38, 39]。較小的魚類由于代謝強度較高, 其食欲更為旺盛[40], 因此隨著魚體規格的增大攝食率(FR)會出現減小的現象[39, 41]。同時, 隨著軍曹魚的生長發育, 其尾鰭形狀會發生顯著變化, 稚魚期為尖形, 幼魚期為圓形, 再經由截尾、凹尾, 成魚期為叉尾形[42]。從這一變化不難推測, 隨著軍曹魚的生長發育, 其游泳能力逐漸增強, 代謝水平也隨之升高。因此, 綜合以上因素推測攝食率的下降或維持自身新陳代謝所消耗能量的增加是導致特定生長率下降的主要原因。
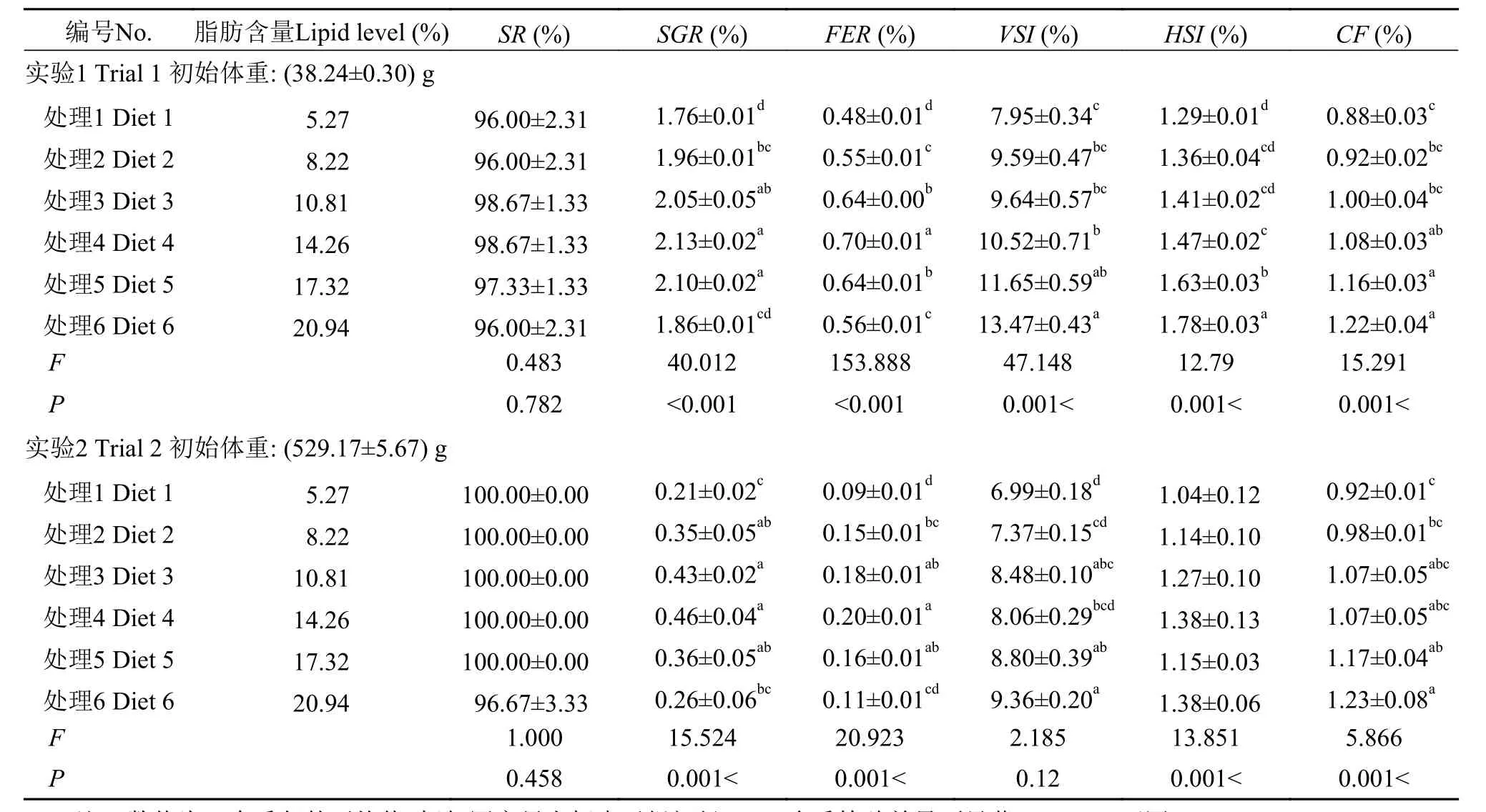
表 2 軍曹魚2個不同生長階段成活率(SR)、特定生長率(SGR)、飼料效率(FER)、臟體比(VSI)、肝體比(HSI)、肥滿度(CF) (%, 濕重; 均值±標準誤)*Tab. 2 Effect of lipid levels on Survival ratio (SR), Specific growth rate (SGR), Feed efficiency ratio (FER), Viscerosomatic index (VSI),Hepatosomatic index (HSI) and Condition factor (CF) of cobia at two different body weight (Means±SEM)*
3.2 全魚及組織組成
研究表明魚體脂肪主要受食物等外源性因素影響[43]。本研究發現, 40 g規格的軍曹魚飼料脂肪含量的升高顯著降低了體蛋白質的含量, 而500 g規格的軍曹魚卻未受飼料脂肪含量變化的影響, 這與大黃魚的研究結果是一致的[32]。攝入過多脂肪時,脂肪主要會在腸系膜和肝臟等組織中沉積下來[6]。這可能是造成魚體VSI和CF上升的主要原因。軍曹魚幼魚全魚脂肪含量隨著飼料中脂肪含量的升高顯著升高(P<0.05), 而500 g規格的軍曹魚體脂肪并無顯著性差異(P>0.05)。根據40 g和500 g規格的軍曹魚的體組成變化趨勢, 可以推測在軍曹魚生長的過程中, 魚體對飼料脂肪的利用能力逐漸加強, 這與斜帶石斑魚[5]、大黃魚[32]上的研究結果是一致的。同時, 這可能也是脂肪在500 g規格的軍曹魚肝臟和肌肉等組織的沉積未顯現出顯著差異, 進而未顯著影響其肝體比(P>0.05)的原因。對2個不同規格的軍曹魚投喂相同營養組成的飼料, 養殖實驗結束后, 其脂肪沉積的效果卻存在著差異。這可能是由于魚體規格不同, 魚體或組織的蛋白或脂肪含量存在差異造成的, 另外, 不同規格軍曹魚之間的代謝差異可能也影響了最終的生長效果。
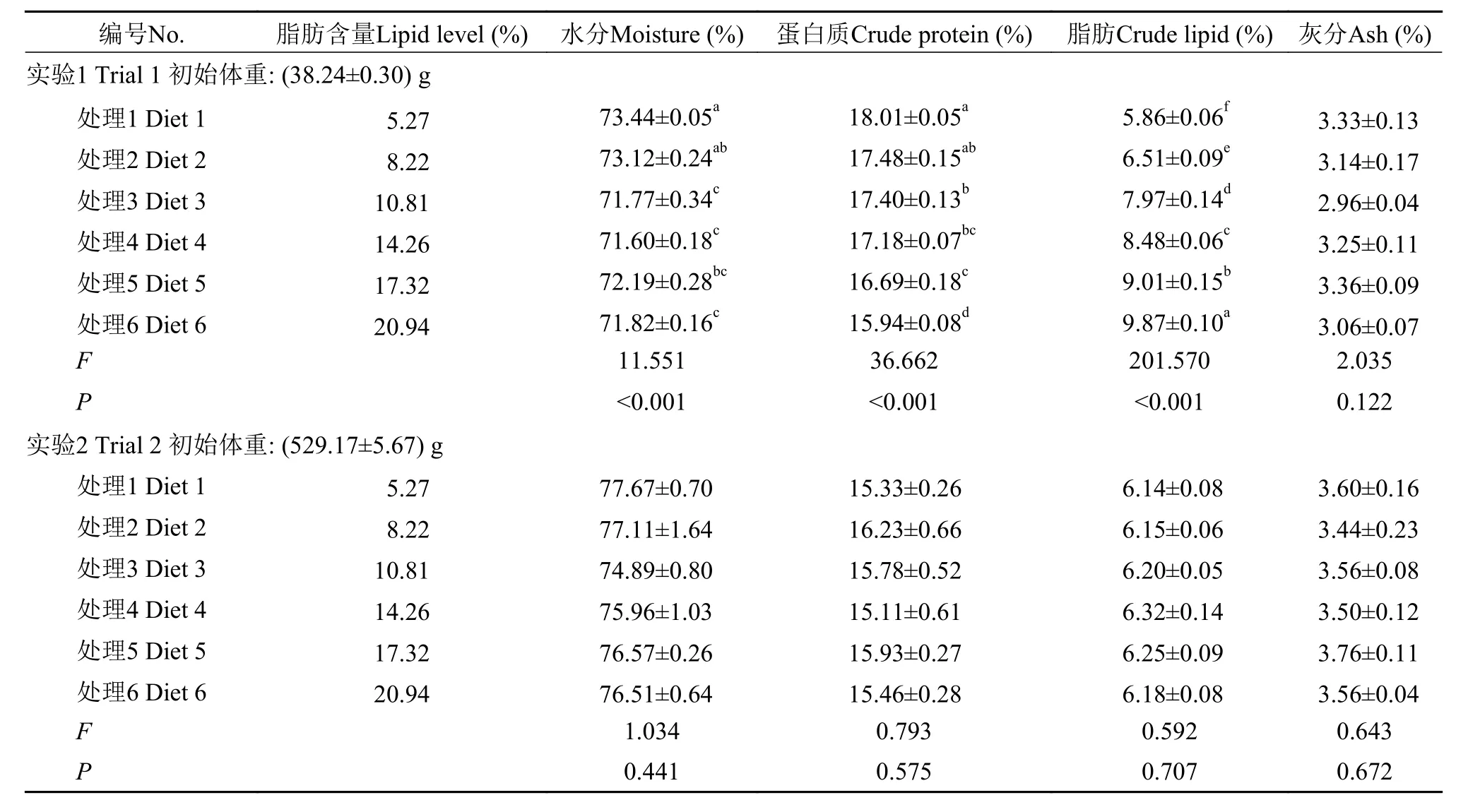
表 3 兩種規格的軍曹魚全魚組成(%, 濕重; 均值±標準誤)Tab. 3 Effect of lipid level on whole-body composition of cobia at two body weight (%, wet weight; Means±SEM)
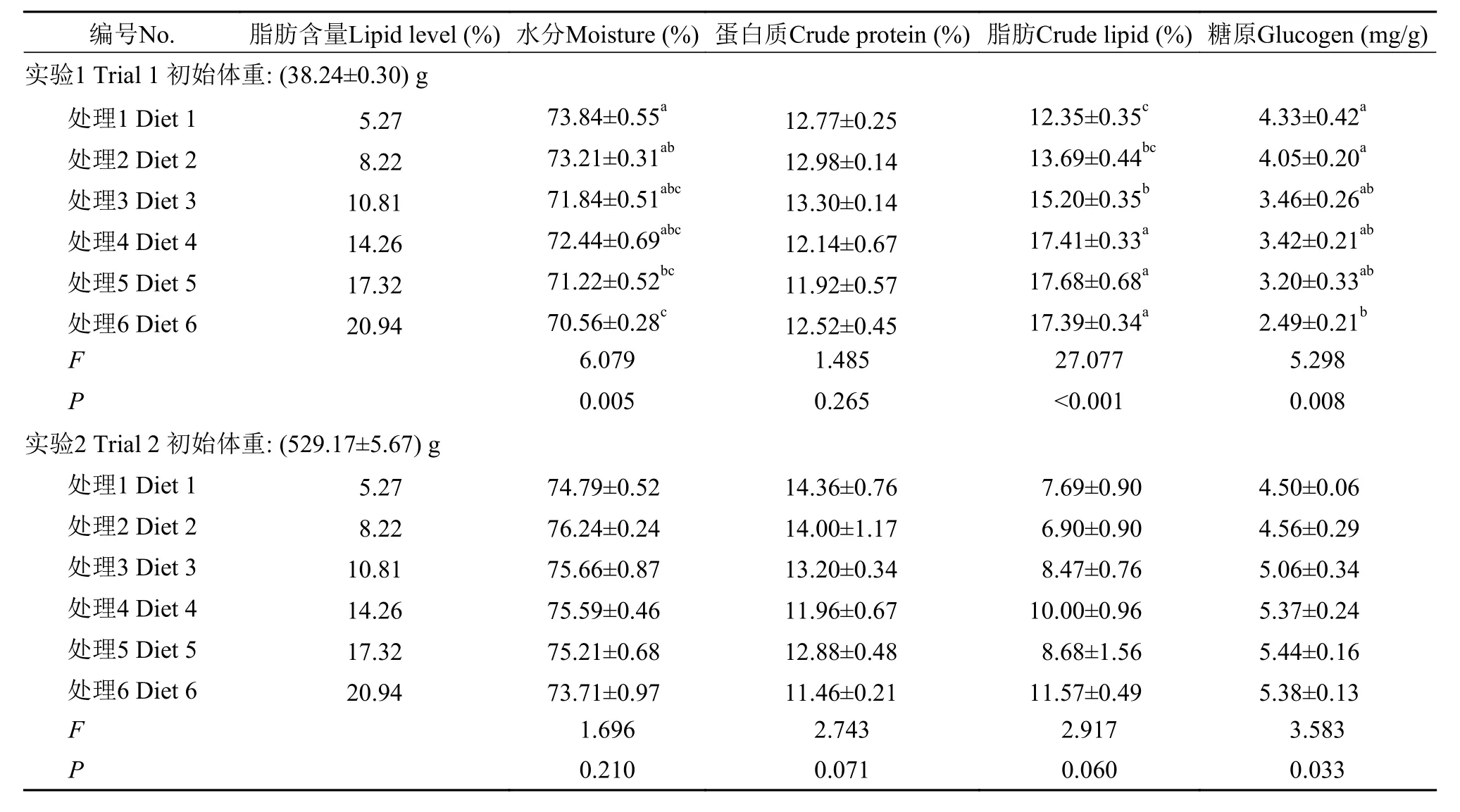
表 4 兩種規格的軍曹魚肝臟組成(%, 濕重; 均值±標準誤)Tab. 4 Effect of lipid level on liver composition of cobia at two body weight (%, wet weight; Means±SEM)
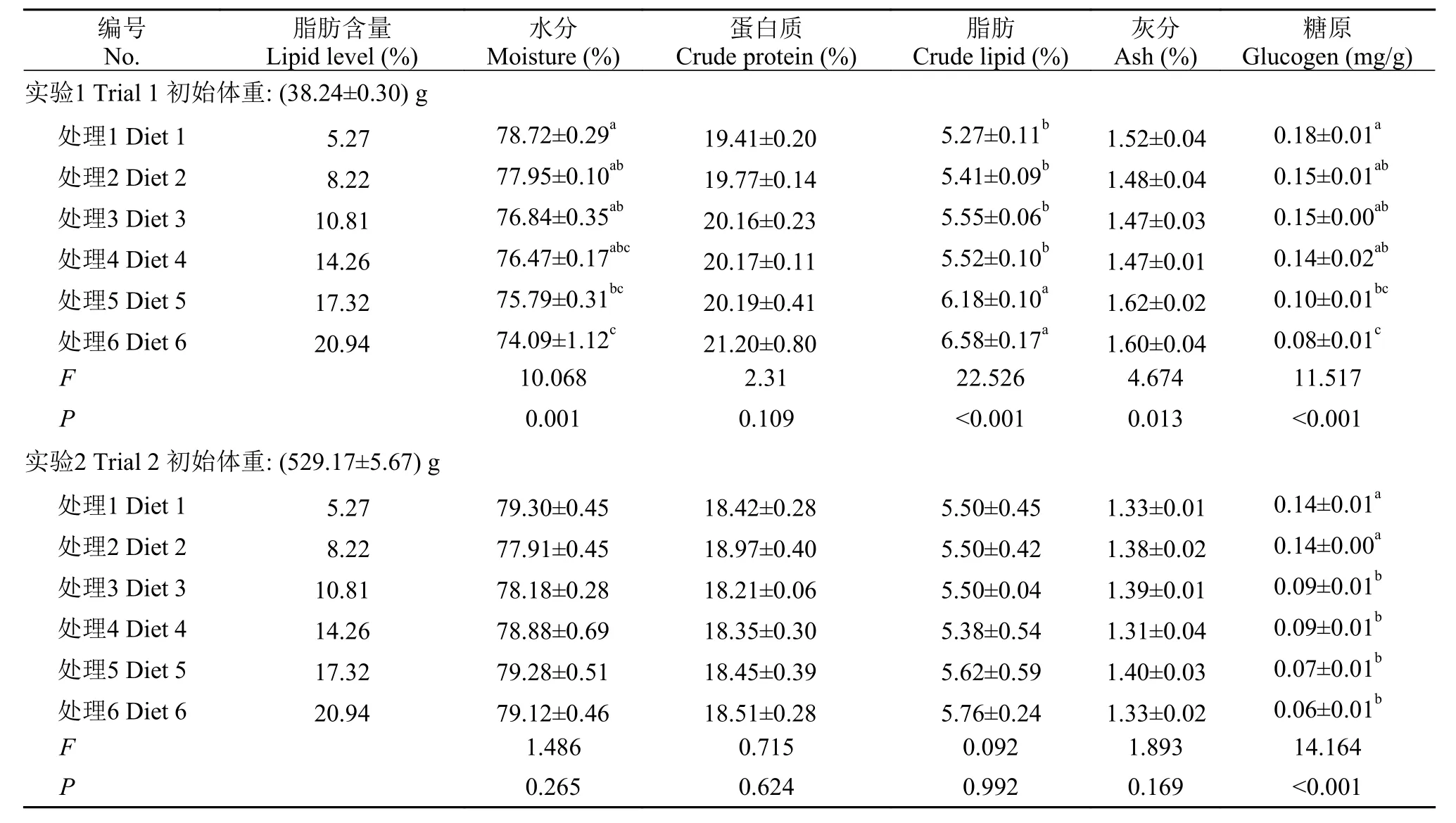
表 5 兩種規格的軍曹魚肌肉組成(%, 濕重; 均值±標準誤)Tab. 5 Effect of lipid level on muscle composition of cobia at two body weight (%, wet weight; Means±SEM)

表 6 兩種規格的軍曹魚血清生化指標(%, 濕重; 均值±標準誤)Tab. 6 Effect of lipid level on serum biochemical indices of cobia at two body weight (%, wet weight; Means±SEM)
3.3 血清生化指標
與哺乳動物類似, 魚類脂類轉運主要通過脂蛋白(Lipoproteins)經由外源性和內源性2個循環[44]。小腸中乳糜微粒(CM)轉運小腸從食物中吸收的脂類的方式稱為外源性途徑, 而內源性途徑是指肝臟中極低密度脂蛋白(VLDL)向外分泌脂類并轉化為中間密度脂蛋白(IDL)和低密度脂蛋白(LDL)[45, 46]。另外, 還存在著主要作用是將各組織中的膽固醇再運送回肝臟進行代謝的高密度脂蛋白(HDL)[32]。攝食高脂飼料的軍曹魚幼魚血清總甘油三酯、血清總膽固醇、LDL-C水平顯著升高, 這與草魚(Ctenopharngon idellus)[47, 48]、紅鰭東方鲀(Takifugu rubripes)[49]、大黃魚[50]和大菱鲆(Scophthal musmaximusL.)[51]上的研究結果是一致的。高水平的脂肪攝入并經由腸道消化吸收, 最終使血清TG含量升高。同時, 血清中高水平的總甘油三酯表明攝食高脂飼料組軍曹魚肝臟VLDL分泌量的增加。血管上皮細胞膜上的受體識別了VLDL后被LPL降解, 并最終轉化為LDL[51]。之前的研究表明: 魚體主要通過以下途徑對脂肪的轉運進行調控, 當攝入脂肪含量過低時, 魚體可經由apoB 100抑制VLDL的組裝和分泌[52, 53]。而攝入高脂飼料(具有高水平的n-3 LC-PUFA)時, 則可降低肝臟VLDL分泌[54]并且增加乳糜顆粒的代謝和清除[55—57]來降低血漿TG濃度,以便于抑制肝臟脂肪的過度沉積。隨著軍曹魚的生長發育, 該脂肪調節途徑的調節能力也會顯著提高。因此, 為闡明海水肉食性魚類糖脂代謝之間的關系, 還需對不同水平的糖和糖脂比的飼料對軍曹魚糖脂代謝關鍵酶的活性及表達水平的影響進行研究。
綜上所述, 以SGR和FER為評定指標, 40 g規格軍曹魚的脂肪需求量為13.79%—14.56%; 500 g規格軍曹魚的脂肪需求量為13.10%—13.49%。隨著飼料脂肪水平的增加, 血清TG、VSI與CF均顯著升高。而小規格的軍曹魚血清TC、HDL-C、LDLC和GLU以及魚體、肝臟和肌肉的脂肪含量均顯著升高, 魚體蛋白含量卻顯著下降; 而對較大規格的軍曹魚影響卻不顯著。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
小讀者(2021年2期)2021-03-29 05:03:48
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
