鎘脅迫對河南華溪蟹兩種C型凝集素免疫應答的影響
2019-03-11 09:04:04趙芳芳張左兵
水生生物學報 2019年2期
趙芳芳 郎 朗 張左兵 王 蘭
(山西大學生命科學學院, 太原 030006)
凝集素(Lectins)幾乎存在于所有動物、植物、微生物等生物有機體[1]。動物凝集素分為C型、F型、I型、L型、M型、P型及R型等13個家族[2]。其中, C型凝集素(C-type lectin, CTL)是一類糖類識別蛋白, 至少含有一個識別糖基的結構域(Carbohyrate recognition domain, CRD), 被稱為C型凝集素樣結構域(C-type lectin domain, CTLD), 可特異性地結合異物表面的糖類分子, 在蛋白轉運、細胞信號轉導、病原識別及作為效應分子等生物過程中起重要作用[2—4]。大量研究證明, 蝦蟹類CTL具有促進凝集、血細胞吞噬、血細胞包裹、激活酚氧化酶原系統及抗病菌等免疫功能[5—7]。目前, CTL已在凡納濱對蝦(Litopenaeus vannamei)[8]、羅氏沼蝦(Macrobrachium rosenbergii)[9]、中華絨螯蟹(Eriocheir sinensis)[10, 11]和三疣梭子蟹(Portunus trituberculatus)[12]等多種蝦蟹中成功克隆。CTL對病菌及其類似物脅迫會產生應答反應, 例如: 脂多糖能夠上調中華絨螯蟹肝胰腺EsLecD的表達[13], 嗜水氣單胞菌能夠下調中華絨鰲蟹EsCTLDcp-2在肝胰腺中的表達[14], 副溶血弧菌(Vibrio parahemolyticus)可誘導擬穴青蟹(Scylla paramamosain)Sp-lectin3在儲精囊和射精管中的表達以及Sp-lectin4在肝胰腺中的表達[15]。
鎘是廣泛存在于環境中的有毒有害物質[16]。研究表明, 鎘可以誘導細胞發生氧化應激產生大量的活性氧, 直接引起DNA、蛋白質等生物大分子的氧化損傷, 引發細胞凋亡并致癌[17, 18]。本課題組前期研究表明, 鎘脅迫能夠對甲殼動物生殖系統[19, 20]、消化系統[21]、免疫系統[22, 23]等造成損害。河南華溪蟹(Sinopotamon honanense)僅有非特異性免疫系統, 對重金屬污染敏感, 是研究重金屬脅迫下先天性免疫的理想模型。研究證實, 鎘能夠對河南華溪蟹免疫系統產生影響, 例如: 鎘脅迫能夠降低河南華溪蟹血淋巴細胞總數, 對超氧化物歧化酶、酸性磷酸酶、堿性磷酸酶等免疫相關酶活力也有不同程度的影響[22], 鎘脅迫能夠上調河南華溪蟹腸道及鰓中Shtoll3在響應嗜水單胞菌感染過程中的表達[23]。Ca2+是細胞內的第二信使, 參與多種信息傳遞。細胞內局部Ca2+特異的時空分布影響著鈣離子依賴性蛋白的功能和基因表達的模式。鎘可導致細胞鈣穩態失調, 干擾細胞內與Ca2+相關的信息傳遞。而鎘脅迫后Ca2+依賴的河南華溪蟹CTL的應答如何?對此, 目前尚未見相關的研究報道。
本研究在前期對河南華溪蟹轉錄組測序結果中鑒定的ShLec21與ShLec23部分序列基礎上, 采用RACE方法克隆了2種CTL的cDNA全長; 采用實時熒光定量PCR方法, 研究了2種CTL的組織表達模式和單獨鎘脅迫及鎘脅迫后再用細菌感染過程的中2種CTL的應答模式, 旨在為深入探索重金屬脅迫對無脊椎動物先天性免疫系統的影響提供科學依據。

表 1 引物名稱及序列Tab. 1 Primers used in this study
1 材料與方法
1.1 實驗材料
河南華溪蟹購自太原市五龍口海鮮市場, 體重1.9—3.5 g, 體長4.0—6.5 cm。在實驗室經曝氣自來水空腹飼養1周, 后喂食飼養1周, 實驗前1天停止喂食。實驗時, 取6只健康河南華溪蟹的血淋巴、鰓、肝胰腺、腸道、肌肉、卵巢以及精巢組織, 用于實時熒光定量PCR組織表達模式檢測。
實驗設置對照組(0 Cd), 低濃度鎘處理組(7.25 mg/L Cd, 1/32LD50)和高濃度鎘處理組(29 mg/L Cd,1/8LD50)三個組, 每組8只。在鎘處理7d后, 注射嗜水氣單胞菌(4.8×104cfu/mL, 1/4LD50)100 μL[23]。在整體處理24h后, 取血淋巴、肝胰腺, 用于實時熒光定量PCR檢測不同濃度鎘聯合細菌脅迫后ShLec21和ShLec23基因的表達變化。
隨機取健康河南華溪蟹, 用鎘濃度14.5 mg/L染毒, 經3個不同時間段(1d、4d和7d)處理, 同時設對照組(即鎘染毒0)。在每個時間段內又分為2個亞組: 亞組一注射0.85% 生理鹽水100 μL, 亞組二注射嗜水氣單胞菌(4.8×104cfu/mL; 1/4LD50)100 μL[23];每亞組6只蟹。在處理12h后, 取血淋巴、肝胰腺,以備后續實時熒光定量PCR檢測不同時間鎘聯合細菌脅迫后基因表達變化之用。
1.2 實驗方法
克隆河南華溪蟹ShLec21和ShLec23基因取河南華溪蟹血淋巴、鰓、肝胰腺和腸道等組織器官, 提取總RNA, 構建SMART cDNA文庫[23]。設計ShLec21與ShLec23的 3′RACE和5′RACE引物(表 1),分別以5′-RACE-Ready cDNA和3′-RACE-Ready cDNA為模板, 進行巢式PCR。首輪PCR反應使用KOD-FX-Neo (TOYOBO, 貨號: KFX-201), PCR擴增條件: 94℃ 2min; 98℃ 10s, 66℃ 30s, 68℃ 2min,37個循環; 68℃ 7min。將首輪PCR擴增產物稀釋100倍后作為模板, 使用Es Taq MasterMix(康維世紀, 貨號: CW0690)進行第二輪PCR反應, 擴增條件:94℃ 2min; 94℃ 30s, 66℃ 30s, 72℃ 3min, 37個循環; 72℃ 7min。RACE擴增產物經1.2%瓊脂糖凝膠電泳分析, 切膠回收, 連接pMD19-T-simple載體, 轉化Trans5α感受態細胞, 測序。最后, 對測序正確的序列進行拼接, 并設計引物(表 1), 使用KOD-FXNeo進行PCR擴增。同上測序, 進行ShLec21和ShLec23全長驗證。
使用NCBI的BLAST工具(https://blast.ncbi.nlm.nih.gov/Blast.cgi)對序列進行相似性分析, 使用ORF Finder程序(https://www.ncbi.nlm.nih.gov/orffinder/)對開放閱讀框進行分析, 使用SMART (http://smart.embl-heidelberg.de/)在線軟件預測結構域和信號肽,使用PROSITE (http://prosite.expasy.org/prosite.html/)對二硫鍵進行預測, 使用ClustalX1.8.1進行多重序列比對; 使用MEGA 5.05軟件以鄰接法(neighbourjoining, NJ)構建系統進化樹。
實時熒光定量PCR構建克隆質粒pMD19-T-simple-ShLec21和pMD19-T-simple-ShLec23并進行質粒提取(表 1), 經微量分光光度計測定質粒濃度及純度, 依據公式: 質粒拷貝數=(6.02×1023×質粒濃度)/[(2692+目的基因片段長度)×660][24], 計算出質粒拷貝數。將標準質粒用無菌水稀釋成101—1010copies/μL的10個梯度, 選取105、106、107、108、109和1010的6個梯度為模板, 以ShLec21和ShLec23特異性引物(表 1), 每個反應樣本設置2個重復, 使用SYBR?Premix ExTaqTMⅡ(TaKaRa, 貨號: RR20A)進行實時熒光定量檢測, 操作方法按照Lang等[25]描述進行。反應結束后根據結果繪制標準曲線。
取“材料與方法”中1.1健康河南華溪蟹血淋巴、鰓、肝胰腺、腸道、肌肉、卵巢和精巢, 以及不同處理條件的血淋巴和肝胰腺, 提取總RNA并反轉錄合成cDNA作為模板[23], 根據上述反應條件進行實時熒光定量反應。在反應結束后, 根據標準曲線, 計算得到ShLec21和ShLec23的準確拷貝數。采用GraphPad Prims 7繪圖, 結果用平均值±標準差(Mean±SD)表示。采用SPSS17.0 進行Kruskal-Wallis非參數檢驗; 采用Mann-Whitney檢驗兩組數據之間的統計學差異。*P<0.05表示差異顯著,**P<0.01表示差異極顯著。
2 結果
2.1 河南華溪蟹ShLec21和ShLec23的克隆與序列分析
通過RACE PCR擴增, 得到河南華溪蟹ShLec21和ShLec23全長cDNA序列。其中,ShLec21全長863 bp, 包括120 bp的5′UTR、編碼152 aa的長459 bp的開放閱讀框及284 bp的3′UTR。ShLec23全長681 bp, 包括72 bp的5′UTR、編碼164 aa的長495 bp的開放閱讀框及114 bp的3′UTR。通過SMART預測,ShLec21含有19 aa的信號肽和130 aa的CTLD;ShLec23含有17 aa的信號肽和134 aa的CTLD。并且,ShLec21與ShLec23結構域CTLD中分別含有EPE與EPI糖基結合位點。另外, 通過多序列比對及PROSITE預測,ShLec21含有2個保守的半胱氨酸殘基(Cys50和Cys149)形成一個二硫鍵;ShLec23含有4個保守的半胱氨酸殘基(Cys59、Cys138、 Cys154和Cys162)形成兩個二硫鍵。
2.2 河南華溪蟹ShLec21和ShLec23的系統進化分析
利用ClustalX1.8.1軟件, 將河南華溪蟹ShLec21和ShLec23, 與中華絨螯蟹、三疣梭子蟹、擬穴青蟹及凡納濱對蝦的CTL的氨基酸進行序列多重比對。結果表明,ShLec21和ShLec23與其他蝦蟹類CTL的CTLD結構域具有一定的保守性。ShLec21與中華絨螯蟹EsLecG的相似性為65%, ShLec23與三疣梭子蟹的PtLP相似性為48%。而ShLec21與ShLec23基因編碼的氨基酸序列相似性僅為32%。分別選擇不同的無脊椎動物和脊椎動物CTL的典型蛋白, 用NJ法構建CTL典型蛋白的系統進化樹。結果顯示,ShLec21和ShLec23分別與蝦蟹類CTL聚類在無脊椎動物分支中兩個不同的分支(圖 1)。
2.3 河南華溪蟹ShLec21和ShLec23的組織表達模式
河南華溪蟹ShLec21和ShLec23的標準曲線分別為:Y= -3.4501X+35.371、Y= -3.3598X+35.419,其中Y為CT值,X為起始模板數量。ShLec21與ShLec23標準曲線的回歸系數R2, 分別為: 0.9985和0.9995; 擴增效率E, 分別為: 94.92%與98.44%。結果顯示,ShLec21和ShLec23標準質粒在稀釋濃度范圍內存在良好的線性關系。
采用實時熒光定量PCR方法, 檢測了河南華溪蟹ShLec21和ShLec23在血淋巴、鰓、肝胰腺、腸道、肌肉、卵巢和精巢7種組織器官中的表達情況。結果顯示,ShLec21和ShLec23 主要在肝胰腺組織中表達, 在鰓組織中表達量較低, 而血淋巴、腸道、肌肉、卵巢和精巢組織中只有微量表達(圖 2)。
2.4 鎘脅迫對河南華溪蟹ShLec21和ShLec23應答變化的影響
實時熒光定量PCR結果顯示, 在鎘處理河南華溪蟹并用嗜水氣單胞菌感染過程中, 肝胰腺、血淋巴ShLec23在29 mg/L高濃度鎘組中, mRNA表達量分別顯著(P<0.05)與極顯著(P<0.01)高于對照組的,且存在明顯的劑量-效應關系。而在肝胰腺、血淋巴中ShLec21在不同處理組之間的表達沒有顯著性差異(圖 3)。

圖 1 無脊椎動物和脊椎動物CTL的NJ系統進化樹 (河南華溪蟹的兩種CTL用下劃線標出)Fig. 1 NJ phylogenetic tree constructed from the amino acid sequence of C-type lectin of invertebrate and vertebrate (Two C-type lectins are marked in underline of S. honanense)
如圖 4a—b所示, 河南華溪蟹肝胰腺與血淋巴中ShLec21表達量, 在單獨鎘脅迫不同時間段(0、1d、4d和7d)無顯著性差異; 在單獨嗜水氣單胞菌感染后(即單獨鎘染毒0),ShLec21表達量在肝胰腺組織中顯著性下調(P<0.05); 但是, 在鎘脅迫不同時間段(0、1d、4d和7d)后嗜水氣單胞菌感染過程中,ShLec21在肝胰腺中的表達量在處理4d的較1d和對照組的顯著性上調(P<0.05), 7d的較1d的呈顯著性上調(P<0.05),ShLec21在血淋巴中的表達量在處理1d的較對照的顯著性上調(P<0.05)。
如圖 4c—d所示, 河南華溪蟹肝胰腺和血淋巴中ShLec23表達量, 在單獨鎘脅迫不同時間段(0、1d、4d和7d)同樣無顯著性差異; 在單獨嗜水氣單胞菌感染后,ShLec23表達量在肝胰腺中表現為極顯著(P<0.01)下調, 在血淋巴中表現為顯著性下調(P<0.05); 但是, 在鎘脅迫不同時間段(0、1d、4d和7d)后嗜水氣單胞菌感染過程中,ShLec23在肝胰腺中的表達量鎘處理4d的較1d的表現為顯著性上調(P<0.05), 4d的較對照的表現為極顯著性上調(P<0.01),ShLec23表達量在血淋巴中4d的較對照的顯著性上調(P<0.05)。
3 討論
脊椎動物CTL蛋白家族具有高度保守性, 而無脊椎動物則呈現一定多樣性。在結構上, CTL至少含有一個形成雙環結構的CTLD結構域[26]。河南華溪蟹Shlec21和Shlec23均有一個CTLD, 與墨吉對蝦(Fenneropenaeus merguiensis)FmLC2[27]、中華絨鰲蟹EsLecG[10]和三疣梭子蟹PtLP[12]的報道一致。而有的CTL中含有2個CTLD, 例如羅氏沼蝦MrLec1、MrLec2、MrLec3及MrLec4[28]。CTL雙環結構的穩定主要依賴于4個保守的半胱氨酸形成的二硫鍵[29],Shlec23 CTLD含有4個保守的半胱氨酸(Cys59、Cys138、Cys154和Cys162), 形成2個二硫橋, 具有典型的雙環結構。Shlec21則僅有2個保守的半胱氨酸殘基(Cys50和Cys149)形成一個二硫橋, 沒有典型的雙環結構, 這方面還未見相關報道。CTL的CTLD中含有4個Ca2+結合位點, 其中Ca2+結合位點2可通過2個典型的基序參與對糖基的識別[29]。第一個基序為EPN (Glu-Pro-Asn), 或者QPD (Gln-Pro-Asp); 第二個基序為WND (Trp-Asn-Asp)。EPN與QPD位點分別識別甘露糖和半乳糖[29]。2個糖基識別基序常會出現變異, 例如EPD、EPK、EPS、EPQ、QPG、QPS、QPN、QPT、YPT、WHD、FND及MND等[2, 30], 或者在CTLD檢測不到。在ShLec21與ShLec23中, 分別含有EPE (Glu-Pro-Glu)和EPI (Glu-Pro- Ile), 兩者可能是EPN位點的變異, 替代EPN識別甘露糖。而EPE位點在中華絨鰲蟹EsLecGCTLD中也有發現[10]。另外, 系統進化結果表明,ShLec21和ShLec23雖然同屬于無脊椎動物分支, 但是隸屬無脊椎動物中不同的亞類。因此, 我們推測ShLec21和ShLec23在河南華溪蟹的先天性免疫中可能發揮不同的作用。
CTL具有不同的組織表達模式。凡納濱對蝦LvCTL3在肝胰腺、胃、幽門盲囊、上皮、肌肉、眼柄、柄節、腸、血細胞和鰓中廣泛表達[31], 而中國對蝦(Fenneropenaeus chinensis)Fclectin僅在血淋巴中有表達[32], 中華絨螯蟹EsCTLDcp、EsCTLD1、EsCTLD2及凡納濱對蝦LvLT僅在肝胰腺中表達[33, 34]。組織表達模式分析結果說明, 河南華溪蟹Shlec21和Shlec23在被檢測組織中普遍表達, 但主要在肝胰腺中表達。
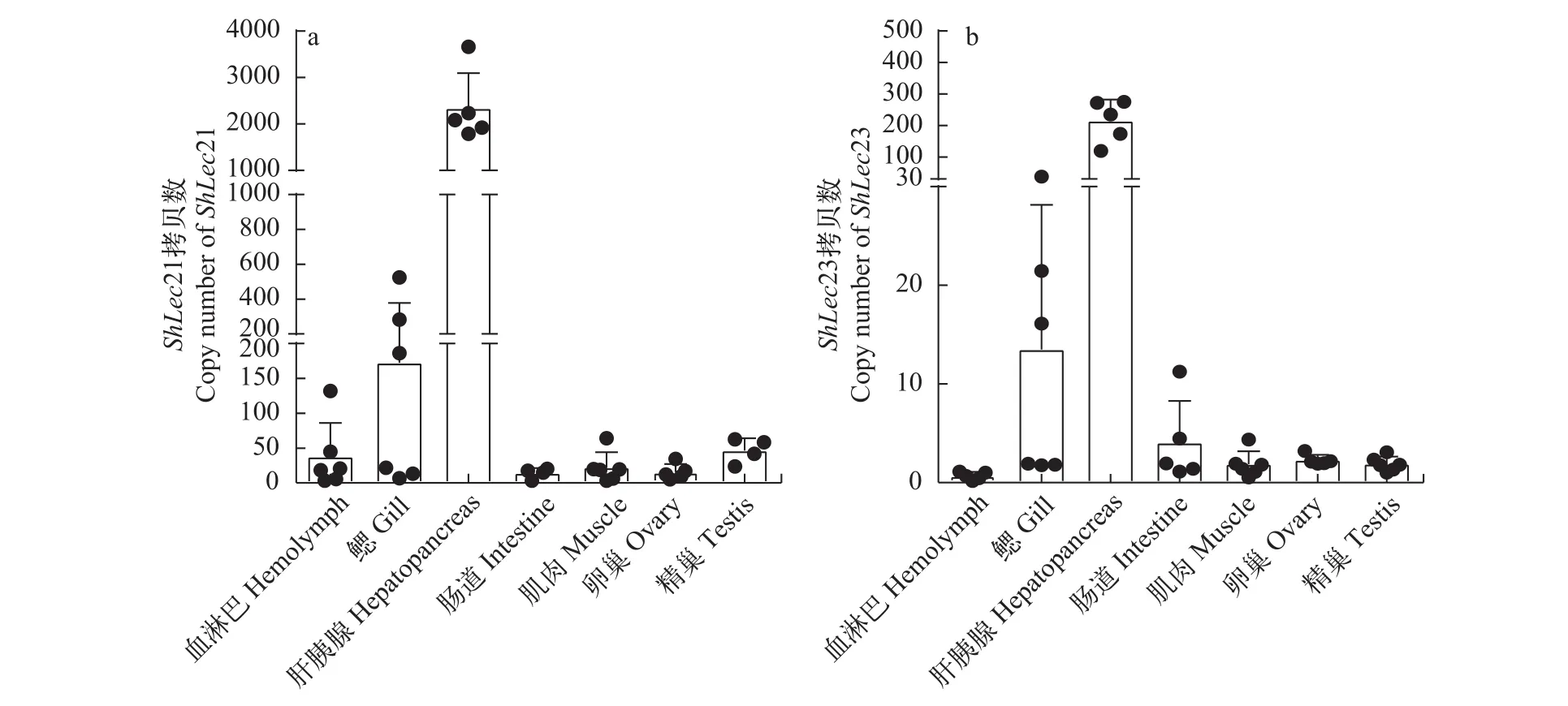
圖 2 河南華溪蟹各組織ShLec21和ShLec23 mRNA表達模式Fig. 2 Expression patterns ShLec21 and ShLec23 mRNA level in different tissues of S. henanense

圖 3 河南華溪蟹在不同濃度鎘處理后再用嗜水氣單胞菌感染肝胰腺和血淋巴中ShLec21和ShLec23 mRNA的表達模式Fig. 3 Effects on ShLec21 and ShLec23 expression in hepatopancreas and hemolymph of S. henanense by different concentrations of cadmium subsequently infected by bacteria A. hydrophila

圖 4 河南華溪蟹鎘脅迫不同時間再嗜水氣單胞菌感染后肝胰腺和血淋巴中ShLec21和ShLec23 mRNA表達模式Fig. 4 Effects of ShLec21 and ShLec23 expression in hepatopancreas and hemolymph of S. henanense by different treatment times of cadmium subsequently infected by bacteria A. hydrophila
Shlec21和Shlec23是存在于河南華溪蟹中2種鈣依賴活性的C型凝集素。為探明鎘脅迫對Shlec21和Shlec23免疫應答的影響, 本文首先檢測了河南華溪蟹不同濃度鎘處理后再用細菌感染過程中,ShLec21和ShLec23在肝胰腺和血淋巴兩個免疫組織中的應答變化。結果發現, 在高濃度鎘脅迫7d再用嗜水氣單胞菌感染后,ShLec23表達量在肝胰腺和血淋巴中顯著性上調。之后我們研究了河南華溪蟹經鎘脅迫不同時間段后再嗜水氣單胞菌感染過程中,ShLec21和ShLec23在肝胰腺、血淋巴兩個免疫組織中的應答變化。在本研究的脅迫條件下,單獨鎘處理不同時間, 對肝胰腺、血淋巴中ShLec21和ShLec23表達量沒有顯著性影響; 而在不同時間鎘脅迫河南華溪蟹后再細菌感染過程中, 肝胰腺和血淋巴中ShLec21和ShLec23 表達量在有些時間段上調。值得提到的是, 基因表達在脅迫時間維度上存在差異, 本研究結果也應考慮由于ShLec21和ShLec23表達在時間維度上存在的差異而對結果造成的影響。另外, 在本研究中嗜水氣單胞菌能夠下調ShLec21與ShLec23在肝胰腺或血淋巴中的表達(圖 4), 這與中華絨鰲蟹肝胰腺EsCTLDcp-2[14]結果一致。總之, 本研究表明, 鎘能夠上調河南華溪蟹ShLec21和ShLec23響應嗜水氣單胞菌感染過程中的表達, 在先天性免疫中具有一定的調節作用。
