瑞士乳桿菌特性及應用研究進展
2019-03-28 01:34:06岳瑩雪李柏良宋月閆芬芬王玉琦霍貴成
食品與發酵工業 2019年5期
岳瑩雪,李柏良,宋月,閆芬芬,王玉琦,霍貴成
(乳品科學教育部重點實驗室(東北農業大學),黑龍江 哈爾濱,150030)
瑞士乳桿菌(Lactobacillushelveticus)是革蘭氏陽性乳酸桿菌,最適生長溫度為42℃,通常呈白色或淺灰色不透明狀,在瓊脂平板上菌落的直徑為2~3 mm或更小。同型發酵產DL-乳酸,在甘露糖和果糖中緩慢的產酸或不產酸[1]。對瑞士乳桿菌進行基因組序列測序,然后進行基因功能的注釋,鑒定基因組編碼區的未知功能基因,可挖掘其在生物體生命活動中起重要作用的新功能因子,這對其基因組和菌株的益生功能兩方面的研究具有重要的意義[2]。瑞士乳桿菌具有公認的安全性和益生特性,在乳品中得到廣泛的應用,例如在牛奶中產生大量的乳酸,能夠迅速使奶酪裂解以及表達復雜的蛋白水解酶序列[3]。此外,它可作為主要的乳酸菌生產意大利和瑞士奶酪[4]。
本文從基因組特征、益生特性和應用3方面綜述了瑞士乳桿菌基因組與益生特性的聯系,蛋白水解系統及其產生的生物活性物質抗高血壓的益生特性,所產胞外多糖的益生特性,在腸道中和對宿主的免疫調節益生特性。最后,討論了瑞士乳桿菌在食品領域的應用及其表面活性劑的應用研究進展。
1 瑞士乳桿菌基因組特征
有時對益生菌的益生機理尚不能進行清楚的解釋,因此,需要通過基因組學、蛋白組學及代謝組學3方面,從遺傳的角度進行分析,以挖掘其潛在的益生特性。截至2018年,在National Center for Biotechnology Information(NCBI)網站能夠查詢到達到完成圖水平的瑞士乳桿菌13株,見表1。
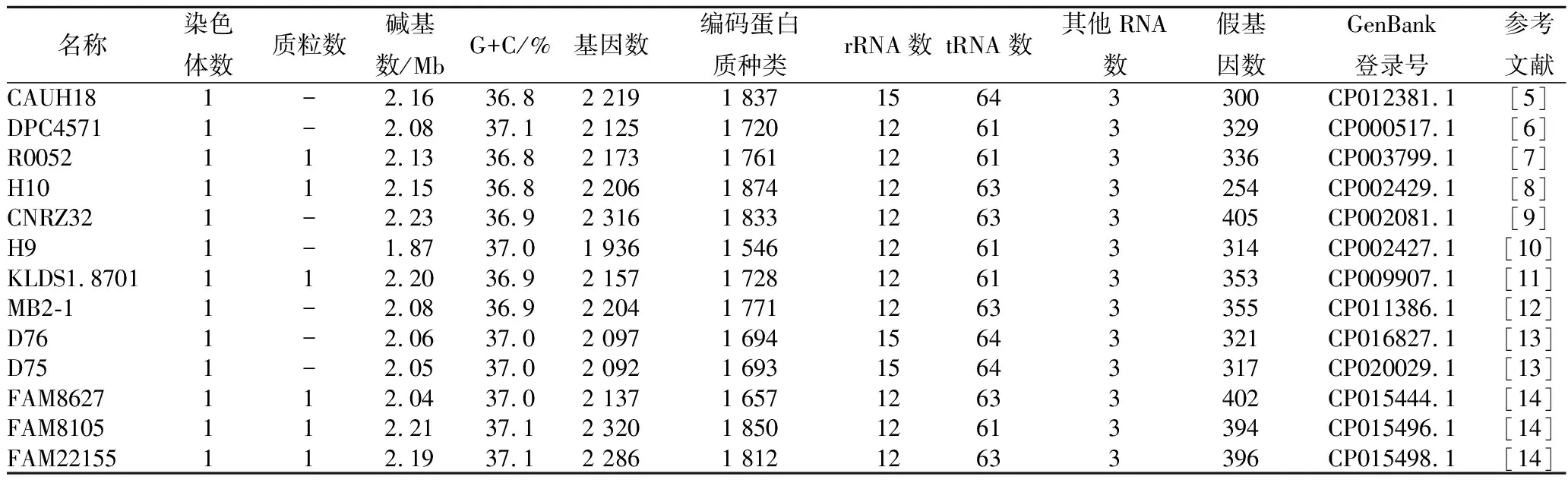
表1 13株瑞士乳桿菌的全基因組信息Table 1 Genome-wide information on 13 Lactobacillus helveticus
注:“-”表示無相關數據。
許多研究人員對瑞士乳桿菌進行全基因組分析,以更好地挖掘菌株自身所具有的益生功能。其中,基因組序列顯示:瑞士乳桿菌CAUH18具有新型胞外多糖生物合成的基因簇,同時含有可在胃腸道中定殖的細胞表面聚集促進因子的基因,該基因組序列為進一步研究其益生菌功能和分子遺傳學提供了基礎[5]。瑞士乳桿菌R0052只攜帶4種細胞膜包裹相關蛋白酶之一prtH4[15],它還具有獨特的表面層蛋白,已被證明可抑制大腸桿菌O157: H7與人細胞的黏附[16]。瑞士乳桿菌R0052具有編碼3種黏液結合蛋白(MUB)前體的基因,這些蛋白質在黏附到腸黏液層上發揮重要的作用,并且這種特殊的瑞士乳桿菌能夠持續存在于腸道中[7]。瑞士乳桿菌KLDS1.8701具有比其他測序的瑞士乳桿菌更多的黏液結合蛋白(HUO_04880,HUO_07225,HUO_07230和HUO_07235),這表明該菌株可能在腸道有較強的定殖能力,并與腸道微生物如益生菌的相互作用中占有優勢[11]。瑞士乳桿菌MB2-1基因組包含21個胞外多糖相關基因,其中10個基因獨特地存在于MB2-1,并被認為是確定獨特胞外多糖形成的關鍵酶。因此,MB2-1最重要的特征之一是具有產生高黏度和高產量胞外多糖的能力[12]。瑞士乳桿菌H9是一種能在牛奶發酵過程中產生抗高血壓肽的益生菌。利用瑞士乳桿菌的蛋白質組和轉錄組分析牛奶發酵過程,證明了某些肽酶和肽轉運系統可能是產生這種肽的關鍵因子[17]。瑞士乳桿菌CNRZ32與瑞士乳桿菌DPC4571[6],瑞士乳桿菌H10[8]和瑞士乳桿菌R0052[7]的全基因組比較表明,其含有超過180個預測的編碼序列,而不存在于其他測序的瑞士乳桿菌中。包括用于磷酸膽堿修飾的胞外多糖生物合成的獨特基因簇,幾個基因簇表明CNRZ32至少含有一個原噬菌體。比較基因組學也證明,瑞士乳桿菌CNRZ32是唯一測序的攜帶編碼4種不同細胞膜蛋白相關蛋白酶基因的瑞士乳桿菌菌株[15]。
2 瑞士乳桿菌蛋白水解系統及水解肽的抗高血壓益生特性
2.1 蛋白水解系統
乳酸菌的蛋白水解體系是其在牛奶中生長必不可少的,并且水解對發酵乳制品的風味也有許多影響,而瑞士乳桿菌被認為是對乳酪蛋白水解最有效的一種,其蛋白水解系統通常由3種類型的組分構成:(1)細胞包膜蛋白酶(CEP)將酪蛋白水解成寡肽;(2)將寡肽轉移到細菌膜上的轉運系統;(3)具有不同和部分重疊特異性的許多細胞內肽酶,形成游離細胞內氨基酸庫[19],瑞士乳桿菌蛋白水解系統如圖1所示。
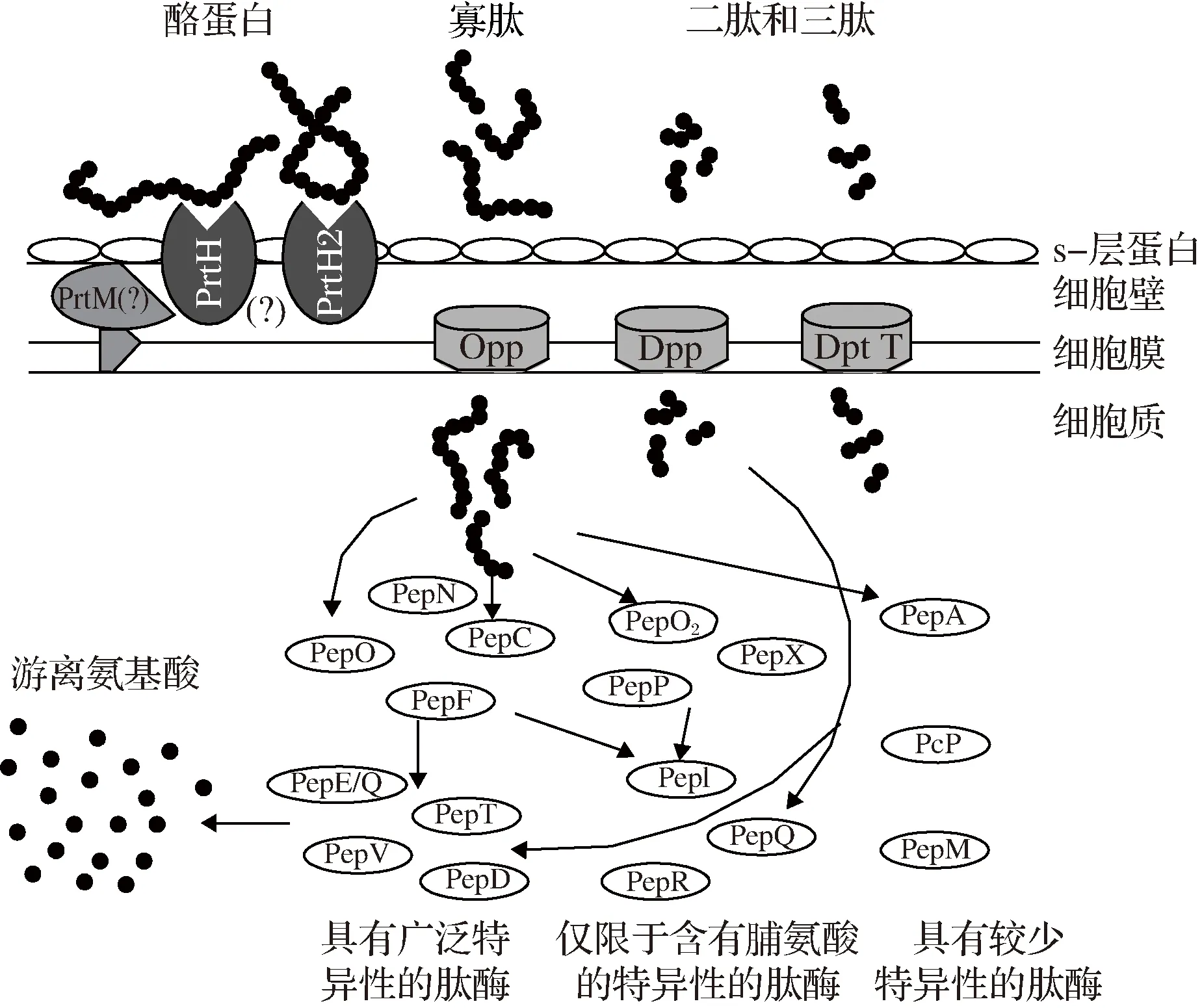
圖1 瑞士乳桿菌蛋白水解系統的示意圖Fig.1 Schematic diagram of Lactobacillus helveticusproteolytic system
但關于CEP在細胞表面的位置、PrtM的存在以及寡肽位移的大小仍然存在問題。根據文獻報道,瑞士乳桿菌可能擁有1個或多個編碼CEP的基因,將酪蛋白水解酶分解成寡肽[18]。根據生長培養基的蛋白水解活性差異,瑞士乳桿菌在富含肽或氨基酸的培養基(如MRS),或在補充有卡索松、卡薩米諾酸的化學成分確定培養基中,CEP的生物合成能力降低[20]。因此,與MRS培養基相比,在牛奶中生長的幾種瑞士乳桿菌的CEP活性高9~12倍[21]。對瑞士乳桿菌SBT11087的CEP活性研究發現,較少的αS1-酪蛋白由瑞士乳桿菌SBT11087細胞水解,同時β-酪蛋白的水解具有特異性,但這種特異性是有限的。降解之后,可在約20 kDa處觀察到條帶,該條帶含有β-酪蛋白的N端RELEE序列,這表明瑞士乳桿菌SBT11087的CEP最初水解的是β-酪蛋白C端區域。瑞士乳桿菌SBT11087的CEP水解酪蛋白效率低于其他CEP,但對β-酪蛋白具有特異性[22]。還有研究評估了4種瑞士乳桿菌ASCC953,ASCC474,ASCC1188和ASCC1315 的蛋白水解活性以及它們釋放生物活性化合物的能力,這些結果表明,瑞士乳桿菌發酵的牛奶可能釋放具有重要功能特性的生物活性化合物,這些化合物的特性和活性表現出高度應變和發酵時間依賴性[23]。
2.2 瑞士乳桿菌水解肽的抗高血壓益生特性
高血壓是心血管疾病的危險因素之一,包括冠心病,外周動脈疾病等。瑞士乳桿菌可以通過水解乳蛋白來釋放大量的肽,包括發酵乳中的生物活性肽,如抗高血壓肽Val-Pro-Pro(VPP)、Ile-Pro-Pro(IPP)和具有抑制血管緊張素轉換的活性酶(ACE),其中ACE是控制高血壓的有效靶點[24]。ELFAHRI等[25]研究發現一種瑞士乳桿菌發酵乳中血管緊張素轉化酶抑制活性最高,從開始時的6.33%上升到發酵結束時的48.69%,獲得的結果與之前的報道一致,并且明顯高于其他乳酸桿菌。KILPI等[26]研究的野生型瑞士乳桿菌CNRZ32在開始培養時表現出恒定且相當低的血管緊張素轉換的活性酶抑制活性,而在肽酶陰性突變體中,在90 h培養期間活性均勻增加,結果表明缺失pepN基因的瑞士乳桿菌可能會延緩短寡肽的水解,從而導致血管緊張素轉換的活性酶抑制肽在其水解成更短的肽或游離氨基酸之前暫時積累。PAN等[27]發現,通過3步反相高效液相色譜法對從瑞士乳桿菌JCM1004 的無細胞提取物產生的脫脂乳水解產物進行純化,產生了2種有效ACE抑制肽,這2種肽恢復了75%的初始ACE抑制活性,表明水解產生的大部分ACE活性歸因于這2種肽。通過序列分析,發現這2種肽是Val-Pro-Pro(VPP)和Ile-Pro-Pro(IPP)。VPP和IPP顯示出比其他報道的從奶制品中純化的血管緊張素轉換的活性酶抑制肽具有更高的血管緊張素轉化酶抑制活性。
3 瑞士乳桿菌所產胞外多糖的益生特性
胞外多糖(EPS)是從乳酸菌中分離出來的最重要組分之一,乳酸菌合成的胞外多糖大多以2種形式存在,一種是莢膜多糖(CPSs),它附著在細胞表面,另一種是釋放到細胞外滲透到培養基中的多糖(EPSs),能夠調節機體免疫力,是一類效果優良的免疫調節劑,它與抗氧化劑、抗癌劑、免疫調節劑和降膽固醇活性都相關[28]。LI等[29]研究了LHEPS (純化的胞外多糖組分)的體外抗增殖活性,利用來自瑞士乳桿菌MB2-1的3種純化LHEPS組分(LHEPS-1,LHEPS-2和LHEPS-3)和粗多糖LHEPS對人胃癌BGC-823細胞的增殖潛力進行研究,顯示出LHEPS具有潛在應用前景,特別是LHEPS-2有可能作為預防胃癌的功能成分。LI等[30]使用乳清作為基礎培養基,對瑞士乳桿菌MB2-1胞外多糖的最高產量進行優化,并對胞外多糖的化學性質、乳化、抗氧化劑和抗菌膜活性進行了研究,結果表明,EPS在不同的油、烴基質中表現出優異的乳化活性,體外抗氧化測定顯示胞外多糖具有強抗氧化性,特別是清除羥基和超氧自由基的能力。此外,抑制細菌生物膜的形成是該胞外多糖的另一個優點。目前,已有研究發現瑞士乳桿菌MB2-1在發酵乳制品中具有高黏和酸化的特性,分泌產生的莢膜多糖具有較好的抗氧化能力[31]。
4 瑞士乳桿菌菌體的免疫益生特性
人類腸道是一個復雜的環境,其中有多達1 000種不同的細菌會影響到腸道的上皮細胞,以及免疫系統[32]。而瑞士乳桿菌具有調控腸道免疫的益生特性。研究表明,瑞士乳桿菌對體重增加、行為、腸道微生物群和結腸細胞因子分泌的影響取決于小鼠的基因型和飲食[33]。王友湘[34]比較小鼠腸道微生物菌群,分別利用瑞士乳桿菌和大腸桿菌對小鼠進行灌胃,結果發現,瑞士乳桿菌對腸道乳酸菌和雙歧桿菌具有增殖的作用,對大腸桿菌和腸球菌等致病菌的生長具有抑制作用,能夠降低腸道內大腸桿菌的數量,起到免疫作用,而大腸桿菌對小鼠腸道黏膜免疫起到反作用。
瑞士乳桿菌還具有調節宿主免疫的特性。據報道,含有瑞士乳桿菌SBT2171(LH2171)的奶酪具有免疫調節作用,包括調節性T細胞的增加和小鼠中促炎性細胞因子的減少[35]。KIM等[36]研究的瑞士乳桿菌HY7801在膠原誘導的關節炎中表現出預防和治療效果,其主要是在CD4+ T細胞中上調IL-10表達,進一步的研究表明腸系膜CD11c+ DCs可能在益生菌誘導的免疫調節中起關鍵作用。瑞士乳桿菌BIM LhM3含有2種不同的噬菌體抗性機制:抑制噬菌體吸附和一個I型R/M系統。在瑞士乳桿菌中,噬菌體受體已經存在并確定在S層。噬菌體抗性菌株LhM3的S層蛋白質序列缺失6個氨基酸,它修飾了蛋白質折疊,結果表明此菌株干擾了噬菌體對宿主細胞的吸附,保護自己免受噬菌體攻擊[37]。
5 瑞士乳桿菌的應用
5.1 瑞士乳桿菌在食品方面的應用
瑞士乳桿菌常用于乳制品行業,制造超硬、半硬和瑞士奶酪發酵劑或輔助培養物,或用于某些發酵乳當中,現在也廣泛地應用于乳酒、食品酸乳飲料及飼料添加劑等發酵產品中。瑞士乳桿菌可產生許多具有抑菌活性的多肽物質,具有抗真菌活性,有研究表明其抗真菌來源于有機酸和瑞士乳桿菌KLDS1.8701自身,使其在食品保鮮、防腐和延長貨架期等方面起著重要的作用[38]。ZHOU[39]等將瑞士乳桿菌H9應用于酸奶中,評價其在發酵和儲存中的作用,結果表明瑞士乳桿菌H9可以促進發酵牛奶的酸化和蛋白水解,從而縮短發酵時間并且顯著提高發酵乳的ACE活性,此結果將對乳制品行業開發新型功能性乳制品具有重要意義。在干酪的生產過程中會產生大量的乳清,馬玲等[40]利用瑞士乳桿菌和干酪乳桿菌作為益生菌發酵劑,對發酵乳清飲料進行了研究。賀志華[1]研究了瑞士乳桿菌發酵綠豆乳,生產出的產品含有益生菌和豆類乳品2方面的功能特性,是一種新型的低脂肪,高蛋白的功能性乳制品。閔建[41]研究了瑞士乳桿菌和干酪乳桿菌制備大豆酸奶,這種組合比生產普通酸奶的菌株發酵大豆酸奶的口感要好,并且發現瑞士乳桿菌與干酪乳桿菌組合發酵產生的大豆酸奶各方面指標也都是最優的。因此,將益生菌例如瑞士乳桿菌用于大豆酸奶中是一個很好的研究方向。瑞士乳桿菌還可以制備凍干發酵劑,陳岑[42]利用高產黏瑞士乳桿菌MB2-1的凍干發酵劑制備酸乳并且使用酸乳粉制備具有多種用途的酸乳泡騰片,獲得集咀嚼、泡騰和再制酸乳多個優點于一體的益生酸乳泡騰片。
5.2 瑞士乳桿菌生物表面活性劑的應用
生物表面活性劑是具有表面和乳化性質的兩親性分子,細菌附著到表面后形成生物膜,對食品、環境和生物醫學領域都具有重要的影響。食品加工環境中生物膜的產生對消費者的健康有害。有研究表明,生物膜形成導致抗菌產品的抗性增加[43]。SHARMA等[44]對生物表面活性劑生產及其生物醫學和食品應用進行篩選,結果表明應用于生物醫學領域中的瑞士乳桿菌MRTL91 的生物表面活性劑,可用于替代抗微生物劑抵抗病原微生物,使其成為常規抗生素的合適替代物以供在食品和化妝品領域進一步使用。
6 展望
瑞士乳桿菌在益生菌中是蛋白水解能力較強的一種菌株,雖然水解酪蛋白產生的抗高壓肽已成為研究的熱點,但細胞包膜蛋白酶與酪蛋白相互作用的活性和特異性以及細胞包膜蛋白酶釋放肽的轉運系統還有待深入研究。除了其發酵產物中產生的抗高血壓肽外,瑞士乳桿菌還產生胞外多糖,利用發酵條件優化、基因和代謝工程的手段尋找高產胞外多糖的瑞士乳桿菌是接下來要進行的工作。由于胞外多糖的結構組成不同,對其進行分離純化、結構解析以及益生功能的研究也是今后的研究方向。瑞士乳桿菌及其代謝產物對抗氧化、抑菌、炎癥、癌癥等方面的作用已有研究,但對其發揮這些益生功能的機制未徹底研究清楚。利用微生物基因組學可發現瑞士乳桿菌中新的基因,并可分析它們所編碼的具有特殊功能的多肽和基因調控元件,從而加深對瑞士乳桿菌生命活動機理的重要代謝和調控機制的認識,所以在基因水平上的研究有利于瑞士乳桿菌功能基因研究、利用及菌種改良。在產品開發方面需開發新型功能性乳制品以發揮瑞士乳桿菌的應用價值。綜上所述,瑞士乳桿菌是具有廣闊發展前景的一種益生菌。
