Toll樣受體家族基因變異與結核易感相關性的研究進展
2019-04-11 05:40:24吉桂宜唐懷蓉張苗苗吳壽全賀建清
醫學綜述 2019年6期
關鍵詞:研究
吉桂宜,唐懷蓉,黃 燕,張苗苗,吳壽全,王 羽,賀建清※
(四川大學華西醫院 a.健康管理中心,b.呼吸與危重癥醫學科,成都 610041)
結核是由結核分枝桿菌感染引起的一種慢性傳染病[1-3]。2017年,全球有新發結核病1 400萬,同時有約160萬人死于結核[1]。中國是結核高發國家,發病率達63/10萬[1]。據報道,全球約1/3的人口感染結核分枝桿菌,然而僅5%~15%的感染者發展為活躍結核,其余人群以潛伏結核感染的狀態存在,表明結核易感性存在個體差異[4]。研究表明,宿主基因因素在結核的發病機制中起重要作用[5-6]。目前,已發現了許多重要的與結核易感性相關的候選基因,如細胞因子及其受體基因[腫瘤壞死因子、白細胞介素(interleukin,IL)-1β及IL-6]、趨化因子及其受體基因(趨化因子配體10及受激活調節正常T細胞表達和分泌因子)和模式識別受體基因[Toll樣受體(Toll-like receptors,TLRs)、甘露糖受體及Dectin-1(樹突狀細胞相關C型凝集素1)]等[5-6]。
機體依賴固有免疫反應及Th1細胞介導的適應性免疫反應來抵御結核分枝桿菌的入侵。TLRs是一類進化保守的胚系編碼的模式識別受體,可通過識別細菌的保守結構從而在機體免疫反應中發揮重要作用[7]。在結核分枝桿菌感染初期,TLRs就參與固有免疫反應,可促進巨噬細胞的直接殺傷功能和樹突狀細胞的分化成熟,激活獲得性免疫反應[8]。多項研究表明,TLRs的基因變異與結核易感性之間存在相關性[9-11]。現就TLRs基因變異與結核的相關性進行綜述,為探索新的抗結核治療方法提供理論依據。
1 TLRs簡介
TLRs廣泛存在于哺乳動物、人類、昆蟲及植物中。Toll蛋白最早是在果蠅體內發現的,稱為dToll,1997年,研究者首次在人體內分離出dToll的同系物,命名為TLR4[12-13]。目前,已發現哺乳動物有11種TLRs,其中,人類的10種TLRs是具有功能的[14]。人類TLRs屬于Ⅰ型跨膜蛋白受體,由胞外區、跨膜區及胞內區三部分組成[15]。胞外區由24個富含亮氨酸的重復序列(leucine-rich repeats,LRR)組成,可識別病原體;跨膜區是富含半胱氨酸的結構區域,與LRR區的C端連接;胞內區結構較保守,含200個氨基酸,與IL-1受體和IL-18受體的胞內區具有高度的同源性,因此,又被稱為Toll/IL-1受體同源區(Toll/IL-1 receptor homologous region,TIR),它是TLRs向下游信號轉導的重要部分[15-17]。相較于胞內區,不同TLRs的胞外區差異較大,這有助于對不同病原微生物配體的識別。
TLRs在人體中分布廣泛,不同細胞的TLRs種類存在差異。TLR1主要分布于單核細胞、樹突狀細胞、淋巴細胞、自然殺傷細胞及多形核白細胞;TLR2、TLR4、TLR5廣泛表達于除淋巴細胞及自然殺傷細胞外的免疫細胞;TLR3主要分布于未成熟的樹突狀細胞等[15]。不同的TLRs分布于細胞的不同區域,如TLR1、TLR2、TLR4、TLR6分布于細胞表面,TLR3、TLR7、TLR8、TLR9分布于細胞內[15]。每種組織至少表達一種TLRs,不同組織器官的TLRs種類和數量也存在差異,如脾臟和外周血中可表達全部的TLRs,而與外界接觸的部位(如呼吸道),TLRs的種類和數量均是最多的[18-20]。
TLRs最主要的功能是促進細胞因子的合成與釋放,引起炎癥反應,同時還可促進免疫細胞的成熟與功能化,從而介導機體的固有免疫及獲得性免疫反應[21]。它主要通過以下兩條信號途徑發揮作用[15,22]:①髓樣分化因子88(myeloid differentiation factor 88,MyD88)依賴途徑。當TLRs被激活后,誘導活化其下游的MyD88及IL-1受體相關激酶[15]。除TLR3外的TLRs均可激活該通路。其中,僅TLR2與TLR4需要MyD88和TIR功能區的接頭蛋白協同作用完成信號轉導[15,23]。②MyD88非依賴途徑。當TLRs被激活后,誘導活化其下游的腫瘤壞死因子受體相關因子6。TLR3及位于內涵體上的TLR4通過該通路發揮作用[23]。兩條途徑均可進一步活化核因子κB,引起級聯信號放大反應,從而啟動各種炎癥細胞因子基因的轉錄,產生一系列免疫和炎癥反應[22],見圖1。
2 TLRs與抗結核感染
固有免疫反應是機體對抗結核分枝桿菌感染的第一道也是非常重要的屏障。人體吸入結核分枝桿菌后,首先激活肺泡巨噬細胞及樹突狀細胞產生的固有免疫反應[24]。在感染初期,吞入巨噬細胞內的結核分枝桿菌可進行胞內復制,并可有效穿過肺泡間隔播散到肺外組織[24]。胞內復制及病原體的傳播發生在適應性免疫反應之前,表明結核分枝桿菌可建立一種避免被機體免疫系統清除并可持續存在的獨特自我保護機制[25-26]。固有免疫系統有各種模式識別受體,它們存在于全身各個器官和組織中[27]。模式識別受體可特異性識別病原相關分子模式,從而啟動并調節機體的固有免疫反應[28]。
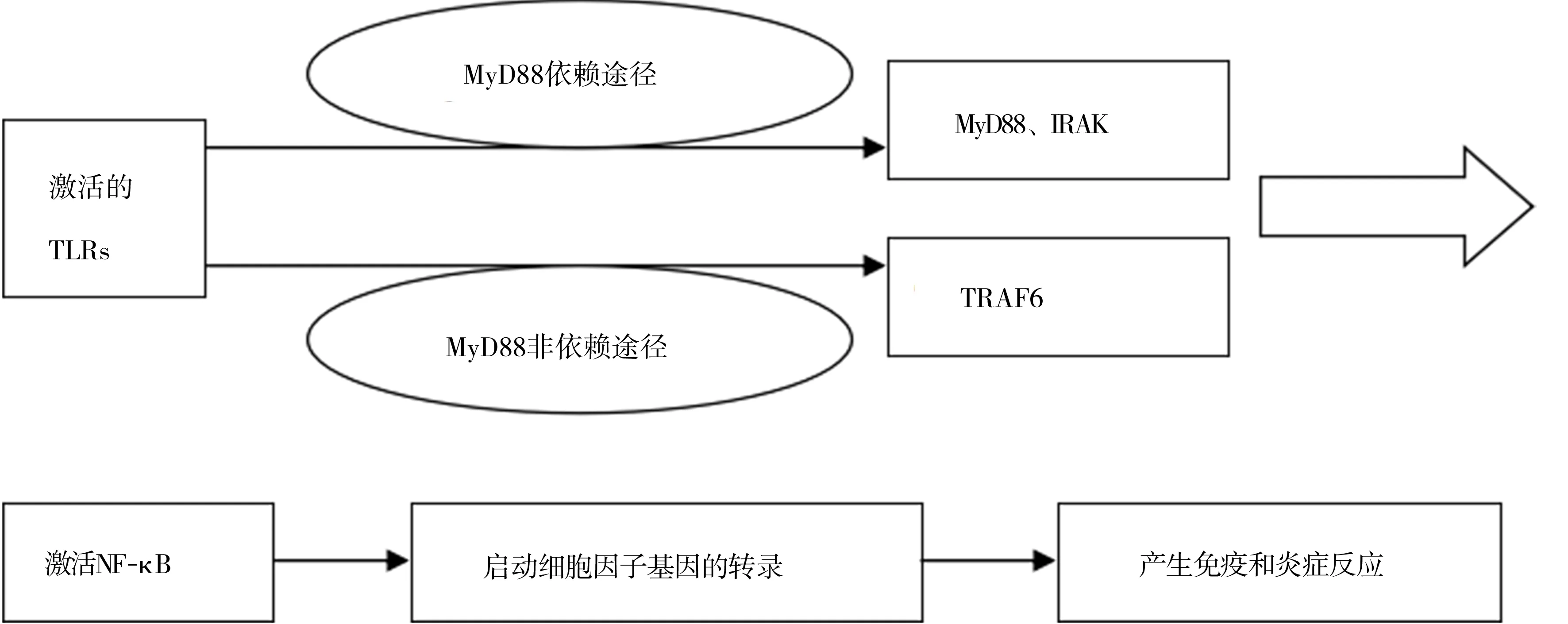
TLRs:Toll樣受體;MyD88:髓樣分化因子88;IRAK:白細胞介素1受體相關激酶;TRAF6:腫瘤壞死因子受體相關因子6;NF-κB:核因子κB
圖1TLRs作用過程
TLRs屬于模式識別受體中的一種,它可通過識別結核分枝桿菌的病原相關分子模式,從而誘導機體產生保護性免疫反應或炎癥反應。10種TLRs中,僅6種(TLR1、TLR2、TLR4、TLR6、TLR8和TLR9)與結核分枝桿菌的識別相關[14]。結核分枝桿菌細胞表面存在與TLRs等受體相互作用的重要配體,包括分子量為19 000~27 000的脂蛋白、38 000的糖脂蛋白、阿拉伯糖甘露聚糖脂和磷脂酰肌醇甘露聚糖[15]。不同的TLRs家族成員可識別不同的成分,如TLR2可識別脂蛋白、阿拉伯糖甘露聚糖脂及磷脂酰肌醇甘露聚糖,TLR4可識別脂多糖,而TLR9可唯一識別細菌DNA中的CpG序列[13,15]。不同的TLRs之間還可形成異源二聚體的組合識別形式,從而擴大對病原相關分子模式的識別范圍,如TLR1/TLR2和TLR2/TLR6[13,29]。TLRs與病原相關分子模式相互作用而啟動細胞內信號轉導通路,通過MyD88依賴途徑或MyD88非依賴途徑最終激活核因子κB,從而使編碼各種細胞因子及受體分子等的基因轉錄,引起多種細胞因子(如IL-1、IL-6、IL-8、IL-12、腫瘤壞死因子α及γ干擾素等)和趨化因子的表達[15,30]。
3 TLRs基因變異與結核易感性
3.1TLR1基因變異與結核易感相關性 TLR1又稱CD281,由胞外區、跨膜區及胞內區組成,各部分氨基酸含量分別為581個、23個和181個,共含786個氨基酸[31]。TLR1可在脾臟及外周血中高表達[31],在機體對抗結核分枝桿菌感染中起重要作用。Hawn等[32]在西雅圖及越南人群中的研究發現,TLR1編碼區存在17種變異[主要是單核苷酸多態性(single nucleotide polymorphism,SNP)],其中,7種屬于同義突變,包括C114T(H38H),A914T(H305L),C944T(P315L),T1583C (C528C),G1677A(P559P),T1760G(V587G),T1892G (L631R),10種屬于非同義突變,包括G1968A(L656L),C2198T(P733L),T130C(S44P),A1482G(V494V),C1938T (H646H),G239C(R80T),C352T(H118Y),A743G(N248S),A1518G(S506S)和T1805G(I602S),非同義突變中有7種在胞外區,2種在胞內區。
研究表明TLR2可分別與TLR1及TLR6形成受體對,從而擴大對病原菌的識別,其胞外區的LRR區可使TLR1/2及TLR2/6受體對多種微生物受體激動劑表現出不同的特異性[33-35]。Omueti等[36]發現,TLR1的LRR區域有3個非同義突變SNPs(rs4833095、rs3923647和rs5743613),其中,rs5743613位點可能會影響許多感染性疾病的固有免疫反應及易感性。研究顯示,TLR1多態性,如rs4833095A和 rs5743618G,可使其在細胞表面表達缺失和功能缺陷,從而極大地影響結核易感性[37]。rs5743611、rs4833095和rs5743618位點變異均為非同義突變SNPs,研究發現,它們與侵襲性曲霉病及克羅恩病的易感性相關[38-39]。據報道,rs5743618G也可降低結核的易感性并增加瘧疾及幽門螺桿菌誘導的胃疾病的發生風險[40-42]。在美國人中,rs4833095、rs5743618、rs3923647和rs5743613位點均可影響抗結核免疫反應及麻風病的易感性[43]。rs4833095位點位于TLR1的跨膜區,可影響受體的轉導和功能,與美國人的結核易感性相關[43]。Dittrich等[10]報道,rs4833095A可降低印度人結核的易感性,并且在體外實驗中它可增強對結核分枝桿菌溶解產物的免疫反應。Sinha等[44]對印度人群的研究也發現,rs4833095A可降低北印度人的結核易感性。rs5743618(602Ile/Ser)是核因子κB信號通路上重要的功能位點,在美國黑人的研究中發現,該位點可影響結核易感性[43];Qi等[45]在中國漢族兒童中的研究也發現,該位點與結核易感性相關;而Ma等[46]研究發現,該位點在中國漢族人中的最小等位基因頻率極低,小于2%,與漢族人的結核易感性之間并不存在相關性;Sinha等[44]也發現該位點的多態性與北印度人的結核易感性不相關。
3.2TLR2基因變異與結核易感相關性 TLR2由784個氨基酸組成,其胞外區含588個氨基酸,跨膜區含21個氨基酸,胞內區含175個氨基酸[47]。在抗結核分枝桿菌感染中,TLR2對巨噬細胞的活化起重要作用[48]。TLR2可識別分子量為19 000的脂蛋白,促進炎性細胞因子的產生,如主要由Th1細胞分泌的腫瘤壞死因子α和γ干擾素[49-52]。免疫反應的激活不僅需要吞噬作用,還需要功能性TLRs的參與。目前,關于TLR2功能性SNPs的報道較少。研究發現TLR2基因多態性可能會降低巨噬細胞對細菌多肽的反應,從而導致機體免疫反應減弱[53]。Texereau等[54]對TLR2的17個SNPs進行研究發現了9個可引起氨基酸改變的非同義突變SNPs。TLR2的胞外區非常重要,可分別與TLR1或TLR6形成異源二聚體,特異性結合不同的配體,從而擴大對微生物的識別譜[33-35],研究者在該區域發現了5個SNPs,其中3個(rs5743708、rs12191786和Pro681His)可通過阻止TLR2與MyD88結合而減弱核因子κB的活化,從而影響細胞信號轉導及炎性細胞因子的產生[54-58]。Pro681His主要出現在亞洲和非洲人群中,在白種人中暫未發現,據報道,該錯義突變與韓國人的結節性麻風及突尼斯人的結核易感性均相關[59-60]。rs5743708和rs12191786是兩個重要的功能位點,目前關于TLR2基因多態性與結核易感性的研究大多集中在這兩個位點。研究顯示,它們分別與土耳其人及突尼斯人的結核易感性相關[60-61]。在中國人群的研究中發現,rs5743708和rs3804099位點與肺結核的發病相關,rs3804100變異在結核潛伏感染者中的頻率高于未感染結核分枝桿菌的健康人群[62-63],Sun等[64]報道,rs5743708位點的多態性與結核易感性相關;而Ma等[46]發現,在中國漢族人中,rs5743708位點的多態性與肺結核易感性之間不相關;rs1898830和rs3804099的多態性與中國漢族兒童的結核易感性也不相關[65]。Wu等[62]在中國人群中進行基因-基因及基因-環境相互作用分析發現,TLR2 rs3804100、TLR2 rs1898830和TLR4 rs10759931交互作用預測肺結核的準確性可達80%。
3.3TLR4基因變異與結核易感相關性 TLR4含839個氨基酸,細菌的脂多糖及脂磷壁酸均可將其活化,脂多糖及脂磷壁酸首先結合在CD14受體簇上,然后轉移給TLR4,形成同源二聚體,再與髓樣分化蛋白2組成復合體,最后運輸到細胞表面[73-74]。TLR4的跨膜區(第632~839位氨基酸)對于其功能性寡聚反應來說至關重要,該區域有一個錯義突變Val651Phe,它可改變TLR4對脂多糖的反應[75]。研究者在LRR區發現了5個低頻的錯義突變(Ser73Arg、Ala97Val、Tyr98Cys、Thr175Ala和rs4986791),這些突變在高加索人中較常見,但在越南人中也可見Thr399Ile頻率的改變[76-77]。一般情況下,氨基酸的改變可能會影響蛋白質的結構和功能,但Tyr98Cys和Thr175Ala錯義突變是否改變了TLR4的功能目前尚不清楚。rs4986790和rs4986791是TLR4基因LRR區的兩個錯義突變位點,它們可影響TLR4胞外結構域,降低對脂多糖的反應,從而減少細胞因子的釋放,使結核分枝桿菌逃避機體的免疫監視,導致感染性疾病易感性增加[78-82]。在坦桑尼亞人群中,rs4986790位點的多態性與人類免疫缺陷病毒感染者的結核易感性相關,但在岡比亞及墨西哥人群中未發現此種相關性[83-85]。研究者在韓國、中國臺灣及日本人群中均未發現rs4986790和rs4986791位點的SNP[86-87]。Wu等[62]在中國人群中的研究發現,rs7873784 G在肺結核患者中的頻率高于健康人,基因-基因及基因-環境相互作用分析顯示,該基因的rs10759932、rs7873784和rs10759931三個位點交互作用預測結核潛伏感染的準確性為84%。
3.4TLR6基因變異與結核易感相關性 TLR6由796個氨基酸組成,僅有1個外顯子[88],它表達于脾臟及外周血白細胞上,可與TLR2形成異源二聚體,識別多種配體,如可溶性結核因子[89],TLR6被激活后,產生細胞信號轉導,核因子κB活化,引起細胞因子分泌及炎癥反應。研究者對中國廣東人的研究發現,在TLR6基因編碼區有7個SNPs,其中5個SNPs是錯義突變[Met59Thr(+176T/C),Ile120Thr(+359T/C),Val327Met(+979G/A),Val465Ile(+1393G/A)和Val470Leu(+1408G/T)],另外2個SNPs(+1083C/G和+1263A/G)沒有氨基酸替換[14]。研究發現非同義突變位點rs5743810的多態性可影響結核易感性[90];Ma等[43]在美國黑人的研究中發現,該位點多態性與結核易感性相關;Sun等[64]研究也發現該位點與結核的發病相關;而Sinha等[44]在北印度人的研究中并未發現此相關性,在中國人群的研究中也未發現該位點的多態性與結核易感性之間存在相關性。
3.5TLR8基因變異與結核易感相關性 TLR8位于X染色體上,可識別病原微生物的單鏈RNA,如RNA病毒[14]。Davila等[11]首次研究了該基因的149個SNPs與印度尼西亞及俄羅斯人群的肺結核易感性之間的相關性,結果發現,在印度尼西亞和俄羅斯男性中,有4個SNPs與肺結核明顯相關,其中,3個SNPs(-129 C/G、-2167 A/G和-1145 A/G)在調節區,1個SNP[rs3764880,1A/G(Met/Val)]在起始密碼子區。攜帶Met/Val的印度尼西亞男性Met等位基因可增加結核的易感性,而Val等位基因可降低結核的易感性[11]。在土耳其兒童中也發現了此種相關性,但未發現-129 C/G SNP與結核易感性相關[91]。Salie等[92]在南非人群中的研究發現,rs3761624、rs3764879和rs3764880三個位點與女性結核易感性相關,而rs3764879和rs3764880兩個位點與男性結核易感性相關。由此可見,TLR8基因的多態性影響疾病易感性可能與性別相關。另外,在中國人群中的研究發現,rs3764879 CC基因型頻率在肺結核患者中的分布明顯高于健康人群,而李彤等[93]在中國東北地區漢族人中的研究并未發現TLR8 SNPs與結核易感性相關。目前,TLR8識別結核分枝桿菌及其胞內信號轉導的機制尚不清楚。
3.6TLR9基因變異與結核易感性相關性 TLR9由1 032個氨基酸組成,含2個外顯子[94],它是哺乳動物細胞識別細菌DNA中的非甲基化CpG序列的主要受體[95]。研究表明TLR9可使樹突狀細胞釋放IL-12,從而抵抗結核分枝桿菌的感染,TLR9缺失的小鼠易感染結核分枝桿菌,而TLR2與TLR9同時缺失的小鼠更易感染結核分枝桿菌[96-97]。rs5743836位于啟動子區域,可能影響TLR9基因的轉錄調節;rs352139位于內含子區域,與印度尼西亞女性的結核易感性相關[98-99];rs187084C可增加印度Baiga部落人群患肺結核的風險[100];但在印度南部人群的研究中發現,啟動子區基因多態性(rs5743836和rs187084)與結核易感性間不存在相關性[101]。另外,Collins等[102]進行基因-基因交互作用分析發現,PTX3 rs2305619、TLR9 rs187084和DC-Sign rs11465421三個位點交互作用,預測結核病的準確度為58.8%。
TLR9的活化對結核分枝桿菌引起的肺肉芽腫性反應的維持至關重要,但其機制尚不清楚[14]。啟動子區的SNPs可通過改變基因轉錄因子的結合潛在地影響基因表達水平,而內含子區域的SNPs可影響mRNA的拼接和(或)增強基因的轉錄[14]。Carvalho等[103]發現,攜帶-1237 TC基因型的外周血單核細胞TLR9和IL-6均高表達,在CpG DNA的刺激下,B細胞的增殖也增加,但機制尚不清楚。
3.7TLR10基因變異與結核易感相關性 TLR10高表達于淋巴組織系統,如淋巴結、扁桃體等,在機體抵抗病原微生物的免疫反應中起重要作用。有研究報道,TLR10與TLR1及TLR6在染色體4p14上有共同的基因座,因此,三者在結構上具有相似性;TLR10可與TLR1或TLR6形成異源二聚體,而TLR1和TLR6均可與TLR2形成受體對,從而擴大對病原微生物的識別,在機體抗炎過程中起重要作用[104];Oosting等[105]發現,TLR10可通過巨噬細胞表面的TLR2起抗炎作用。TLR10 rs4129009A和TLR10 rs10004195T均可增加韓國兒童自身免疫性甲狀腺疾病的發生風險;而TLR10 rs10004195AA基因型則可降低該病的發生風險,TLR10 rs11096956位點的多態性與自身免疫性甲狀腺疾病的發病無相關性[106]。目前,關于TLR10基因多態性與結核易感性之間相關性的研究較少。Bulat-Kardum等[107]報道,TLR10 rs11096957A等位基因可增加克羅地亞人結核的發病風險。
4 結 語
模式識別是宿主抗結核分枝桿菌感染的重要組成部分,它通過TLRs促發抑制機制。TLRs信號間可通過各自的配體進行協調調節,這種協調調節可控制宿主抗結核分枝桿菌感染的免疫反應的程度,在抗結核分枝桿菌的感染中至關重要[14]。TLRs基因的變異(尤其是SNPs)極大地影響了結核的易感性,有獨特TLRs表型的個體可能對結核分枝桿菌配體有或高或低的親和力[14]。因此,需要進一步的深入研究來系統地探索TLRs、接頭蛋白(MyD88、TIR功能區的接頭蛋白、TRIF、 TRAM)和下游分子(如IL-1受體相關激酶和腫瘤壞死因子受體相關因子6)三者之間的相關性,這有助于全面而深入地理解它們的分子生物學特性,評估各種SNPs聯合突變的累積效應。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
