日本結(jié)縷草ZjZFN1基因?qū)M南芥的轉(zhuǎn)化及其耐旱性分析
2019-04-23 07:14:56姜紅巖滕珂檀鵬輝尹淑霞
草業(yè)學(xué)報 2019年4期
姜紅巖,滕珂, 檀鵬輝,尹淑霞*
(1.北京林業(yè)大學(xué)草坪研究所,北京 100083;2.北京市農(nóng)林科學(xué)院,北京草業(yè)與環(huán)境研究發(fā)展中心,北京 100097)
鋅指蛋白是植物中一個轉(zhuǎn)錄因子大家族,主要由半胱氨酸(Cys)和組氨酸(His)組成,結(jié)合鋅離子折疊成手指狀結(jié)構(gòu)的蛋白[1]。 鋅指蛋白被分類為幾個亞家族,例如C2H2,CCCH,C2HC,C2HC4和C5[2]。目前為止,C2H2是植物中鋅指蛋白最豐富的家族之一,在擬南芥(Arabidopsisthaliana)和水稻(Oryzasativa)中分別鑒定出176和189 個C2H2鋅指蛋白[3-4]。這些C2H2鋅指蛋白的功能在模式植物和作物中已經(jīng)得到證明,研究表明它們在植物發(fā)育、植物對生物和非生物脅迫的響應(yīng)中起關(guān)鍵作用。例如,ZAT7或ZAT10的組成型表達(dá)增強了擬南芥對鹽度的耐受性[5-6]。ZAT18[4]和GsZFP1[7]的過表達(dá)增強了擬南芥的干旱耐受性,而GmZFP3[8]負(fù)調(diào)控擬南芥的抗旱性。在鹽芥(Thellungiellasalsuginea)中[9],高鹽、干旱能誘導(dǎo)ThZF1基因表達(dá),在鹽脅迫下擬南芥突變體azf2過表達(dá)ThZF1的生長狀況與野生型擬南芥無明顯差異。越來越多的證據(jù)表明,與許多栽培作物品種相比,一些牧草和草坪草對各種非生物和生物脅迫的耐受性更強[10]。然而C2H2鋅指蛋白在脅迫響應(yīng)中的功能和潛在的轉(zhuǎn)錄調(diào)控機制相關(guān)的基因仍不清楚,尤其是在草坪草中。
日本結(jié)縷草(Zoysiajaponica)是耐旱性最強的草坪草種之一,普遍應(yīng)用于運動場、園林綠化和水土保持等方面[11]。目前為止,對日本結(jié)縷草進行的研究主要集中在評估不同品種之間的非生物脅迫耐受性,報告生理機制以及分子標(biāo)記的發(fā)展[12-13]。由于遺傳資源有限,日本結(jié)縷草耐旱的分子機制尚不清楚。單個C2H2鋅指蛋白的功能研究不僅可以更好地了解其在植物非生物脅迫響應(yīng)中的功能,還可以深入了解植物在脅迫條件下發(fā)生的潛在信號傳導(dǎo)過程。因此,本研究旨在獲得轉(zhuǎn)ZjZFN1基因擬南芥,并對其進行干旱處理,觀察發(fā)芽和生長狀況,并且通過實時熒光定量技術(shù)對干旱處理后轉(zhuǎn)基因擬南芥中抗旱相關(guān)基因的表達(dá)模式進行研究,為進一步探究C2H2型鋅指蛋白ZjZFN1在干旱脅迫中的功能及分子機制提供參考。
1 材料與方法
1.1 材料
本實驗所用日本結(jié)縷草草種為‘Meyer’,由江蘇省中國科學(xué)院植物研究所劉建秀研究員惠贈。實驗所用的野生型擬南芥為本實驗室保存,種植于人工氣候箱(RXZ-380D-LED),培養(yǎng)條件為24/22 ℃(晝/夜),65%濕度,16 h光照。反轉(zhuǎn)錄試劑盒、PrimeSTAR Max DNA Polymerase、pMD19-T載體、SYBR Mix均購自TaKaRa公司;大腸桿菌感受態(tài)DH5α購自北京全式金生物技術(shù)有限公司;2×GoldStar MasterMix購自康為世紀(jì)公司;Seamless Assembly Cloning Kit購自中美泰和生物公司;GUS染色試劑盒購自奧博來公司。
1.2 方法
1.2.1ZjZFN1基因及其啟動子表達(dá)載體的構(gòu)建及擬南芥轉(zhuǎn)化 2017年7月以健康生長3個月的日本結(jié)縷草為材料,利用‘CTAB’方法[14-15]和‘Trizol’方法[16]分別從葉片中提取基因組DNA和總RNA。以提取的DNA為模板,利用染色體步移的方法,以pMD19-T為載體,獲得ZjZFN1啟動子序列。將提取的RNA反轉(zhuǎn)錄為cDNA,并利用RACE技術(shù)進行3′/5′ RACE擴增,以pMD19-T為載體,獲得ZjZFN1基因的ORF。以pMD-ZjZFN1-Pro/pMD-ZjZFN1質(zhì)粒為模板,利用PrimeSTAR Max DNA Polymerase進行PCR擴增,分別用NcoⅠ/Bgl Ⅱ單酶切pCAMBIA1391Z/3302Y載體,通過Seamless Assembly Cloning Kit將純化后PCR產(chǎn)物與酶切產(chǎn)物進行連接。1391Z-ZjZFN1 pro::GUS、3302Y-ZjZFN1載體構(gòu)建成功后,利用凍融法將其轉(zhuǎn)入農(nóng)桿菌GV3101感受態(tài)細(xì)胞中,挑取陽性克隆菌落進行PCR鑒定(表1)。將含有表達(dá)載體的農(nóng)桿菌重懸于侵染液中,使用花序侵染[17]將野生擬南芥的花序浸泡在侵染液中30 s,放于培養(yǎng)箱中暗培養(yǎng)12 h,一周后再次侵染,侵染2~3次。
1.2.2GUS基因表達(dá)檢測 將收獲的轉(zhuǎn)ZjZFN1-Pro的擬南芥種子播種于含有20 mg·L-1潮霉素的MS培養(yǎng)基上進行篩選。擬南芥生長一個月左右,提取擬南芥葉片的DNA,以其為模板,1391Z-ZjZFN1 pro::GUS為陽性對照,野生型擬南芥為陰性對照,進行PCR檢測。將抗性株系繼續(xù)培養(yǎng)并篩選,直至獲得T3代轉(zhuǎn)基因植株,用于GUS化學(xué)組織染色。
將生長2周后的轉(zhuǎn)基因擬南芥移至含有200 mmol·L-1甘露醇的MS培養(yǎng)基上生長,3 d后提取轉(zhuǎn)基因和野生型擬南芥葉片的RNA,并反轉(zhuǎn)錄為cDNA,以其為模板,以qGUS-F/qGUS-R為引物進行實時熒光定量檢測,以擬南芥AtUBQ10基因(NM_116771)作為內(nèi)參基因。采用2-ΔΔCT法[18]計算基因的相對表達(dá)量。
1.2.3轉(zhuǎn)基因擬南芥的篩選及轉(zhuǎn)基因植株的檢測 將收獲的轉(zhuǎn)ZjZFN1的擬南芥種子直接播種于含有草炭、蛭石和珍珠巖(體積比為3∶3∶1)的營養(yǎng)基質(zhì)中,生長兩周后外施60 mg·L-1草銨膦,篩選轉(zhuǎn)基因擬南芥株系。利用CTAB法[14-15]提取轉(zhuǎn)基因型和野生型擬南芥基因組DNA,通過基因組PCR驗證陽性轉(zhuǎn)基因植物,以gDNA為模板,并以3302Y-F/3302Y-R為引物進行PCR鑒定。對表現(xiàn)出對草銨膦100%抗性的株系進行保種并繁殖,直至獲得T3代轉(zhuǎn)基因植株,用于進一步的表型觀察和后續(xù)實驗。
1.2.4轉(zhuǎn)基因擬南芥的干旱處理及抗旱性測定 將轉(zhuǎn)基因擬南芥和野生型擬南芥的種子用70%乙醇和1%次氯酸鈉滅菌,然后分別在含有和不含有200、300 mmol·L-1甘露醇的MS培養(yǎng)基上播種。12 d后,觀察發(fā)芽情況及其表型。為了測定耐旱性,將3周齡幼苗移植到含有草炭、蛭石和珍珠巖(體積比為3∶3∶1)的營養(yǎng)基質(zhì)中,轉(zhuǎn)移至光照培養(yǎng)箱中。對其進行干旱處理21 d后觀察植株表型變化。對干旱處理前和干旱處理第21天時的葉片進行取樣,放于液氮中速凍,之后放于-80 ℃冰箱保存,用于脯氨酸、丙二醛(MDA)含量的測定和抗旱相關(guān)基因表達(dá)分析。
1.2.5轉(zhuǎn)基因擬南芥抗旱相關(guān)基因的表達(dá)分析 利用Trizol法[16]提取上述所取樣品的總RNA,經(jīng)檢驗合格后以其為模板,通過反轉(zhuǎn)錄試劑盒獲得cDNA。通過qRT-PCR技術(shù),對轉(zhuǎn)基因和野生擬南芥中ZjZFN1基因及干旱處理后抗旱相關(guān)的幾個基因抗壞血酸過氧化物酶(ascorbate peroxidase,APX)基因、過氧化物酶(peroxidase,POD)基因、超氧化物歧化酶(superoxide dismutase,SOD)基因、吡咯啉-5-羧酸合成酶(pyrroline-5-carboxylate synthase,P5CS)基因以及晚期胚胎發(fā)生豐富蛋白(late embriogenesis abundant protein,LEA)基因進行實時熒光定量檢測,以擬南芥AtUBQ10基因(NM_116771)作為內(nèi)參基因。采用2-ΔΔCT法[18]計算基因的相對表達(dá)量。
1.3 統(tǒng)計分析
采用WPS中的Excel 2019軟件作圖,采用SPSS 23.0軟件進行統(tǒng)計分析。
2 結(jié)果與分析
2.1 ZjZFN1啟動子活性分析
將轉(zhuǎn)ZjZFN1-Pro::GUS的擬南芥植株進行GUS組織化學(xué)染色,組織化學(xué)染色顯示轉(zhuǎn)ZjZFN1-Pro::GUS擬南芥植株的葉片呈現(xiàn)藍(lán)色(圖1A),表明ZjZFN1的啟動子能夠驅(qū)動GUS基因在葉片中的表達(dá)。在GUS染色結(jié)果的基礎(chǔ)上,用200 mmol·L-1的甘露醇處理轉(zhuǎn)ZjZFN1-Pro::GUS植株,處理3 d后,提取葉片的RNA并反轉(zhuǎn)錄成cDNA,對GUS基因進行熒光定量檢測。熒光定量表達(dá)分析表明,200 mmol·L-1甘露醇處理后GUS基因的表達(dá)水平高于對照中的表達(dá)水平(圖1C)。通過對干旱處理前后的擬南芥植株進行GUS染色(圖1A,B),結(jié)果表明200 mmol·L-1甘露醇處理后的GUS染色強于處理前的染色結(jié)果,與熒光定量結(jié)果相符。
2.2 轉(zhuǎn)基因擬南芥植株的鑒定
通過花序侵染法,轉(zhuǎn)化擬南芥,獲得T0代轉(zhuǎn)基因擬南芥種子。通過將T0代種子播種于營養(yǎng)基質(zhì)中并外施草銨膦,篩選抗性植株。具有抗性的擬南芥植株正常生長,保持綠色,而沒有抗性的擬南芥植株逐漸枯萎死亡。提取具有草銨膦抗性的擬南芥基因組的DNA,進行PCR檢測。PCR產(chǎn)物經(jīng)電泳檢測顯示(圖2),抗性植株DNA的PCR產(chǎn)物與目的條帶大小一致。初步證明,目的基因轉(zhuǎn)化擬南芥成功。獲得T1代轉(zhuǎn)基因植株后,收取T1代種子,繼續(xù)篩選。最終獲得T3代純合轉(zhuǎn)基因株系。因為株系ZFN-2和ZFN-17在36個T 3轉(zhuǎn)基因株系中呈現(xiàn)最高的ZjZFN1轉(zhuǎn)錄水平,所以選擇它們作為表型觀察的代表株系。
2.3 干旱脅迫下轉(zhuǎn)基因擬南芥的生長狀況
將轉(zhuǎn)基因株系ZFN-2、ZFN-17和WT播種于MS培養(yǎng)基上生長,一周之后3種株系之間的發(fā)芽率沒有明顯差異性(圖3A)。而在添加了甘露醇的MS培養(yǎng)基上,轉(zhuǎn)基因株系和WT的發(fā)芽率差異顯著,WT的發(fā)芽率最高(圖3B)。在12 d后,ZFN-2、ZFN-17和WT在MS培養(yǎng)基和添加了200 mmol·L-1甘露醇的MS培養(yǎng)上生長正常,發(fā)芽率基本一致,均長出了幼葉,未含有甘露醇的MS上的擬南芥子葉較大;而在300 mmol·L-1甘露醇的MS培養(yǎng)基上WT的發(fā)芽率明顯高于轉(zhuǎn)基因擬南芥的,僅有WT長出了綠色的子葉(圖3C)。將正常生長3周后的擬南芥移栽到基質(zhì)中生長,干旱處理21 d后,ZFN-2、ZFN-17植株明顯比WT的矮小,葉片失去色澤,萎蔫面積多于WT(圖3D)。

圖1 轉(zhuǎn)基因擬南芥中ZjZFN1啟動子活性分析Fig.1 Activity analysis of ZjZFN1 promoter in transgenic Arabidopsis A,B:轉(zhuǎn)基因擬南芥中的GUS染色 GUS staining in transgenic Arabidopsis.A: 200 mmol·L-1甘露醇處理前 Before treatment with 200 mmol·L-1 mannitol; B: 200 mmol·L-1甘露醇處理后 200 mmol·L-1 mannitol treatment; C: 甘露醇處理后的GUS基因表達(dá)量 GUS gene expression after mannitol treatment. *表示差異顯著(P<0.05),下同。* represent significant difference at P<0.05 with T test, the same below.

圖2 轉(zhuǎn)ZjZFN1基因擬南芥PCR鑒定Fig.2 PCR validation of transgenic A. thaliana CK-: 陰性對照 Negative control; CK+: 陽性對照Positive control; M: Marker; 1~11: 轉(zhuǎn)基因植株Transgenic lines.
2.4 干旱脅迫對轉(zhuǎn)基因擬南芥中脯氨酸、丙二醛含量的影響
脯氨酸是滲透調(diào)節(jié)的重要物質(zhì),在干旱脅迫下發(fā)揮著重要作用。丙二醛是反映干旱脅迫對植物損害的重要指標(biāo),測定轉(zhuǎn)基因和野生型擬南芥葉片中脯氨酸、丙二醛的含量能在一定程度上反映植物的抗旱性[19]。試驗結(jié)果表明,正常生長條件下轉(zhuǎn)基因植株與WT之間的脯氨酸、丙二醛含量沒有顯著差異,而干旱處理21 d后WT中脯氨酸的含量比正常生長條件下的增加了107%,增加的含量分別比ZFN-2、 ZFN-17的多67%、131%(圖4A);而干旱處理21 d后的WT中增加的丙二醛含量比轉(zhuǎn)基因擬南芥ZFN-2、ZFN-17少了20%、56%(圖4B)。
2.5 干旱脅迫下相關(guān)抗性基因的表達(dá)分析
為了探究ZjZFN1基因在轉(zhuǎn)基因擬南芥中的表達(dá)水平,對轉(zhuǎn)基因擬南芥植株 ZFN-2、ZFN-17和野生型擬南芥中的ZjZFN1基因進行qRT-PCR分析,表達(dá)分析表明轉(zhuǎn)基因擬南芥中的ZjZFN1的表達(dá)水平顯著高于野生型擬南芥中的表達(dá)水平(圖5)。為了進一步探究轉(zhuǎn)ZjZFN1基因是否會影響轉(zhuǎn)基因擬南芥中其他抗性相關(guān)基因的表達(dá),對APX、POD、SOD、P5CS、LEA這5種基因進行了qRT-PCR分析。結(jié)果顯示,干旱處理21 d后ZFN-2、ZFN-17中APX的表達(dá)水平均比WT中的提高了59% 左右(圖5);而POD的表達(dá)水平比WT中的分別下降了54%、60%(圖5);SOD、P5CS的表達(dá)水平明顯下降(圖5),SOD的表達(dá)水平比WT中的分別下降了46%、59%,P5CS的表達(dá)水平比WT中的分別下降了74%、75%;LEA的表達(dá)水平比WT中的表達(dá)水平分別下降了13%、25%(圖5)。
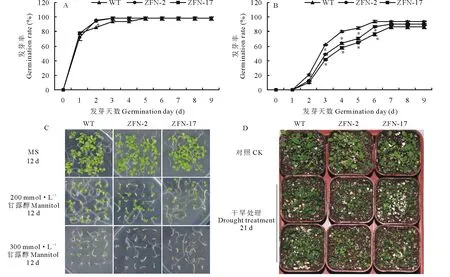
圖3 干旱處理下野生型和轉(zhuǎn)基因擬南芥Fig.3 Drought treatment of wide type (WT) and transgenic A. thaliana A: MS上的發(fā)芽率 Germination rate on MS; B: 在含有200 mmol·L-1甘露醇的MS上的發(fā)芽率 Germination rate on MS containing 200 mmol·L-1 mannitol; C: 在含有不同濃度甘露醇的MS上的發(fā)芽情況 Germination on MS containing different concentrations of mannitol; D: 干旱處理 Drought treatment.

圖4 干旱處理后野生型和轉(zhuǎn)基因擬南芥Fig.4 Wild-type and transgenic Arabidopsis seedlings after drought treatment 不同小寫字母代表單因素方差分析數(shù)據(jù)差異顯著(P<0.05)。下同。The different lowercase letters represent significant differences in one-way ANOVA data (P<0.05). The same below.

圖5 干旱處理下轉(zhuǎn)基因擬南芥相關(guān)抗性基因的表達(dá)Fig.5 Expression of related resistance genes under drought treatment in transgenic Arabidopsis
3 討論
鋅指蛋白是植物中一類重要的轉(zhuǎn)錄因子,目前已經(jīng)在水稻、擬南芥、棉花(Gossypiumhirsutum)、大豆(Glycinemax)等植物中相繼被研究[20]。本實驗從日本結(jié)縷草中分離出一種新的C2H2型鋅指蛋白ZjZFN1,旨在更好地了解其功能和潛在的轉(zhuǎn)錄調(diào)控機制。在前期研究中,我們獲得了ZjZFN1基因及其啟動子的序列。對轉(zhuǎn)ZjZFN1-Pro::GUS擬南芥植株GUS染色后葉片呈現(xiàn)藍(lán)色,結(jié)果表明ZjZFN1的啟動子能夠驅(qū)動GUS基因的表達(dá)。對轉(zhuǎn)ZjZFN1-Pro::GUS擬南芥進行甘露醇處理,熒光定量分析表明甘露醇處理后GUS基因的表達(dá)量顯著增加,這表明ZjZFN1的啟動子上可能存在著干旱響應(yīng)元件。同時,對甘露醇處理后的轉(zhuǎn)基因植株進行GUS染色,結(jié)果表明處理后的染色強于未處理之前的,與熒光定量結(jié)果相符。在此基礎(chǔ)上,通過花序侵染法侵染擬南芥,利用草銨膦篩選擬南芥種子和基因組PCR驗證獲得了轉(zhuǎn)基因擬南芥。目前,C2H2型鋅指蛋白已經(jīng)從不同的物種中相繼被克隆、轉(zhuǎn)化并進行了相關(guān)的功能研究,本實驗通過對日本結(jié)縷草ZjZFN1轉(zhuǎn)化擬南芥探究C2H2鋅指蛋白ZjZFN1的功能。
研究發(fā)現(xiàn),不同物種中C2H2型鋅指蛋白在干旱脅迫響應(yīng)中起著重要作用,多數(shù)植物的C2H2型鋅指蛋白轉(zhuǎn)錄因子在干旱脅迫中起正調(diào)控的作用,提高植物的抗旱性[21-22]。本研究中,在含有甘露醇的MS培養(yǎng)基中生長12 d后,WT的發(fā)芽率高于轉(zhuǎn)基因植株的;干旱處理21 d后,轉(zhuǎn)基因擬南芥相比WT植株矮小、葉片萎蔫。這表明ZjZFN1轉(zhuǎn)錄因子可能在干旱脅迫的種子萌發(fā)過程中起到負(fù)調(diào)控的作用,過表達(dá)ZjZFN1可能降低了植物的抗旱性,這與玉米(Zeamays)中ZmAN13的功能類似[23-24]。研究表明,ZmAN14與ZmAN13具有很高的同源性,而ZmAN14可以提高植物的耐旱性[23],因此不同的鋅指蛋白轉(zhuǎn)錄因子在植物的生長發(fā)育過程中發(fā)揮的功能不同。
為了進一步探究干旱脅迫下ZjZFN1對植物生理功能的影響,本實驗對干旱處理后轉(zhuǎn)基因擬南芥ZFN-2、ZFN-17株系中脯氨酸、丙二醛的含量進行了測定。脯氨酸是植物體內(nèi)的一種重要的滲透調(diào)節(jié)物質(zhì),在干旱脅迫時在植物體內(nèi)會積累大量的游離脯氨酸[25-27]。丙二醛是植物細(xì)胞膜脂過氧化的產(chǎn)物,其含量可表示細(xì)胞膜脂的過氧化程度,在一定程度上反映了干旱脅迫對植物造成的損害[19,25]。兩者均可作為植物抗旱性的重要指標(biāo)。實驗結(jié)果表明,干旱處理后WT、ZFN-2、ZFN-17中的脯氨酸含量都有所升高,而WT中增加的脯氨酸含量高于ZFN-2、ZFN-17中增加的,表明WT的滲透調(diào)節(jié)功能強于轉(zhuǎn)基因植物。干旱處理后WT、ZFN-2、ZFN-17中的丙二醛含量也都有所升高,而ZFN-2、ZFN-17中增加的丙二醛含量均高于WT中增加的,說明轉(zhuǎn)基因植株中的細(xì)胞膜脂損害程度高于WT。干旱處理后植物中脯氨酸、丙二醛含量的變化進一步說明了ZjZFN1在干旱脅迫中可能起負(fù)調(diào)控的作用。
在植物對環(huán)境脅迫的反應(yīng)中,轉(zhuǎn)錄調(diào)控起著主導(dǎo)作用[11]。在干旱脅迫下,植物使用活性氧(Reactive oxygen species,ROS)清除酶(如APX,SOD和POD)以消除ROS積累[28-29]。干旱處理后在轉(zhuǎn)基因植株ZFN-2、ZFN-17中APX的表達(dá)水平比WT中的表達(dá)水平高;而SOD、POD的表達(dá)情況與APX的相反,SOD、POD的轉(zhuǎn)錄水平的提高反映了ZjZFN1過表達(dá)株系中ROS清除能力的增強,這與ZFP179報道的一致[30]。轉(zhuǎn)基因植株中APX與SOD、POD表達(dá)水平的不同表明ZjZFN1過表達(dá)對不同活性氧清除酶的影響不同。P5CS能控制植物體內(nèi)脯氨酸的生物合成以增強滲透脅迫下不同酶的活性,被認(rèn)為是一種重要的抗逆相關(guān)基因[31-32]。干旱處理后轉(zhuǎn)基因植株ZFN-2、ZFN-17中的P5CS的表達(dá)水平低于WT中的表達(dá)水平,表明轉(zhuǎn)基因擬南芥在干旱脅迫中的耐受性降低。LEA是一種逆境脅迫響應(yīng)蛋白[33-34],在干旱脅迫后轉(zhuǎn)基因植株ZFN-2、ZFN-17中LEA的表達(dá)水平低于WT中的表達(dá)水平。通過對這幾種抗旱相關(guān)基因的表達(dá)水平的分析,ZjZFN1的過表達(dá)影響抗旱相關(guān)基因的表達(dá),從而減弱了轉(zhuǎn)基因株系的抗旱性。后期將進行下一步研究,獲得RNAi干擾體系或通過CRISPR技術(shù)將該鹽敏感型基因敲除,從而提高植株的抗旱性。
4 結(jié)論
本研究成功將ZjZFN1基因及其啟動子轉(zhuǎn)化擬南芥,獲得轉(zhuǎn)基因植株。GUS化學(xué)組織染色表明ZjZFN1的啟動子能夠驅(qū)動GUS基因的表達(dá),在甘露醇處理下GUS基因的表達(dá)水平顯著升高。在干旱脅迫下,野生型擬南芥先于轉(zhuǎn)基因擬南芥ZFN-2、ZFN-17株系發(fā)芽;干旱處理21 d后,轉(zhuǎn)基因株系相比野生型擬南芥植株矮小、萎蔫,脯氨酸含量低于野生型擬南芥,而丙二醛含量高于野生型擬南芥。干旱處理下,轉(zhuǎn)基因擬南芥中POD、SOD、P5SC、LED的表達(dá)水平降低,而APX的表達(dá)水平升高。本研究表明ZjZFN1可能減弱植物的抗旱性,為進一步深入研究其功能奠定了基礎(chǔ)。
猜你喜歡
中國民間療法(2021年5期)2021-06-09 09:21:26
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
食藥用菌(2016年6期)2016-03-01 03:24:27
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
護士進修雜志(2015年5期)2015-03-18 19:22:53
中國藥業(yè)(2014年24期)2014-05-26 09:00:09
