水稻LRR-RLK基因LP7的克隆及表達分析
2019-04-29 05:56:44林法明高俊峰李光豪王代長杜長青趙全志
華北農學報 2019年2期
關鍵詞:水稻
林法明,李 珅,王 珂,高俊峰,李光豪,王代長,杜長青, 趙全志
(1.河南農業大學 農學院,河南 鄭州 450046;2.河南農業大學 資源與環境學院,河南 鄭州 450002)
高等植物在逆境脅迫下會感知并傳導逆境脅迫信號,從而啟動植物對逆境脅迫的響應機制,通過調控相關特定功能基因做出各種適應性反應[1-3]。類受體蛋白激酶 (Receptor-like protein kinase, RLK) 是在植物中大量存在的一類跨膜蛋白激酶,在雙子葉模式植物擬南芥中有600個成員,在單子葉模式植物水稻中有1 200多個成員,通常包含胞外配體結合結構域、跨膜結構域和胞內激酶結構域三部分[4-11]。胞外配體結合結構域可感知一些外部逆境信號,進一步通過跨膜傳導,激活胞內的絲氨酸/蘇氨酸激酶結構域,通過磷酸化級聯反應將信號傳遞給下游底物,使信號逐級放大,從而激活下游轉錄因子,誘導抗逆相關基因的表達,開啟脅迫響應應答機制。
富含亮氨酸重復序列的RLK (LRR-RLK)家族是RLK家族中最大的一類亞家族,約占1/2[12-13]。該家族成員最典型的特點是胞外配體結合結構域含有多個LRR,LRR通常是一個由20~29個氨基酸組成的基序,可與配體結合,將胞外信號傳遞至下游,最終使胞內完成響應應答。不同LRR-RLK成員所含有的LRR基序的數目及分布間隔不同,重復次數一般介于3~27次。胞外LRR的多樣性使得LRR-RLK能夠特異地識別不同配體分子,從而正確傳遞不同的胞外信號。近年來,越來越多的證據表明,LRR-RLK成員在調控植物對抗病原菌及各種非生物脅迫中具有十分重要的作用。目前,在植物中已發現并被詳細研究的LRR-RLK成員主要有擬南芥中的BRI1、BKI1、MIK2基因及水稻中的Xa21、LRRK1、DOCS1基因等[14-25],水稻中大部分LRR-RLK基因的功能仍未被揭示。
水稻是我國乃至世界上的主要糧食作物,其產量、品質對我國乃至世界均有重要影響。近年來,水稻生長發育過程中受到低磷脅迫的影響越來越嚴重,降低了水稻的產量和品質。然而,目前針對水稻耐低磷脅迫方面的LRR-RLK研究并不十分深入。因此,為篩選低磷脅迫響應的水稻LRR-RLK基因,河南省水稻生物學重點實驗室對低磷處理的水稻基因芯片表達譜進行分析,發現1個候選LRR-RLK基因的轉錄水平明顯受低磷脅迫誘導。由于該基因為10個候選低磷脅迫響應基因 (命名為LP1-LP10) 中的第7個,因此,將其暫命名為LP7(LOC_Os05g24010),而且其相關研究暫無報道。為此,對LP7基因進行分子克隆、序列比對、亞細胞定位分析,并研究其組織表達模式及低磷脅迫下的表達特征,為深入研究該基因功能奠定理論基礎。
1 材料和方法
1.1 試驗材料及低磷脅迫處理
水稻品種日本晴(Nipponbare)種子及本氏煙草(Nicotianabenthamiana)種子均為河南省水稻生物學重點實驗室保存;瞬時表達載體p35S-GFP由河南農業大學農學院陳鋒課題組趙磊老師提供,由pBI121改造而成。
利用1/2 MS營養液培養水稻幼苗至兩葉一心期,選取整齊度均勻一致的幼苗進行低磷 (10 μmol/L) 脅迫處理,以不進行低磷脅迫處理的材料為對照。處理12 h后,取整株幼苗作為試驗樣品,3次生物學重復。取樣后迅速將樣品置于液氮中冷凍,并保存于-80 ℃超低溫冰箱中備用。
1.2 試驗方法
1.2.1 水稻總RNA的提取及cDNA合成 按照TRNzol總RNA提取試劑盒(TIANGEN)說明書提取水稻不同組織及低磷脅迫處理樣品的總RNA。然后取0.1 μg 總RNA作為模板,按照PrimeScriptTMRT reagent Kit with gDNA Eraser (TaKaRa)試劑盒的操作說明書反轉錄合成cDNA第一鏈。
1.2.2LP7基因的克隆 根據Rice Genome Annotation Project (http://rice.plantbiology.msu.edu/) 網站上公布的水稻LP7基因 (LOC_Os05g24010) 的核苷酸序列,利用Primer Premier 5軟件設計特異引物,上游引物為LP7-F: 5′-ATGGTGAAGAGGCGG CTGTT-3′,下游引物為LP7-R: 5′-CTACGTGCCCCT GTTCTTCA-3′。引物由生工生物工程(上海)股份有限公司合成。以反轉錄合成的cDNA第1鏈為模板進行PCR擴增,50 μL的PCR反應體系包含25 μL Premix PrimeSTAR HS (TaKaRa) DNA聚合酶,上、下游引物 (10 μmol/L) 各2.5 μL,cDNA模板1 μL,ddH2O 補齊至50 μL。PCR反應程序為:98 ℃預變性2 min;98 ℃變性10 s,60 ℃退火10 s,68 ℃延伸 60 s,30個循環;最后68 ℃延伸6 min。PCR產物經1%瓊脂糖凝膠電泳分離,切膠純化回收。回收產物連接至pMD18-T載體,采用熱激法轉化至大腸桿菌DH5α感受態細胞中,篩選陽性克隆送測序。
1.2.3 蛋白質結構及序列分析 利用SMART(http://smart.embl-heidelberg.de/)預測蛋白質結構;利用ExPASy (https://www.expasy.org/)預測蛋白質的分子質量和等電點;利用DNAman 軟件進行多序列比對。
1.2.4 實時熒光定量PCR (qRT-PCR) 分析 利用Primer Premier 5軟件設計qRT-PCR引物,上游引物為LP7-Q-F: 5′-GAGGTGATAATGCAGGACA-3′,下游引物為LP7-Q-R: 5′-TGCGTTGGAGTATGGGT-3′。以水稻OsACTIN基因為內參,其上游引物為:OsACTIN-Q-F: 5′-AACATCGTTCTCAGTGGTGGTA-3′,下游引物為OsACTIN-Q-R: 5′-GGAGGACGGCGATAA CAG-3′,引物均由生工生物工程(上海)股份有限公司合成。利用SYBR?Premix ExTaqTMⅡ(TaKaRa)試劑盒在Bio-Rad CFX96TMReal-time PCR儀上進行qRT-PCR反應。反應體系為:2×SYBR?Premix ExTaqTMⅡ 10 μL,Rox reference Dye Ⅱ 0.2 μL,上、下游引物各2 μL,cDNA模板1 μL (15 ng/μL),ddH2O 補齊至20 μL。反應程序為:95 ℃ 30 s;95 ℃ 10 s,60 ℃ 30 s,35個循環。每個樣品設置3個技術性重復,3個生物學重復。相對表達量根據2-ΔΔCT方法計算[26]。
1.2.5 亞細胞定位 利用Primer Premier 5軟件設計攜帶SmaⅠ酶切位點 (上游引物) 和SpeⅠ酶切位點 (下游引物) 的LP7基因的引物,上游引物為:LP7-GFP-F:5′-CCCGGGATGGTGAAGAGGCGGCTGT T-3′,下游引物為LP7-GFP-R:5′-ACTAGTCGTGCC CCTGTTCTTCAGG-3′。以測序正確的單克隆為模板,PCR擴增LP7基因序列,用SmaⅠ和SpeⅠ進行雙酶切,同時用SmaⅠ和SpeⅠ雙酶切瞬時表達載體p35S-GFP, 分別回收片段,用T4DNA連接酶連接,獲得35S-LP7-GFP融合表達載體,采用熱激法轉化至大腸桿菌DH5α感受態細胞中,用菌落PCR和酶切方法鑒定陽性克隆,并測序驗證。
參照文獻[27]的方法將獲得的重組質粒35S-LP7-GFP轉入農桿菌EHA105菌株中,然后挑取單克隆于5 mL LB液體培養基 (含50 mg/L卡那霉素、100 mg/L利福平)中,28 ℃,180 r/min過夜培養16 h,室溫下4 000 r/min離心5 min,收集菌體。用5 mL注射培養液(10 mmol/L MgCl2、10 mmol/L MES(一水嗎啉乙磺酸)、200 μmmol/L乙酰丁香酮)懸浮菌體,4 000 r/min離心10 min,然后棄上清,最后用注射培養液懸浮至OD600為0.1~0.2,靜置1~2 h,用無針頭的1 mL注射器將其注射至4周齡的本氏煙草葉片中。注射后,25 ℃弱光培養48 h,用激光共聚焦顯微鏡觀察熒光。
2 結果與分析
2.1 LP7基因的克隆及其編碼蛋白序列分析
利用LP7基因序列設計特異引物,以水稻品種日本晴cDNA為模板進行PCR擴增,擴增出1條單一的條帶 (圖1-A)。測序結果顯示,其與Rice Genome Annotation Project (http://rice.plantbiology.msu.edu/) 網站上公布的LP7基因序列完全一致,目的片段長2 832 bp,具有1個完整的開放閱讀框,表明成功克隆了水稻LP7基因。利用SMART軟件分析表明,LP7是典型的LRR-RLK基因家族成員,包含3個典型的結構域:含有2個LRR基序的胞外配體結合結構域、跨膜結構域、胞內激酶結構域(圖1-B)。
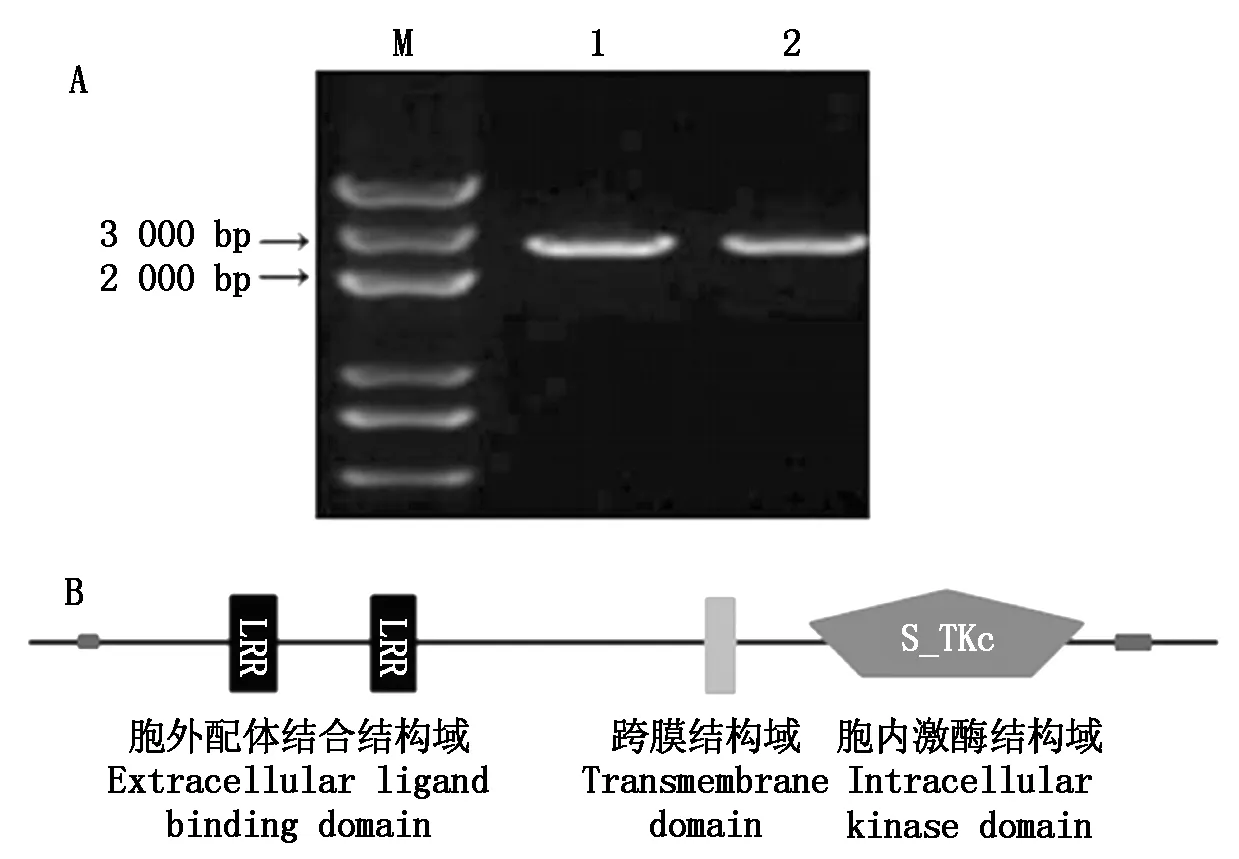
M.DL5000;1-2.LP7基因擴增產物。M.DL5000; 1-2.LP7 gene amplification products.
2.2 LP7蛋白的氨基酸序列比對分析
經ExPASy預測發現,LP7基因編碼蛋白含943個氨基酸,分子質量為103.67 ku,等電點為8.16。將預測的LP7蛋白的氨基酸序列與來自水稻、玉米中的另外2個同源蛋白序列 (EEE63180、NP_001131018) 進行比對 (圖2)發現,LP7蛋白與水稻中的同源蛋白EEE63180的同源性達99%,僅在第75位氨基酸處存在差異,其余序列均完全一致,與玉米中的同源蛋白NP_001131018的同源性達77%,表明LP7基因在單子葉植物中相對比較保守。
2.3 LP7基因的組織表達模式
由圖3可知,LP7基因在水稻根、莖、葉、穗等組織中均有一定量的表達,但在不同組織中的表達水平存在差異,在葉中表達量最高,其次是根,在穗中表達量最低,這可能與植物葉、根在感知逆境信號中發揮重要作用有關。
2.4 LP7蛋白的亞細胞定位分析
由圖4可知,對于轉35S-LP7-GFP載體的煙草葉片,綠色熒光蛋白分布在細胞膜上;而對于轉35S-GFP載體的煙草葉片,綠色熒光蛋白分布在整個細胞膜、細胞質和細胞核中,說明LP7蛋白定位于細胞膜上,這也與SMART在線預測的LP7為一個LRR-RLK的結果一致,暗示LP7作為膜蛋白在接收胞內外信號方面發揮功能。

圖2 LP7蛋白與其他同源蛋白質的氨基酸序列比對結果Fig.2 Multiple amino acid sequences alignment of LP7 and other homologous protein

圖3 LP7基因的組織表達模式分析Fig.3 Expression profile of rice LP7 gene in tissues

圖4 LP7-GFP蛋白與GFP蛋白在本氏煙草葉片細胞中的定位Fig.4 Subcellular localization of LP7-GFP and GFP proteins in leaves of Nicotiana benthamiana
2.5 LP7基因在低磷脅迫條件下的表達分析
由圖5可知,低磷脅迫能明顯誘導LP7基因表達,其表達量較對照增加15倍,表明LP7基因可能在水稻低磷脅迫響應過程中發揮重要功能。

圖5 LP7基因在低磷脅迫條件下的表達Fig.5 Expression level of LP7 gene under low Pi stress
3 結論與討論
RLK在調控植物生長發育過程中具有非常重要的作用。已有研究表明,LRR-RLK作為RLK家族中最大的一類亞家族,在調控植物體生長發育及抗逆脅迫等方面均發揮著十分重要的作用[28-31]。為研究參與低磷脅迫響應的LRR-RLK基因的功能特點,本研究從水稻品種日本晴中克隆到1個LRR-RLK成員LP7基因。氨基酸序列分析表明,LP7蛋白具有典型的LRR-RLK成員特征,為1個LRR-RLK新成員,目前其功能未知。
不同物種的氨基酸序列同源性分析表明,LP7與玉米中的同源蛋白NP_001131018同源性比較高,同源性高達77%,暗示LP7基因在單子葉植物中相對比較保守。
蛋白質在細胞中的定位決定了其行使功能的具體部位。煙草的瞬時表達試驗結果表明,LP7定位在細胞膜上,該結果與已報道的LRR-RLK的亞細胞定位結果一致[28],同時也與LP7作為1個RLK參與細胞外信號接收、傳遞的功能相吻合。
基因組織表達模式分析對研究基因功能十分重要。本研究發現,LP7基因在各組織中均有一定量的表達,在葉中表達量最高,穗中表達量最低。推測可能與植物葉在感知逆境信號中發揮重要作用有關。低磷脅迫條件下的表達分析表明,LP7基因受低磷脅迫誘導表達,與前期芯片數據一致,暗示LP7基因可能在水稻響應低磷脅迫中發揮一定的功能。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
