貝類源抗氧化肽研究進展
2019-04-29 03:31:44楊錫洪渠純純李賽李安奇丁乾李鈺金李麗迪張俊逸解萬翠
食品與機械 2019年3期
楊錫洪渠純純李 賽李安奇丁 乾李鈺金李麗迪張俊逸解萬翠
(1. 青島科技大學海洋科學與生物工程學院,山東 青島 266042;2. 山東省生物化學工程重點實驗室, 山東 青島 266042;3. 農業部冷凍調理海洋食品食點重點實驗室,山東 榮成 264303;4. 榮成泰祥食品股份有限公司,山東 榮成 264303;5. 青島信和源生物科技有限公司,山東 青島 266002)
生物體中活性氧(ROS)和活性氮(RNS)作為在體內發揮重要作用的自由基,參與細胞內信息傳遞過程,并可防御病原體入侵[1]。但過量的自由基會對DNA、蛋白質和膜等各種細胞組分造成氧化損傷。氧化損傷是導致人體衰老和各種疾病如糖尿病、心臟病、中風、動脈硬化和癌癥等的重要因素[2]。研究[3]表明,攝入一定量的抗氧化劑能有效延緩衰老,預防或治療相關疾病。同時,脂質過氧化反應也是造成食品酸敗的主要原因,不僅降低了食物的營養價值,還會產生不良氣味及一些有害物質[4]。抗氧化劑可通過轉移氫原子或電子直接清除自由基,或通過金屬離子螯合作用,抑制自由基的形成,有效阻斷脂質過氧化的鏈式反應[5]。合成抗氧化劑如二丁基羥基甲苯(BHT)、叔丁基羥基茴香醚(BHA)和沒食子酸丙酯(PG)等,雖能有效清除自由基,但其潛在的副作用會對人體產生危害,且成本較高、易產生污染[6]。而天然抗氧化劑具有來源廣泛、成本低且無毒副作用等優點,在越來越多的行業中得到應用。
生物活性肽具備廣泛活性,如抗氧化、抗病毒、抗腫瘤、鎮痛、神經保護、降高血壓和免疫調節等[7]。因此,以生物活性肽作為天然抗氧化劑不僅可阻斷有害的氧化反應,還對人體有其他保護作用。
中國貝類產業發展迅猛[8],貝類及其加工副產物蛋白質豐富,脂肪含量低,是獲取天然抗氧化肽的重要原料[9]。酶解法制備抗氧化肽是目前較為廣泛的方法[10]。隨著現代生物工程的發展,利用基因工程及發酵工程制備貝類源抗氧化肽也取得了一定成果[11]。本文綜述了國內外貝類及其副產物制備抗氧化肽的研究進展,以期對貝類源新資源的高值化利用提供基礎。
1 可控性酶解處理方法及優化
由貝類蛋白制備抗氧化肽通常有化學水解法、微生物發酵法和酶解法等。化學水解法利用酸或堿降解蛋白質獲得活性肽,工藝簡單、成本低,但水解進程不易控制,而且酸堿易導致氨基酸或肽的結構受損,影響生物活性,同時易造成環境問題使其應用受到限制[12];微生物發酵法指利用一定的微生物進行發酵,以此產生的酶能將蛋白質水解從而獲得活性肽,成本較低,但發酵周期長,生成的產物復雜使得提純過程困難[13];酶解法生產肽類具有成本低、條件溫和、產品安全性高等優點,并且水解進程易于控制,廣泛應用于制備貝類源抗氧化肽。
蛋白質的酶促降解受諸多因素影響,使得水解產物的氨基酸組成和分子量大小各異,進而影響肽的抗氧化活性[14]。利用可控酶解技術研究蛋白酶品種、底物濃度、酶用量、pH值、酶解時間及酶解溫度對酶解液水解度、得肽率和抗氧化活性的影響,以獲得最佳酶解工藝條件,能高效制備抗氧化肽[15]。圖1所示為可控酶法制備貝類源抗氧化肽的一般過程。
制備貝類源抗氧化肽的酶解方式主要包括單酶酶解、復合酶解和分段酶解。單酶酶解操作簡單,酶解條件易于控制,但因蛋白酶水解位點高度專一,很難達到預期效果[17]。復合酶解指采用2種或2種以上蛋白酶按照一定比例同時酶解底物,分段酶解指采用多種蛋白酶依次酶解底物,與單酶酶解相比,這2種方法可有效提高酶解效率,但酶解過程中需要控制的條件較多[18]。目前應用于貝類源抗氧化肽生產的常見酶有:動物源蛋白酶,如胃蛋白酶、胰蛋白酶、糜蛋白酶;植物源蛋白酶,如木瓜蛋白酶、菠蘿蛋白酶;微生物源蛋白酶,如枯草桿菌蛋白酶、蠟樣芽胞桿菌蛋白酶等。表1列出了利用海洋貝類及其副產物源制備抗氧化肽的主要方法和結果。
由于蛋白酶具有特定的酶切位點,同一底物經不同蛋白酶酶解后,所得產物的抗氧化活性差異較大[27]。因此,在制備貝類源抗氧化肽時要根據原料及試驗條件挑選合適的蛋白酶,并研究出最佳酶解條件以獲得較多高抗氧化活性的肽。酶解結束后還需對酶解液進行分離、純化、富集及活性檢測,以得到高純度和高活性的抗氧化肽,利于對發揮抗氧化活性組分的進一步研究。
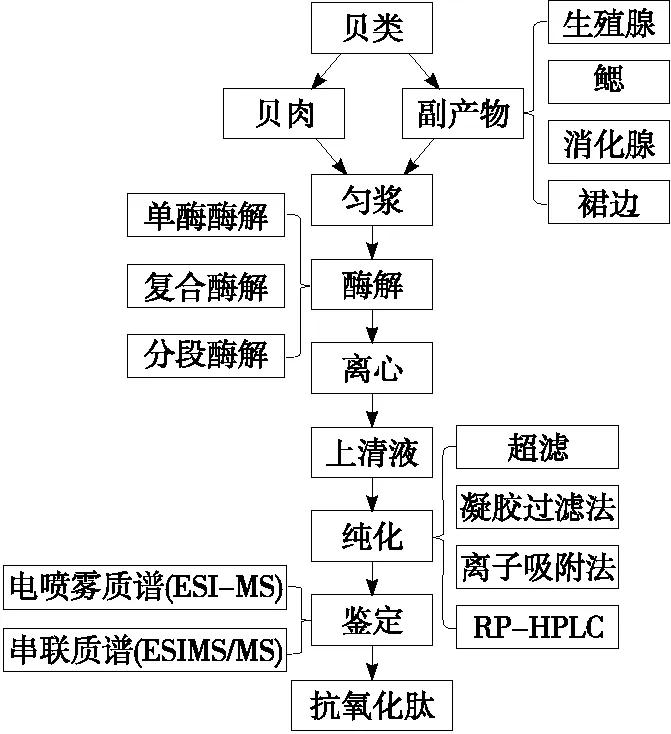
圖1 貝類及其副產品制備抗氧化肽的工藝流程[16]
2 肽的分離純化
研究[28]發現分子量較小的肽往往表現出更高的抗氧化活性。如表1中所示,經不同貝類獲得的抗氧化肽分子量大多處于400~1 500 Da。為獲得具高抗氧化活性的肽組分,需采用一定方法對酶解液中的小分子肽進行分離富集,常用的分離技術包括超濾法、色譜法和色譜—質譜聯用技術。
超濾法常用于抗氧化肽的初步分離,即通過特定截留分子量規格的多孔膜實現對酶解液的分級,如Li等[24]分別利用3,5,10 kDa的超濾膜對短頸蛤水解液進行分離,得到具有不同分子質量范圍的多個肽組分。超濾法操作簡單,無需添加其他化學試劑,但超濾過程中易出現濃差極化使得分離不徹底,常采用色譜法對超濾后的組分進一步純化分離,有時也會同時采用兩種方法以提高分離效果[29]。
色譜法包括離子交換色譜法(IEC)、分子排阻色譜法(SEC)以及反相高效液相色譜法(RP-HPLC)等。其中,IEC又分為陰離子交換色譜(AEC)和陽離子交換色譜(CEC),即以離子交換劑為固定相,依據流動相中的組分與離子交換劑的結合力大小差別達到分離目的。SEC是根據蛋白質和多肽的分子量大小不同,其通過填充柱中的凝膠介質速度不同而實現分離濃縮的,常以Sephadex G-15、Sephadex G-25和Sephadex G-50作為固定相。 如Asha等[21]用30%的乙腈和0.1%的三氟乙酸用作為洗脫液,用Sephadex G-25柱分離了分子量為1~5 kDa的牡蠣酶解液組分,在214 nm處檢測獲得了吸收峰。RP-HPLC分離效率高,通常為純化抗氧化肽的最后一步。體系由非極性固定相和極性流動相組成,常以C18作為固定相,用乙腈或甲醇作為流動相,將洗脫餾分在特定波長下進行檢測,以得到具有最高活性的抗氧化肽組分。Wu 等[25]獲得SEC富集的抗氧化肽后,采用半制備Zorbax SB C18柱,以超純水、0.1%三氟乙酸和甲醇為流動相,在一定洗脫程序下獲得了5組純化的肽,測定各自的抗氧化活性后選擇活性最高的組分用于氨基酸序列測定。
色譜—質譜聯用技術以其自動化和高效的特點,已被廣泛應用于抗氧化肽的分離純化,如氣相色譜—質譜(GC-MS)、液相色譜—串聯質譜(LC-MS/MS)、超高效液相色譜—串聯質譜(UPLC-MS/MS)等。Umayaparvathi 等[20]利用Sephadex G-25凝膠柱獲得牡蠣酶解液中抗氧化活性最高的組分F5后,運用UPLC-MS對其進行分離純化,洗脫收集到的7個肽組分都能有效清除DPPH自由基,其中清除活性最強的3個組分則經 Q-TOF ESI質譜儀獲得了分子量及氨基酸序列(見表1)。Neves等[30]采用UPLC-MS/MS純化并鑒定了鮭魚蛋白酶解液中的十六肽stp-c1的氨基酸序列,發現肽序列Tyr-Pro的氧自由基吸收能力(ORAC)最強。將色譜—質譜聯用技術運用于分離及鑒定抗氧化肽不僅能達到更好的分離效果,還能有效簡化測序過程。
抗氧化肽的分離純化方法多樣,作為結構鑒定前必不可少的一步,研究中應采用適當的分離純化手段逐步得到高純度、高活性的抗氧化成分,以利于對其進行更深入的研究。
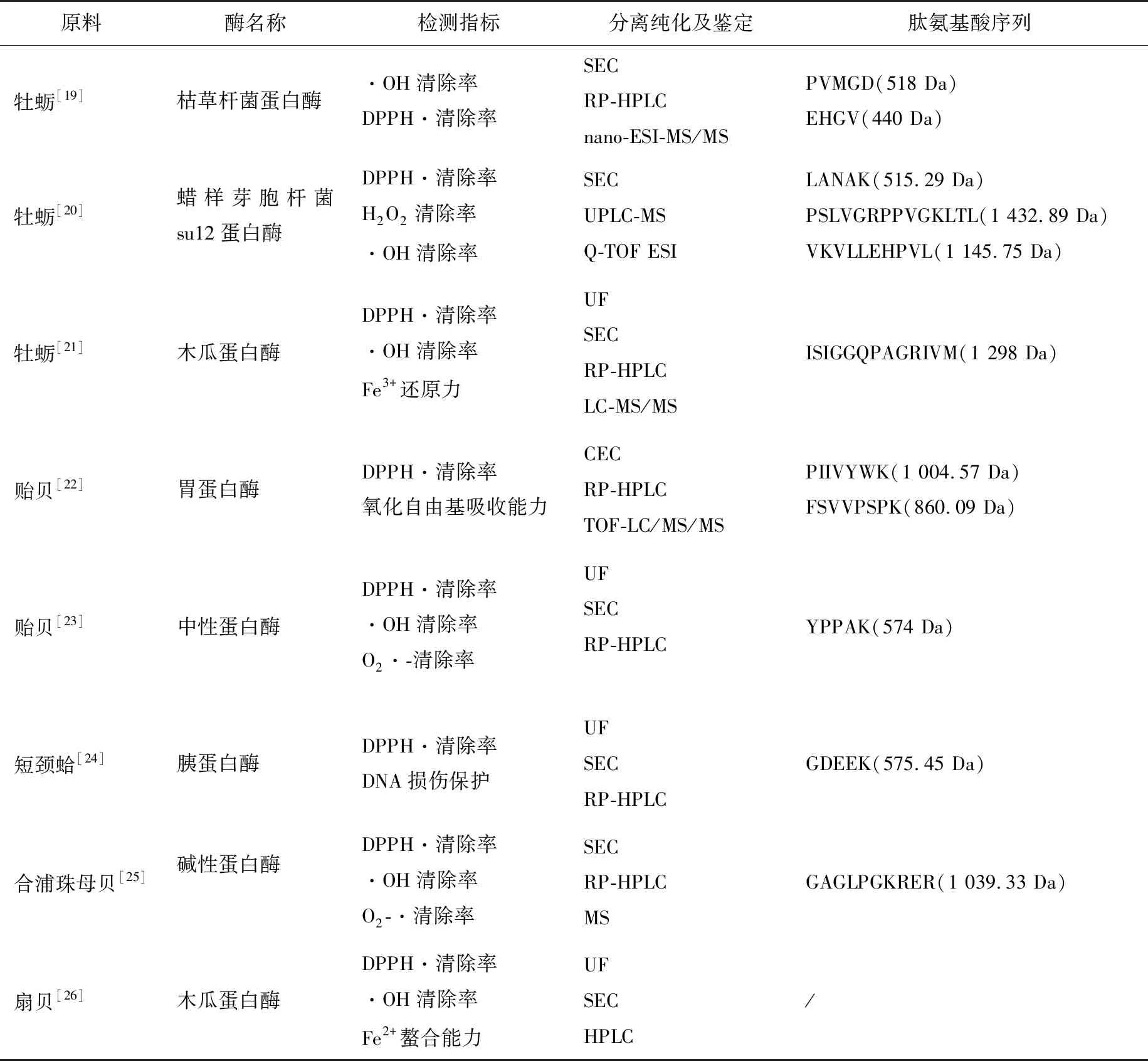
表1 海洋貝類及其副產物源抗氧化肽
3 抗氧化肽活性檢測及結構鑒定
3.1 抗氧化肽活性檢測方法
3.1.1 化學法檢測評價 肽組分在分離純化前,需檢測其抗氧化活性以篩選最佳產物,研究中常測定兩種以上指標相互驗證以表征肽的體外抗氧化活性[31]。肽對各類自由基如·OH、O2·-及NO·等的清除力,最直接地反映了其抗氧化活性;金屬離子螯合能力,即肽與環境中的金屬離子螯合,降低或消除金屬離子對生物細胞毒性危害的能力[32];還原力指肽提供氫原子或電子的能力,具有較高還原力時供電子能力也越強[33]。此外,抗氧化肽的脂質過氧化抑制力也是檢測其氧化活性的一項重要指標。機體內多不飽和脂肪中的碳碳雙鍵(C═C)會降低碳氫鍵的作用力,使氫原子易受自由基攻擊,從而引起脂質過氧化連鎖反應,如圖2所示。含疏水性氨基酸的抗氧化肽因其具有較高的脂溶性,增強了與脂肪酸反應,有效防止氫原子受到攻擊,阻斷脂質過氧化鏈式反應[34]。但生物體內存在著復雜的氧化過程,在體外抗氧化活性較強的肽在生物體內不一定具有同樣的效果。因此,在化學方法評價的基礎上需進一步結合生物試驗進行評價[35]。
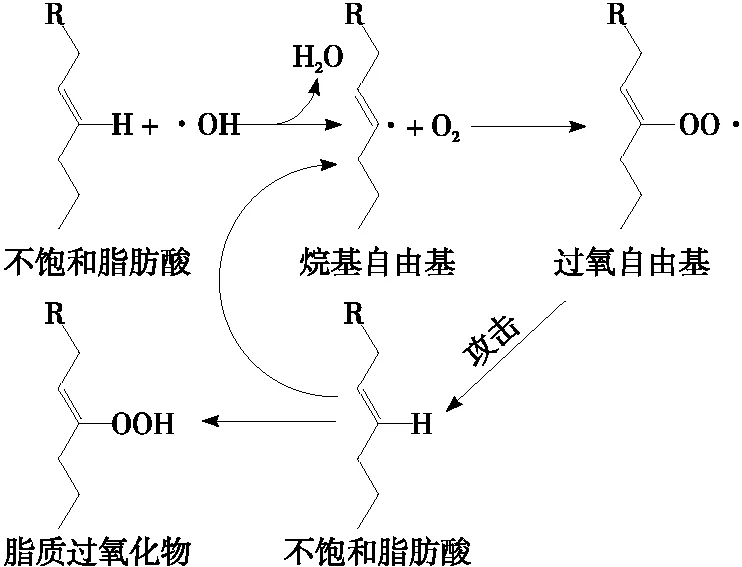
圖2 不飽和脂肪酸氧化過程圖[34]
3.2 肽的分子量及氨基酸序列測定
肽序列組成以及其氨基酸殘基被認為是影響肽抗氧化活性的重要因素[37]。液相色譜—質譜聯用技術(LC-MS/MS)是鑒定多肽序列和分子量的常用方法,通常配備電噴霧電離源(ESI)、聯用混合四級桿飛行時間質譜儀(Q-TOF mass)或三重四極桿—線性離子阱質譜儀對純化的抗氧化肽進行精確測序和分子量測定,與蛋白質測序儀的聯用可提高測序結果的精準性[21]。此外,埃德曼降解法(Edman degradation)也可用于測定肽的氨基酸序列,從蛋白質或多肽氨基末端進行分析。因為此法是以異硫氰酸苯酯對N-端氨基的修飾為基礎,當N-端被其他化學基團所封閉時需要先去除這些基團才能進行測序,且無法得到序列的分子量,有一定的局限性[38]。Chi等[29]對毛蛤分離純化后所得的抗氧化肽段測序,得其序列為Trp-Pro-Pro;Kleekayai等[39]分別從蝦醬水體液得到具有抗氧化作用的肽段,經鑒定的序列為Trp-Pro,如圖3所示。Ongonierma等[40]也提出Trp-Pro具有二肽基肽酶(DPP-IV)抑制活性。Herraiz等[41]也發現含有氨基酸色氨酸Trp的小分子肽能夠提供電子以達到消除陽離子的作用,特別是ABTS自由基。
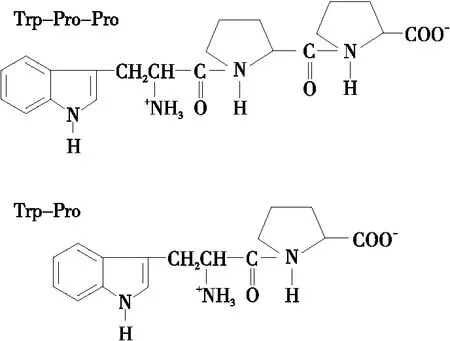
圖3 具有抗氧化活性的氨基酸序列
鑒定具有抗氧化活性肽段的分子量和氨基酸序列,不僅對抗氧化肽構效關系的深層研究具有重要意義,同時對于天然抗氧化肽的體外合成有一定的參考價值,借助這一方法也為合成高活性抗氧化肽能指出新的發展方向。
4 抗氧化肽構效研究
多肽一級結構中氨基酸所含基團、親疏水性、酸堿性及序列,空間構象都會對抗氧化肽活性產生影響。理解結構—活性關系(SAR)有助于預測肽中潛在的抗氧化活性物質,開發新的高活性抗氧化肽[42]。
4.1 一級結構氨基酸組成和序列
氨基酸組成和序列是影響肽抗氧化活性的重要因素[43]。表1給出了近年來部分貝類源抗氧化肽的氨基酸組成與序列,可以看出,這些抗氧化肽的氨基酸序列間同源性甚微,或因其制備原料、水解酶和酶解方式不同造成。研究[44]發現,大部分抗氧化性強的肽除含有Pro、His、Tyr、Trp、Met和Cys等疏水性氨基酸之外,其N端還常含有疏水性殘基。抗氧化肽中疏水性氨基酸的脂肪烴側鏈使其與自由基更易結合,增強肽與多不飽和脂肪酸的反應,從而延緩脂質的過氧化進程[45]。因此,疏水性氨基酸是影響肽抗氧化活性的重要因素。Giménez等[46]研究了3種具有相同分子量的肽:GPLGLLGFLGP-LGLS、GPOGOOGFOGPOGOS和GPOGOOGFLGPOGOS,發現其中疏水性最強的GPLGLLGFLGPLGLS抗氧化活性也最強,進一步證實了疏水性與肽段的抗氧化活性的關聯性較強。含組氨酸的肽可通過供氫、捕獲脂質過氧自由基或利用咪唑基團的金屬離子螯合作用發揮其抗氧化作用[47];肽中的芳香族氨基酸殘基(如酪氨酸、色氨酸和苯丙氨酸)可以向缺電子自由基提供質子從而表現抗氧化性[48]。Wu等[49]從扇貝雌性性腺中分離出2種分子量相似的肽HMSY(536 Da)和PEASY(565 Da),通過對其抗氧化活性的比較證明了該觀點。
此外,在對一些具有脂質過氧化性的貝類源結構鑒定時發現,該類肽具有兩親性。兩親性肽能夠存在于油—水界面,有效猝滅亞油酸乳液體系的水相和油相中的自由基,從而使肽具有抗氧化活性[46]。肽的親脂性和親水性氨基酸殘基可共同作用,促進其與疏水性靶標的結合,從而激發肽的總體抗氧化活性[34]。
以Suetsuna等[44]從酪蛋白水解物中分離出的抗氧化肽Tyr-Phe-Tyr-Pro-Glu-Leu為例,如圖4所示,N端多為疏水性氨基酸(如Val、Leu等),在細胞膜水油層界面處,疏水氨基酸有利于抗氧化肽直接進入細胞質進行ROS清除反應;C端的Cys含易結合自由基(如ROOR、ROO· 、R· 、HO·等)結合的巰基,從而終止自由基鏈式反應[43],發揮抗氧化作用。
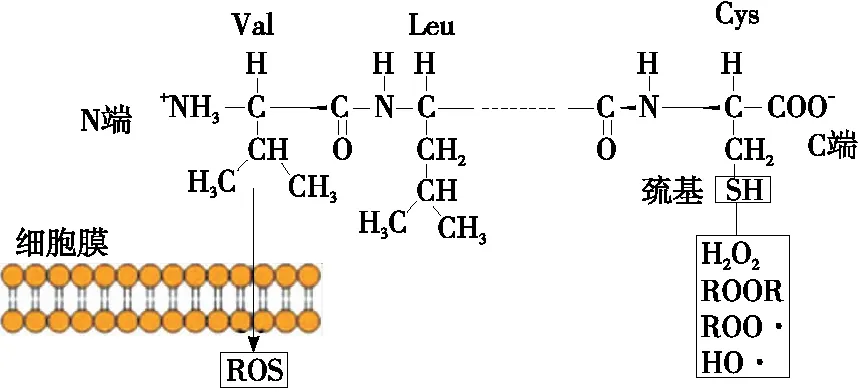
圖4 兩親性抗氧化肽作用機理[43]
4.2 空間構象影響
多肽的空間構象(二級結構、二硫鍵等)也會影響肽的抗氧化活性。Suetsuna等[44]利用蝦肌肉蛋白得到可抑制亞油酸過氧化的肽段IKK、FKK和FIKK,但將相同濃度的氨基酸混合卻無抗氧化活性,說明肽鍵及多肽的結構對抗氧化活性起重要作用。Chen等[50]在對合成肽進行研究時發現,含有組氨酸的人工合成短肽在其單獨存在時,并不具有抗氧化能力,則在一定程度上說明,肽的抗氧化性是所有短肽在一定空間結構上的協同作用產生的。此外,韋緒芹等[51]認為,通過人工合成貝類抗氧化肽的氨基酸序列,采用插入、替換或剔除的方法對肽空間構象進行修飾也會影響其抗氧化活性。
5 結論與展望
作為一種天然活性物質,貝類源抗氧化肽的安全性高、毒副作用小,在生物學、醫學和食品科學領域發揮重要作用,但目前相關研究中仍存在一些問題:酶解法無法制得單一活性肽,且其富集、純化過程耗時較長;活性結構序列測定繁瑣復雜,成本高;抗氧化肽構效關系仍待進一步驗證;活性位點的分布及其作用機理、貝類源抗氧化肽與其他抗氧化劑之間是否存在協同或拮抗作用等問題都值得深入研究。
除抗氧化性外,貝類源肽還具有抗菌性、光保護性和抗光老化活性等其他生物活性,因此被認為是開發藥用化妝品的更安全的選擇。盡管貝類源抗氧化肽在醫學、食品、化妝品等行業的應用潛力很大,但目前多數研究僅處于試驗階段,需要進一步研究其商業化發展的安全性。此外,通過開發一些補充劑,進一步提高貝類源抗氧化肽的生物利用率和組織再生功效,增加其應用潛力,也是一個具有前景的研究方向。
