MeJA對低溫脅迫下冬小麥抗寒生理及關鍵基因表達量的影響
2019-05-15 06:21:22樊曉培羅力力徐慶華
麥類作物學報 2019年4期
趙 虎,樊曉培,羅力力,張 瑞,蒼 晶,徐慶華,于 晶,張 達
(東北農業大學生命科學學院,黑龍江哈爾濱 150030)
低溫限制著全世界農作物的分布及產量。植物對低溫的應答是一個涉及多基因、多信號途徑的復雜過程,植物激素脫落酸(ABA)、水楊酸(SA)、茉莉酸(JA)等參與這個過程,并發揮重要作用[1]。JA及其衍生物茉莉酸甲酯(MeJA)是主要的茉莉酸類物質,可作為信號分子,參與植物響應生物及非生物脅迫。
有研究表明,外施MeJA可以顯著提高低溫脅迫下小麥幼苗超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)等抗氧化酶的活性,增加可溶性蛋白的含量,降低相對電導率和丙二醛(MDA)含量,從而維持細胞膜的完整性,增強小麥抵抗低溫的能力[2]。低溫脅迫增加了小麥[3]、水稻[4]和擬南芥[5]體內JA的水平,促進了擬南芥、水稻JA生物合成基因的表達[4-5],抑制了水稻JA分解代謝相關酶基因的表達[4]。JA信號途徑包括信號產生、轉導及下游基因表達[6],COI1/JAZ/MYC2是JA信號轉導的核心模塊。COI1(Coronatine insensitive 1,冠菌素不敏感因子1)是JA信號轉導途徑的受體。JAZ蛋白(Jasmonate ZIM-domain,茉莉酸ZIM結構域)是轉錄抑制因子,可與MYC2結合抑制JA早期響應基因的表達。MYC2(Myelocytomatosis protein 2,髓細胞組織增生蛋白2)是JA信號途徑的轉錄激活因子,在其信號轉導中起重要作用[7-8]。 PDF1.2(Plant defensin 1.2,植物防衛素)基因編碼一種植物防衛蛋白,研究JA不敏感型和缺陷型突變體發現,擬南芥中AtPDF1.2、AtVSP2(Vegetative storage proteins 2)基因的表達受JAs的誘導及JA信號轉導途徑的調控。
東農冬麥1號是首例能在東北寒冷地區安全越冬的小麥栽培品種,可耐受-30 ℃低溫,返青率高于85%,其強抗寒性得到公認。本課題組前期用東農冬麥1號分蘗節構建了三個低溫脅迫(5 ℃、-10 ℃和-25 ℃)相關的miRNA庫[9],應用生物信息學軟件對其靶基因進行了GO富集分析和KEGG通路分析[10-11],發現植物激素信號轉導途徑中 JA代謝的關鍵基因發生了表達變化。然而,JA是否調控冬小麥低溫脅迫下的抗寒響應以及JA信號轉導途徑中的COI1、MYC2在冬小麥抗寒應答中所起的作用尚不明確,故本研究以東農冬麥1號為材料,探討外源MeJA處理、大田自然降溫條件下,冬小麥越冬期在低溫脅迫下抗寒生理的變化,并在最佳濃度處理下,研究TaPDF1.2、TaCOI1、TaMYC2基因表達量的變化,以期為揭示JA信號途徑關鍵基因在冬小麥響應抗寒中的作用機制、激素JA調控冬小麥抗寒分子機制的研究提供理論依據。
1 材料與方法
1.1 材料與處理
供試材料為冬小麥(TriticumaestivumL.)品種東農冬麥1號,由東北農業大學農學院小麥室提供。試驗于2016-2017年在東北農業大學校內試驗田進行。2016年9月13日播種,完全區組設計,小區面積5 m2(2.5 m×2 m),每個小區種植12行,行距15 cm。每行播種150粒,播深4~5 cm,常規管理。在小麥三葉期(2016年10月1日)分別對葉片噴施濃度為0.5、1.0和2.0 mmol·L-1的MeJA和蒸餾水(含0.02%的酒精)。大田自然降溫且最低氣溫連續10日為5 ℃(2016年10月8日)、0 ℃(2016年10月25日)、-10 ℃(2016年11月9日)、-25 ℃(2017年1月11日)時分別取葉片及分蘗節(地下部分主根以上約1~2 cm左右的白色分蘗密集部位,并避免留取綠色部位),迅速洗凈吸干,剪成1 cm左右大小并分別裝袋,用液氮迅速冷凍后,置于-80 ℃的超低溫冷凍儲存箱中保存備用。
1.2 生理指標的測定[12]
低溫傷害程度的測定采用外滲電導法,丙二醛含量測定采用硫代巴比妥酸法,脯氨酸含量測定采用茚三酮顯色法,可溶性糖含量測定采用蒽酮法,可溶性蛋白質含量測定采用考馬斯亮藍G-250法,SOD活性測定采用NBT法,POD活性測定采用愈創木酚法,CAT活性測定采用紫外吸收法。設置3次生物學重復。
1.3 基因表達分析
基因表達分析采用RT-qPCR 法。根據已得到的TaPDF1.2、TaCOI1和TaMYC2基因(GenBank登錄號分別為NM_123809.4、HM447645.1 和KM382421.1)序列設計實時定量PCR引物,TaPDF1.2基因的正向引物和反向引物分別為5′-CCTTATCTTCGCTGCTCTTGTTC-3′和5′-CT TCTGTGCTTCCACCATTGC-3′,TaCOI1基因的正向引物和反向引物分別為5′-TTGCGGT GAGGAATGTGATTG-3′和5′-TACGGCTAT GGCTGTTAGGC-3′。TaMYC2基因的正向引物和反向引物分別為5′-GATACCCGGGGAACTTG AGCTAG-3′和5′-TCTTCCCGCCGACTTCAT GA-3′。
依次進行總RNA的提取(Trizol法)及基因組DNA的去除、反轉錄,并以反轉錄形成的cDNA為模板進行RT-qPCR檢測。在Mx-3000p Real-Time PCR System(Stratagene,美國)上按照SYBR?Premix Ex TaqTMII(Tli RNaseH Plus)(TaKaRa,日本)說明書進行RT-qPCR。以小麥TaActin為內參基因,其擴增引物序列為5′-CCTTAGTACCTTCCAACAGATGT-3′和5′-CCAGACAACTCGCAACTTAGA-3′。采取2-△△Ct法計算待測基因的相對表達量。
1.4 試驗數據統計分析
用Microsoft Office 2016 Excel作圖,用SPSS 22軟件進行顯著性分析。
2 結果與分析
2.1 MeJA對低溫脅迫下東農冬麥1號抗寒生理的影響
2.1.1 對低溫傷害程度的影響
由表1可知,隨著溫度的降低,處理組與對照組冬小麥葉片和分蘗節的相對電導率均逐漸增大,-25 ℃達到最大值。分蘗節的相對電導率均小于葉片,表明分蘗節的受傷害程度小于葉片,這與分蘗節作為冬小麥主要的越冬器官有關。
5 ℃下,葉片中MeJA各濃度處理組與對照組的相對電導率差異均不顯著,可能是因為MeJA在該溫度下尚未發揮作用;0 ℃、-10 ℃和-25 ℃下,與對照相比,1.0 mmol·L-1的MeJA處理均顯著降低了葉片的相對電導率,而0.5和2.0 mmol·L-1MeJA處理的效果在有些溫度下不明顯。5 ℃和0 ℃下,分蘗節中MeJA各濃度處理組與對照組的相對電導率差異均不顯著,可能是因為MeJA在這兩個溫度下尚未發揮作用;-10 ℃和-25 ℃下,與對照相比,1.0 mmol·L-1的MeJA處理均顯著降低了分蘗節中的相對電導率,0.5和2.0 mmol·L-1MeJA處理的效果在有些溫度下不明顯。說明1.0 mmol·L-1的MeJA處理可以提高小麥的抗寒性。
2.1.2 對丙二醛(MDA)含量的影響
由表1可知,隨著溫度的降低,處理組與對照組冬小麥葉片中MDA含量均呈先下降后上升的趨勢,而分蘗節中MDA的含量均呈逐漸增加的趨勢,-25 ℃均達到最大值。分蘗節的MDA含量低于葉片,表明分蘗節的膜脂過氧化程度小于葉片、抗寒性強于葉片。
5 ℃下,葉片中0.5和1.0 mmol·L-1MeJA處理組與對照組的MDA含量差異不顯著,2.0 mmol·L-1MeJA處理的MDA含量顯著高于對照,分蘗節中MeJA各濃度處理組與對照組的MDA含量差異均不顯著,說明該溫度尚未構成對東農冬麥1號的脅迫,尚未引起JA信號分子對低溫脅迫的響應,此時,高濃度的MeJA處理對小麥葉片反倒有傷害。0 ℃、-10 ℃和-25 ℃下,與對照相比,0.5和1.0 mmol·L-1MeJA處理顯著降低了葉片中的MDA含量,1.0和2.0 mmol·L-1MeJA處理顯著降低了分蘗節中的MDA含量。表明1.0 mmol·L-1外源MeJA處理可以提高冬小麥的抗寒性。
2.1.3 對脯氨酸含量的影響
從表1可知,隨著溫度的降低,處理組與對照組冬小麥葉片及分蘗節中脯氨酸的含量均呈先升高后降低的趨勢,-10 ℃達到最大值。分蘗節的脯氨酸含量大于葉片。與對照組相比,1.0 mmol·L-1MeJA處理均顯著提高了小麥葉片及分蘗節的脯氨酸含量,0.5和2.0 mmol·L-1MeJA處理的效果在有些溫度下不明顯。說明濃度適宜的外源MeJA可能會通過提高脯氨酸的含量來增加細胞的保水力,從而提高其抗寒性。
2.1.4 對可溶性蛋白及可溶性糖含量的影響
從表1可知,隨著溫度的降低,處理組與對照組葉片及分蘗節中可溶性蛋白及可溶性糖含量均呈先增加后降低的趨勢,最大值均出現在-10 ℃。分蘗節的可溶性蛋白及可溶性糖含量均高于葉片。與對照組相比,1.0 mmol·L-1的MeJA處理均顯著提高了冬小麥葉片和分蘗節中可溶性蛋白及可溶性糖的含量,而0.5和2.0 mmol·L-1MeJA處理的效果在有些溫度下不明顯。說明濃度適宜的外源MeJA可以通過提高可溶性蛋白及可溶性糖的積累來提高小麥的滲透調節能力,從而提高其抗寒性。
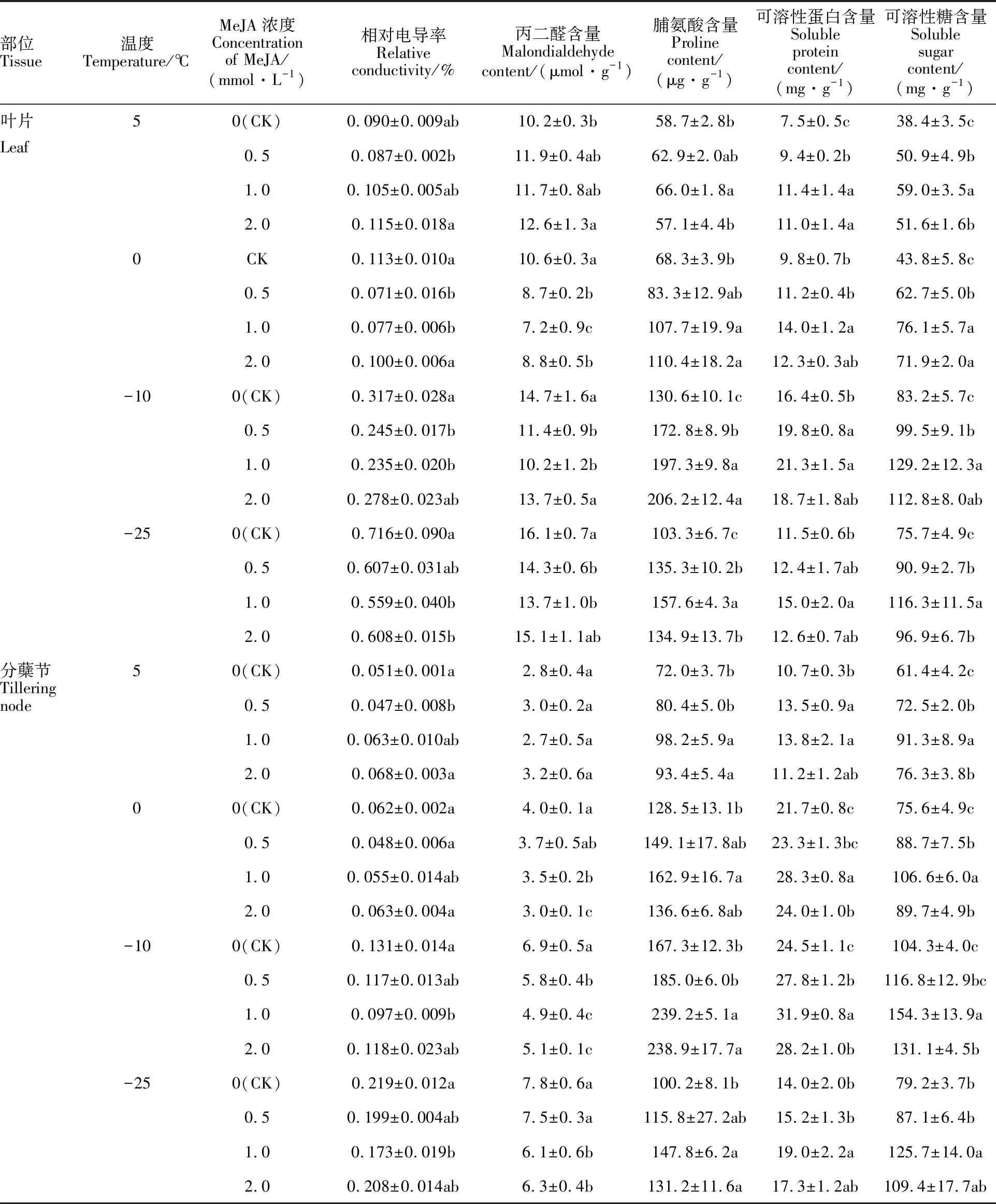
表1 MeJA對低溫脅迫下東農冬麥1號抗寒生理的影響Table 1 Effect of MeJA on cold resistance physiology of Dongnong Dongmai 1 under low temperature stress
同一溫度、同一指標的數據后字母不同代表不同濃度的MeJA處理間差異顯著(P<0.05)。下表同。
Different letters within the same temperature and the same indicator data represent significant differences between different concentrations of MeJA(P<0.05) . The same in table 2 and table 3.
2.2 MeJA對低溫脅迫下東農冬麥1號抗氧化酶活性的影響
由表2可知,隨著溫度的降低,處理組與對照組葉片及分蘗節的SOD活性均呈逐漸升高的趨勢,且最大值均出現在-25 ℃。分蘗節的SOD活性小于葉片,這可能與SOD主要存在于質體有關。與對照組相比,4種低溫下1.0 mmol·L-1的MeJA處理均顯著提高了葉片及分蘗節的SOD活性,而0.5和2.0 mmol·L-1MeJA處理的效果在有些溫度下不明顯。

表2 MeJA對低溫脅迫下東農冬麥1號抗氧化酶活性的影響Table 2 Effect of MeJA on antioxidant enzyme activity of Dongnong Dongmai 1 under low temperature stress
隨著溫度的降低,處理組與對照組葉片及分蘗節POD活性均呈先升高后降低的趨勢,且最大值均出現在-10 ℃。分蘗節的POD活性大于葉片。與對照組相比,4種低溫下1.0 mmol·L-1的MeJA處理均顯著提高了葉片及分蘗節中POD的活性,而0.5和2.0 mmol·L-1MeJA處理的效果在有些溫度下不明顯。
隨著溫度的降低,處理組與對照組葉片及分蘗節中CAT活性均呈先逐漸降低、后略有上升的趨勢,最大值均出現在5 ℃。分蘗節的CAT活性小于葉片。與對照組相比,4種低溫下1.0 mmol·L-1的MeJA處理均顯著提高了葉片及分蘗節中CAT的活性,而0.5和2.0 mmol·L-1MeJA處理的效果在有些溫度下不明顯。
這些結果表明,低溫脅迫下,冬小麥可以通過激活自身的抗氧化防御系統、提高抗氧化酶活性、清除ROS來提高抗寒性。
2.3 MeJA對東農冬麥1號返青率的影響
越冬后,調查了不同濃度MeJA處理下東農冬麥1號的返青率。結果表明,1.0和2.0 mmol·L-1MeJA 處理下的返青率分別為 91.5%和90.7%,顯著(P<0.05)高于對照組的84.7%和0.5 mmol·L-1MeJA處理下的86.5%。
因此,結合2.1節和2.2節的結果,篩選不同低溫脅迫下1.0 mmol·L-1MeJA處理的小麥葉片和分蘗節,進行JA響應基因TaPDF1.2和JA信號轉導途徑關鍵基因TaCOI1、TaMYC2的表達分析。
2.4 MeJA處理對低溫脅迫下東農冬麥1號葉片和分蘗節中 TaPDF1.2、 TaCOI1、 TaMYC2表達變化的影響
從表3可知,隨著溫度的降低,對照組與1.0 mmol·L-1MeJA處理組葉片及分蘗節中TaPDF1.2、TaCOI1、TaMYC2基因的表達量均逐漸增加,-10 ℃達到最大值,此后呈下降趨勢,但-25 ℃仍高于5 ℃,整個過程表現為上調趨勢。與對照組相比,不同溫度下,1.0 mmol·L-1MeJA處理均顯著提高了冬小麥葉片及分蘗節中TaPDF1.2、TaCOI1、TaMYC2基因的表達量(P<0.05);5 ℃下小麥葉片中TaCOI1基因表達量的變化不明顯。
3 討 論
本課題組前期的研究發現,東農冬麥1號在越冬期隨著溫度降低,其體內糖含量逐漸增加;外源ABA處理提高了糖代謝關鍵酶的活性;外源SA處理提高了東農冬麥1號蔗糖含量、蔗糖代謝相關酶的活性及基因的表達[13-14]。本研究發現,隨著溫度的降低,東農冬麥1號葉片和分蘗節中可溶性蛋白、脯氨酸、可溶性糖含量逐漸增加;當溫度降至-25 ℃時,上述滲透調節物質含量降低,表明極寒溫度導致小麥滲透調節機制受損。外源噴施不同濃度的MeJA,均能不同程度提高東農冬麥1號葉片和分蘗節中上述滲透調節物質的積累,其中1.0 mmol·L-1MeJA處理的效果最明顯。這可能與前人發現JAs誘導產生多種化學物質、細胞保護分子[15]有關。
本研究發現,在5 ℃、0 ℃時,東農冬麥1號葉片和分蘗節相對電導率較低,說明其在該低溫下耐受性較好;該溫度下,MeJA處理作用不明顯。隨著溫度降至-10 ℃、-25 ℃時,葉片和分蘗節的相對電導率增大,表明其受低溫脅迫程度變大;該溫度下,1.0 mmol·L-1MeJA處理顯著降低了其相對電導率,表明MeJA提高了膜的穩定性。這與前人的研究結果[16]相一致。隨著溫度的降低,東農冬麥1號葉片和分蘗節MDA含量逐漸升高,表明其膜脂過氧化程度加劇;不同濃度MeJA處理均降低了東農冬麥1號葉片和分蘗節的MDA含量,說明MeJA有助于減輕低溫脅迫對東農冬麥1號膜系統的傷害。
JAs作為信號分子,可激活脅迫響應中的某些信號轉導途徑[17]。有研究表明,0.1 mol·L-1MeJA處理唐菖蒲葉片,可顯著上調GhCOI1基因的表達;處理12 h內,GhCOI1的表達量呈現先上升后下降再上升的趨勢[18]。還有研究表明,適宜濃度的MeJA處理可以促進擬南芥[19]、水稻[20]中COI1基因的相對表達。JA正調控擬南芥的抗凍性,外施JA可顯著提高擬南芥冷馴化誘導的抗凍能力,這與低溫處理誘導擬南芥內源JA合成相關酶的表達、提高內源JA的含量相一致。
本研究發現,1.0 mmol·L-1外源MeJA處理促進了低溫脅迫下東農冬麥1號葉片及分蘗節中TaCOI1、TaMYC2、TaPDF1.2基因的表達,其表達量變化均呈先升后降趨勢,這與前人的研究結果一致。其中TaPDF1.2是JA的響應基因,其表達量增加提示JA含量的增加;而TaCOI1、TaMYC2是JA信號轉導途徑關鍵基因,其表達量的增加可促進下游與抗寒相關基因的轉錄,進而提高植物的抗寒性。這一結果與非生物脅迫可能提高內源JA含量相吻合,也與本課題組前期轉錄組數據顯示的低溫脅迫下TaCOI1、TaMYC2基因的表達量上調相一致。這也提示我們,東農冬麥1號的強抗寒性與ABA、SA、JA等多種激素均有關,處于這些激素互作節點的基因如某些轉錄因子,可能在其中起到關鍵作用。對重要轉錄因子基因的功能分析將是我們下一步關注的重點。
