NLRC5在固有免疫及呼吸系統(tǒng)疾病中的作用
2019-05-15 11:13:56甘聰田曼
國際呼吸雜志 2019年9期
關鍵詞:研究
甘聰 田曼
南京醫(yī)科大學附屬兒童醫(yī)院呼吸科 210008
先天性免疫反應亦被稱為固有免疫反應,是機體有效識別和抵抗外來病原侵襲的第一道防線。模式識別受體是先天免疫反應中的關鍵傳感器,其通過對病原相關分子模式(pathogen-associated molecular patterns,PAMPs)和損傷相關分子模式(damage-associated molecular patterns,DAMPs)的識別,引發(fā)炎性細胞產(chǎn)生促炎因子和趨化因子,誘導免疫細胞向感染部位募集,促發(fā)免疫炎癥反應[1]。目前已知的模式識別受體包括Toll樣受體家族、RIG-I樣受體家族、NOD 樣受體家族及C型凝集素家族[1]。
NOD樣受體的主要結(jié)構域包括:N 端的易變的氨基端效應結(jié)構域、中間端的NOD 域、以及C 端的富含亮氨酸重復序列,各個結(jié)構域其功能上也各有不同。根據(jù)其N 端易變氨基端效應結(jié)構域的不同,NOD 樣受體可分為3個亞家族:NLRCs,其N-末端具有半胱天冬酶激活和募集域(caspase activation and recruitment domain,CARD);NAIPs,其 N-末端具有桿狀病毒抑制劑重復序列(baculovirus inhibitor repeats,BIRs);及NLRPs,其N 末端具有熱蛋白結(jié)構域(pyrin domain,PYD)[2-3](圖1)。
NLRC5是近年來在NLRCs家族中新發(fā)現(xiàn)的成員,已有研究表明NLRC5能參與機體多種免疫反應,在固有免疫系統(tǒng)中具有重要的調(diào)節(jié)作用[4-5],本文將著重總結(jié)近年來對NLRC5的表達、免疫作用和信號傳導途徑,及其在呼吸系統(tǒng)疾病中的作用的研究。

圖1 NOD樣受體家族的結(jié)構[3]
1 NLRC5的結(jié)構
NLRC5由1 866個氨基酸組成,其C 末端包含27個富含亮氨酸重復序列,是所有NOD 樣受體中最長的,因此NLRC5是NOD 樣受體家族中最大的蛋白成員[6]。與其他NOD 樣受體類似,NLRC5 具有典型的三維特征結(jié)構:N端的CARD 結(jié)構域、中間端的NOD 結(jié)構域和C 端的LRR結(jié)構域[3]。其N 端的CARD 結(jié)構域由重復的α螺旋組成,與其他大多數(shù)NOD 樣受體中的CARD 區(qū)和PYD 區(qū)很少有或沒有相似性,因此被稱為非典型CARD 結(jié)構域,亦被稱為死亡折疊區(qū)[7]。與家族其他成員結(jié)構比對表明,NLRC5與MHC Ⅱ類反式激活劑(MHC class Ⅱtransactivator,CⅡTA)密切相關[6]。
2 NLRC5的表達
2.1 NLRC5的細胞內(nèi)表達定位 與其他大多數(shù)NOD 樣受體蛋白不同,NLRC5是核蛋白,但NLRC5的細胞定位顯示,NLRC5在細胞質(zhì)和細胞核中均有表達[6]。研究發(fā)現(xiàn),CⅡTA在其N 和C 末端之間具有幾個核定位信號(nuclear localization signal,NLS),參與細胞核-質(zhì)轉(zhuǎn)運行為,這對其調(diào)節(jié)MHC Ⅱ類基因反式激活至關重要[8]。與之類似的是,NLRC5在其CARD 和NOD 結(jié)構域之間也具有雙向的NLS,NLRC5 介導的MHC Ⅰ類基因表達需要NLRC5核分布和完整的NLS,而NOD 結(jié)構域?qū)LRC5核轉(zhuǎn)位及MHC Ⅰ類基因的反式激活至關重要[9]。用蛋白核輸出抑制劑來普霉素B處理后,NLRC5主要在細胞核中表達,這表明NLRC5可在細胞質(zhì)和細胞核之間穿梭[9]。
2.2 NLRC5的組織表達分布 研究證實NLRC5 在人類和小鼠的多種組織中廣泛表達,尤其在骨髓、淋巴結(jié)、胸腺和脾臟等免疫組織中呈高表達水平[6]。進一步的研究發(fā)現(xiàn),NLRC5主要在造血細胞及免疫細胞中高表達,包括骨髓衍生的巨噬細胞、樹突狀細胞、CD4+T 淋巴細胞、CD8+T 淋巴細胞、B淋巴細胞等小鼠原代細胞及人THP-1細胞、Jurkat T 細胞、Raji B 細胞等[6]。此外,NLRC5在肺、小腸、結(jié)腸和子宮等在具有黏膜表面的組織中高度表達,表明NLRC5可能參與系統(tǒng)免疫信號傳導和黏膜界面宿主防御反應[6]。
2.3 NLRC5的表達調(diào)控 NLRC5可以被各種刺激誘導表達。γ干擾素(interferonγ,IFN-γ)是一種被廣泛報道的可誘導NLRC5 表達的刺激物。在CD4+T 淋巴細胞、CD8+T 淋巴細胞、骨髓巨噬細胞、樹突狀細胞及THP-1、HeLa、CaCo2、HT-29 等多種免疫和上皮細胞系中,IFN-γ能強烈誘導NLRC5轉(zhuǎn)錄水平,此外NLRC5 能被脂多糖(lipopolysaccharide,LPS)、多聚(肌苷酸-胞苷酸)、β干擾素(interferonβ,IFN-β)或病毒感染誘導表達[10]。NLRC5的啟動子包含2 個信號傳導與轉(zhuǎn)錄激活因子1(signal transducers andactivators of transcription-1,STAT1)結(jié)合位點,其中一個與IFN-γ結(jié)合位點重疊,另一個與核轉(zhuǎn)錄因子κB(nuclear factorκB,NF-κB)結(jié)合位點重疊[11]。在Stat1-/-小鼠骨髓衍生的巨噬細胞和T 細胞中,用干擾素或可誘導干擾素產(chǎn)生的物質(zhì)處理后,NLRC5表達顯著減少,由此可見STAT1可能在NLRC5表達的調(diào)節(jié)中起重要作用[12]。此外,研究發(fā)現(xiàn),用雙鏈DNA 巨細胞病毒感染人包皮成纖維細胞后,NLRC5 m RNA 表達顯著上調(diào),繼而應用JAK/STAT 途徑的化學抑制劑和IFN-γ中和抗體處理細胞后,NLRC5表達上調(diào)均被消除,這表明巨細胞病毒誘導的NLRC5表達上調(diào)涉及IFN-γ的自分泌,及IFN-γ受體介導的JAK/STAT 信號途徑的激活[11]。總的來說,NLRC5的表達受多種因素的調(diào)控,可能與IFN-γ及其介導的JAK/STAT 信號途徑相關,但更多的調(diào)控途徑有待進一步研究發(fā)現(xiàn)。
3 NLRC5在固有免疫反應中的作用
機體的固有免疫反應主要由樹突狀細胞、巨噬細胞、自然殺傷細胞、肥大細胞、中性粒細胞等固有免疫細胞介導,通過PAMPs或DAMPs激發(fā)促炎信號轉(zhuǎn)導途徑、Ⅰ型干擾素反應和炎癥小體活化,在防止炎性疾病及免疫性疾病的發(fā)生中具有重要地位[13]。現(xiàn)有關于NLRC5的免疫調(diào)控作用研究也多集中在上述幾個方面。研究發(fā)現(xiàn),NLRC5能調(diào)控NF-κB 炎性傳導 通路、Ⅰ型干擾素(Type Ⅰinterferon,IFN-Ⅰ)介導的炎癥反應及炎癥小體激活,尤其對MHC Ⅰ類基因的表達調(diào)控具有重要作用。
3.1 NLRC5調(diào)節(jié)MHC Ⅰ類基因表達 MHC Ⅰ是一類可以與幾乎所有有核細胞中內(nèi)源性抗原肽結(jié)合的細胞表面受體,能被自然殺傷細胞的抑制性受體識別并抑制其細胞毒性作用[10]。此外,抗原蛋白被蛋白酶水解成的抗原肽后,能通過與MCH Ⅰ類分子結(jié)合并轉(zhuǎn)移至抗原呈遞細胞表面,介導MHC Ⅰ類限制性CD8+T 細胞活化,對宿主防御病原體感染和抗腫瘤免疫反應至關重要[14]。研究發(fā)現(xiàn),在過表達NLRC5可以誘導各種MHC Ⅰ類家族基因及與抗原加工呈遞相關的基因的表達,且NLRC5所有結(jié)構域是MHCⅠ類基因誘導所必需的[15]。已知MHC Ⅱ類基因啟動子上順式調(diào)控元件X1-box、X2-box和Y-box分別與轉(zhuǎn)錄調(diào)節(jié)因子X 復合物、環(huán)磷酸腺苷反應元件結(jié)合蛋白/轉(zhuǎn)錄活化因子1、核轉(zhuǎn)錄因子Y 蛋白復合物結(jié)合形成大分子蛋白復合物,并與CⅡTA結(jié)合形成MHC Ⅱ類增強體,從而調(diào)節(jié)MHC Ⅱ類基因表達[10,16](圖2)。而MHC Ⅰ類和MHCⅡ類基因在其啟動子區(qū)域都具有保守的順式調(diào)控元件X1-box、X2-box和Y-box,且NLRC5 與CⅡTA結(jié)構上具有高度相似性,這表明NLRC5介導的MHC Ⅰ類基因反式激活機制可能與CⅡTA介導的MHC Ⅱ類基因激活機制相似[10,17](圖3)。此外,NLRC5 可能影響組蛋白甲基化,表明NLRC5可能通過調(diào)節(jié)MHC Ⅰ類基因座的染色體活化狀態(tài)來影響基因表達[15]。
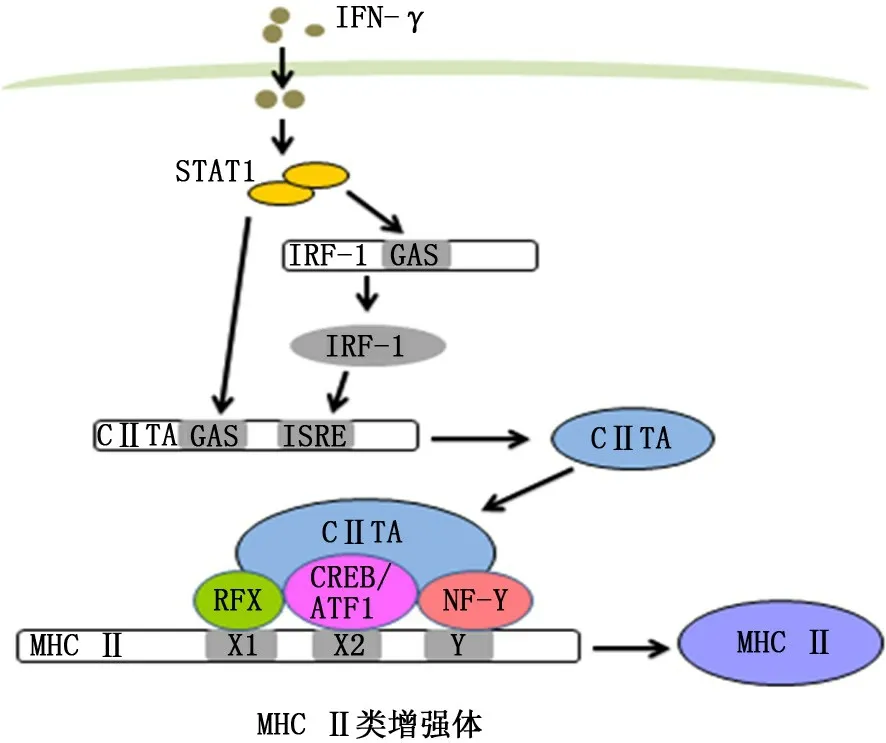
圖2 MHC Ⅱ類增強體轉(zhuǎn)錄調(diào)節(jié)MHC Ⅱ類基因模型[10]
3.2 NLRC5是NF-κB的負調(diào)控因子 NF-κB 轉(zhuǎn)錄因子家族是先天免疫應答的關鍵組成部分,多種模式識別受體可通過對PAMPs的識別,激活下游NF-κB 信號途徑,參與固有免疫反應的促炎信號傳遞[18]。有研究認為,NLRC5是Toll樣受體4信號的負調(diào)節(jié)因子,可以直接與NF-κB抑制激酶α和β相互作用,并阻斷其磷酸化,從而抑制NF-κB活化[19]。Benko等[6]發(fā)現(xiàn)NLRC5 的過表達可能通過影響p65 NF-κB核轉(zhuǎn)位來抑制下游NF-κB信號傳導。此外,NLRC5的CARD可直接與RIG-Ⅰ樣受體的CARD相互作用并抑制NF-κB的激活[19]。
3.3 NLRC5是IFN-Ⅰ的負調(diào)控因子 外來病原入侵機體后可被模式識別受體識別,進而誘導機體產(chǎn)生IFN-Ⅰ應答,并形成抵抗病原入侵的宿主防御反應,在固有免疫反應中發(fā)揮重要作用[20]。IFN-Ⅰ家族包含13~14 個功能性IFN-α基因、IFN-β和幾種其他亞型,具有抗病毒、抗增殖和免疫調(diào)節(jié)等重要作用[21]。研究發(fā)現(xiàn),NRLC5 可與RIG-Ⅰ和MDA5相互作用,有效抑制RIG-Ⅰ樣受體介導的IFN-Ⅰ反應[19]。轉(zhuǎn)染siRNA 介導的NLRC5沉默導致巨細胞病毒和仙臺病毒感染后IFN-Ⅰ的產(chǎn)生分泌減少[11,22]。用LPS攻擊或水皰性口炎病毒感染NLRC5 缺陷型小鼠,其血清中IFN-β含量增加[23]。進一步,在LPS刺激或水皰性口炎病毒感染后的NLRC5缺陷型小鼠胚胎成纖維細胞、腹腔巨噬細胞和骨髓來源的巨噬細胞中IFN-β的產(chǎn)生也顯示增加,然而NLRC5缺陷型小鼠骨髓來源的樹突狀細胞中沒有發(fā)現(xiàn)IFN-β增加這表明NLRC5對IFN-β的負調(diào)控作用可能存在細胞類型特異性[23]。
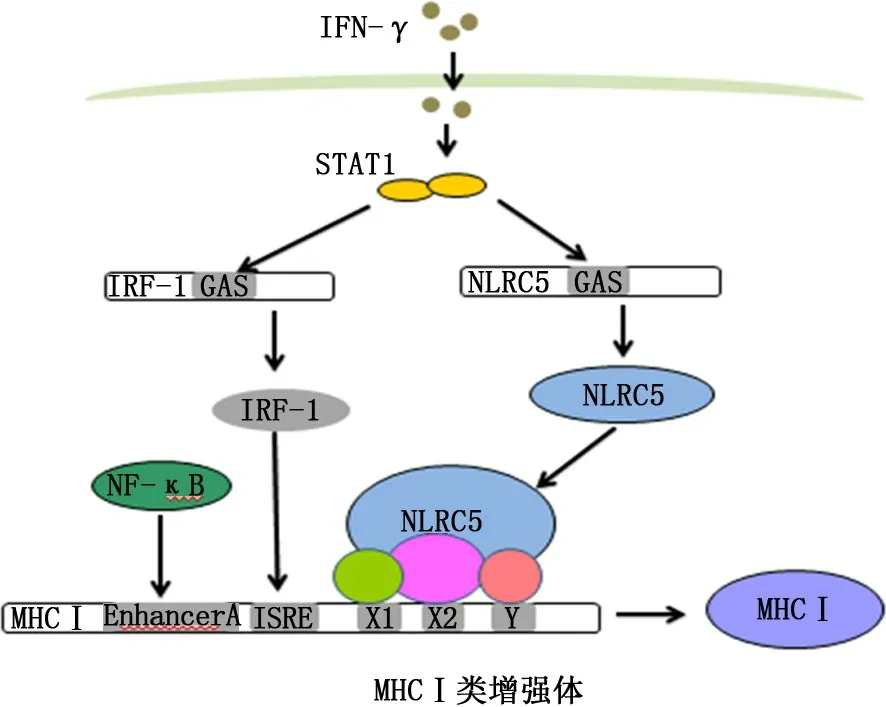
圖3 MHC Ⅰ類增強體轉(zhuǎn)錄調(diào)節(jié)MHC Ⅰ類基因模型[10]
3.4 NLRC5是炎癥小體的激活劑 炎癥小體是一組能識別PAMPs和DAMPs的細胞內(nèi)模式識別受體大分子復合物,其被認為是介導固有免疫反應的重要平臺,能夠調(diào)節(jié)半胱天冬酶1(caspase-1)的活化,促進IL-1β 前體和IL-18前體的切割成熟,并能激發(fā)高炎癥細胞凋亡[2]。先前的研究表明NLRP1和NLRP3是炎癥小體的激活劑[24]。在最近的研究中NLRC5 也被提出作為炎癥小體的激活劑。在293T 細胞中,NLRC5 的過表達增加 了caspase-1 的活化,從而增加IL-1β的產(chǎn)生和釋放[10]。此外,體外研究發(fā)現(xiàn),在人類骨髓THP-1細胞和原代單核細胞中NLRC5基因表達沉默后,大腸桿菌感染導致的caspase-1 活化和IL-1β成熟、分泌減少[4]。
4 NLRC5與呼吸系統(tǒng)疾病
對病原體的識別和炎癥反應的激活是呼吸系統(tǒng)疾病中免疫反應的重要組成部分。NLRC5作為NLRs的一員,能調(diào)控MHC Ⅰ分子表達、激活NF-κB 信號通路,誘發(fā)炎癥因子產(chǎn)生,與呼吸道病毒感染、支氣管哮喘、急性肺損傷、肺纖維化和肺癌等疾病的發(fā)生發(fā)展密切相關。
4.1 NLRC5與流行性感冒 Ranjan等[25]研究發(fā)現(xiàn),在人肺上皮A549細胞系和正常人支氣管上皮細胞中NLRC5的過表達可抑制流感病毒復制,并能誘導IFN-β表達增強,其進一步的區(qū)域作用研究表明,NLRC5 通過其N-末端死亡結(jié)構域與RIG-Ⅰ相互作用以增強抗流感病毒感染的免疫應答。Lupfer等[26]研究發(fā)現(xiàn),NLRC5 基因敲除小鼠在甲型流感病毒感染之前即表現(xiàn)出MHCⅠ表達減少及CD8+T細胞數(shù)量減少,在甲型流感病毒感染的情況下,NLRC5基因敲除小鼠出現(xiàn)CD8+T 細胞功能降低和病毒清除受損。這表明,NLRC5對于促進流感病毒感染期間的宿主防御具有重要作用。
4.2 NLRC5 與支氣管哮喘(哮喘) Guo 等[27]研究發(fā)現(xiàn),RSV 感染后,A549氣道上皮細胞中RIG-Ⅰ表達上調(diào),導致干擾素調(diào)節(jié)因子3活化和IFN-β分泌,并反過來誘導A549細胞中的NLRC5 表達,從而上調(diào)MHCⅠ表達,促進細胞毒性T 細胞活化和抗病毒免疫,這可能與嬰幼兒RSV 感染致細支氣管炎,并誘發(fā)成年后持續(xù)性喘息和過敏性哮喘有關。國內(nèi)也有研究發(fā)現(xiàn),RSV 感染可過誘導干擾素釋放和病毒雙鏈RNA 顯著上調(diào)NLRC5的表達,且抑制NLRC5的表達后能降低RSV 感染上調(diào)的MHCⅠ表達,這可能是RSV 所致免疫病理損傷及誘發(fā)哮喘的潛在原因[28]。Triantafilou等[29]發(fā)現(xiàn),鼻病毒通過其離子通道2B 蛋白觸發(fā)支氣管細胞NLRP3和NLRC5炎性體激活,并誘導IL-1β的分泌增加,從而可能誘發(fā)哮喘的急性發(fā)作。
4.3 NLRC5與急性肺損傷 早前國內(nèi)已有文獻報道,在腹腔注射LPS建立的小鼠急性肺損傷模型中,NLRC5 的表達顯著上調(diào),這表明NLRC5可能參與急性肺損傷的發(fā)病[30]。近期,國內(nèi)也有學者發(fā)現(xiàn),在氣管滴注LPS誘導建立小鼠急性肺損傷模型中,小鼠肺組織和肺泡巨噬細胞中NLRC5的表達明顯增高,在體外LPS誘導刺激的肺泡巨噬細胞中過表達NLRC5 時,腫瘤壞死因子α、IL-1β 和IL-6的表達下降,NF-κB 和p38 MAPK 信號通路也受到抑制,而沉默NLRC5后腫瘤壞死因子α、IL-1β和IL-6的表達上升,NF-κB 和p38MAPK 信號通路磷酸化上調(diào),這些結(jié)果提示NLRC5 能夠負調(diào)控肺泡巨噬細胞腫瘤壞死因子α、IL-1β和IL-6的的表達,其機制可能與NLRC5負調(diào)控NF-κB 和p38 MAPK 信號通路密切相關[31]。
4.4 NLRC5與其他呼吸系統(tǒng)疾病 國內(nèi)研究發(fā)現(xiàn),在博來霉素誘導大鼠肺纖維化模型中,大鼠肺組織中NLRC5的表達增加,且其表達量隨時間的延長逐漸增加,提示NLRC5可能參與肺纖維化形成的整個過程[32]。此外,在非小細胞肺癌患者中,NLRC5的表達與MHC Ⅰ類的表達相關,且核NLRC5陽性和MHC Ⅰ類陽性的Ⅲ期非小細胞肺癌患者總生存期發(fā)生率較低,這提示NLRC5和MHC Ⅰ類可能是Ⅲ期非小細胞肺癌患者的陰性預后指標[33]。
5 總結(jié)與展望
NLRC5作為NOD 樣受體家族最大成員,通過模式識別參與機體先天性免疫反應。已有多項研究表明,NLRC5能特異性調(diào)節(jié)MHC Ⅰ類基因表達、參與MHC Ⅰ類抗原呈遞的相關基因的轉(zhuǎn)錄,并調(diào)節(jié)CD8+T 細胞活化以抵抗細胞內(nèi)病原感染的宿主防御。NLRC5 可能抑制NF-κB 的活化并負調(diào)控IFN-Ⅰ產(chǎn)生。此外,NLRC5 可能作為激活劑促進炎癥小體的活化。
近年來,國內(nèi)外已有文獻總結(jié)NLRC5 在固有免疫中的重要作用,但缺乏NLRC5 與相關系統(tǒng)疾病的總結(jié)。鑒于MHC Ⅰ類抗原提呈、NF-κB 信號通路的激活和炎癥小體的活化同樣是呼吸系統(tǒng)疾病中免疫反應的重要組成部分,近年來不斷的研究表明,NLRC5 與呼吸道病毒感染性疾病、哮喘、急性肺損傷、肺纖維化、肺癌的發(fā)病密切相關。免疫炎癥長期以來一直是呼吸系統(tǒng)疾病的研究熱點,NLRC5的發(fā)現(xiàn)和研究不僅拓展了呼吸系統(tǒng)免疫研究領域,也給此類疾病的發(fā)病機制與治療帶來了新視角。通過靶向改變NLRC5的表達,我們可以干擾MHC Ⅰ表達、NF-κB信號通路和炎癥小體的活化,進而減輕炎癥反應。此外NLRC5在肺纖維化和肺癌中還具有成為潛在的診斷和預后指標可能。因此,在呼吸系統(tǒng)方面,NLRC5的未來研究價值是巨大的。但目前關于NLRC5在呼吸系統(tǒng)疾病中的作用研究多集中在細胞和動物研究,仍缺乏足夠的臨床研究,且關于NLRC5在免疫信號調(diào)控途徑方面的作用尚不完全清楚,NLRC5的作用和應用還有待進一步探索和拓展。
利益沖突所有作者均聲明不存在利益沖突
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
