理解雙人肢體運動表達的友好和敵對意圖的ERP證據*
2019-05-17 02:18:58黃志華王益文
心理學報 2019年5期
黃 亮 楊 雪 黃志華 王益文
?
理解雙人肢體運動表達的友好和敵對意圖的ERP證據
黃 亮楊 雪黃志華王益文
(閩南師范大學應用心理研究所, 漳州 363000) (福州大學心理與認知科學研究所, 福州 350116) (福州大學中國社會信任研究中心, 福州 350116) (福州大學數學與計算機科學學院, 福州 350116)
已有的意圖理解神經成像研究大多關注理解單一個體的中性或負性意圖的腦功能定位, 而大腦理解依靠雙人肢體運動表達的友好和敵對意圖的動態時間過程尚不清楚。本研究記錄了20名健康被試完成三種不同意圖推理任務時的腦電成分。三種意圖分別為:1)友好意圖; 2)敵對意圖; 3)中性無互動意圖。行為結果發現理解敵對意圖的反應時最短。電生理學結果表明, 在額中區的N250 (170~270 ms)上, 中性意圖比友好和敵對意圖均更負, 且友好意圖比敵對意圖也更負; 在大腦右半球的P300 (270~450 ms)上, 敵對意圖比友好和中性意圖均更正, 且友好意圖比中性意圖也更正。對友好和敵對意圖的N250和P300進行溯源分析分別定位于額中回(BA10)和腦島(BA45)。結果表明大腦在多個階段對雙人肢體運動表達的互動意圖進行分類理解, 對負性敵對意圖表現出較早的理解且晚期持續性的評價加工。
心理理論; 友好意圖; 敵對意圖; 雙人互動; 事件相關電位
1 引言
社會互動意圖理解能力是個體成功與他人進行互動交流的關鍵技能之一(Cacioppo, Berntson, & Decety, 2010; 王益文, 黃亮, 張振, 宋娟, 白麗英, 2014; Wang, Huang, Zhang, Zhang, & Cacioppo, 2015)。這種技能在區分他人是朋友還是敵人時顯得尤為重要。大量實證研究(Carter & Pelphrey, 2008; van Wount& sanfey, 2008; Vrticka, Andersson, Sander & Vuilleumier, 2009)表明非言語行為, 包括面部表情、肢體語言, 提供了個體對他人互動意圖進行歸因的重要信息, 有助于判斷對方的行為意圖是友好還是敵對的。依據心理哲學和認知語用學, 互動(交際)意圖指一個人(A)的行為目標在當下至少指向另外一個人(B), 并被B所識別接收(Walter et al., 2004)。其中, 雙人依賴肢體運動來表達互動意圖不僅是生活中一種常見且重要的社交情境, 而且是個體在畢生發展(尤其是生命早期)與他人進行社會交往、相互學習和維持人際關系的重要手段, 探察大腦如何理解雙人肢體表達的不同性質的互動意圖具有現實意義。已有部分學者(Grafton, 2009; Ortigue, Sinigaglia, Rizzolatti, & Grafton, 2010)提出具身模仿理論, 認為當個體自己過去的動作經歷與當前觀察到的他人的動作一致時能夠促進個體對他人互動意圖的理解。當被觀察者越為個體所熟悉時, 越容易促進互動意圖的理解(Cacioppo, Juan, & Monteleone, 2017)。認知神經影像學研究已表明, 額?頂動作觀察網絡和社會腦網絡(包含內側前額葉皮層(medial prefrontal cortex, mPFC), 顳頂聯合皮層(temporal parietal junction, TPJ)和后部顳上溝等區域)在理解雙人肢體運動表達的互動意圖中扮演著重要角色(Centelles, Assaiante, Nazarian, Anton, & Schmitz, 2011)。盡管大多數神經成像研究(Ortigue et al, 2010; Young & Saxe, 2009)在探察理解中性意圖的神經機制方面取得了一定進展, 越來越多的研究開始逐漸由此轉向理解帶有情緒內容的動作意圖方面邁進。在過去的5年里, 絕大多數的神經成像研究(Decety & Cacioppo, 2012; Sinke, Sorger, Goebel, & de Gelder, 2010)集中揭示了消極性(疼痛或傷害)意圖領域的腦動態加工網絡, 包括TPJ、背外側mPFC、杏仁核和扣帶皮層等負責動作觀察、疼痛、心理理論和道德判斷的腦區。近來對積極性意圖的認知神經機制的研究也有了新的進展。Yoder和Decety (2014)研究發現在兩人互動的社會情境下, 與傷害性動作相比, 互助性動作顯著激活了右側背外側PFC和右側TPJ等一般被認為是負責自我?他人評價和道德決策判斷的腦區。甘甜、石睿、劉超和羅躍嘉(2018)采用經顱直流電刺激技術證實右側TPJ在助人意圖加工中的核心作用。以上神經成像研究對揭示大腦理解不同性質的互動意圖的空間特征較有裨益。然而, 由于功能性核磁共振技術(functional magnetic resonance image, fMRI)的時間分辨率較低, 對不同性質的互動意圖(比如, 友好意圖和敵對意圖)在這些大腦神經網絡中的動態時空加工特征尚不清楚。
正確理解不同性質的社會互動意圖是個體生存與發展必備技能之一。只有準確而快速地理解他人的互動意圖, 我們才能正確地選擇應對策略和反應方式, 做出與對方展開合作還是應對沖突的決策(王益文等, 2014; Wang et al., 2015)。認知推理他人不同性質的互動意圖有助于評價他人的行為是好還是壞, 是有害還是無害, 是友好還是敵對, 這是人類社會行為的一個重要成分(Blakemore & Decety, 2001; Malle & Holbrook, 2012)。發展心理學研究發現2歲甚至更早的幼兒, 與敵對個體相比, 更多表現出對友好個體的偏好(Buon et al., 2014), 且能依據互動雙方共同或相反的評價來理解他們之間是朋友還是敵人(Liberman, Kinzler & Woodward, 2014)。據研究者所知, 目前采用高時間分辨率的事件相關電位(Event-relate potential, ERP)技術探察社會互動情境下不同性質的互動意圖的神經動態加工時程的研究尚不多。Wang等人(2012)采用卡通漫畫作為實驗材料, 研究結果發現了個體理解私人意圖誘發的N250成分要顯著負于交際意圖, 而交際意圖在400~600 ms時間窗口誘發的晚期正成分(late positive component, LPC)則顯著正于私人意圖, 并據此提出理解私人意圖(相當于不存在互動的生物運動)要先于理解交際(互動)意圖的兩階段加工模型。Wang, Zheng, Lin, Wu和Shen (2011)及王益文、黃亮等人(2012)還發現, 理解單人心理是理解互動心理的基礎, 兩者屬于心理理論不同層次的加工水平, 并據此提出心理理論的多層次模型。然而上述研究并沒有對不同性質的互動(交際)意圖進行細致區分, 上述理論模型的內涵有待進一步豐富并獲得更多一致性證據的支持。Decety和Cacioppo (2012)要求被試觀看兩種簡短的包含道德內涵的動作場景:故意傷害行為和無意傷害行為。研究結果發現在刺激呈現后大腦右側后部顳上溝(60 ms)、杏仁核/顳極(122 ms)和腹內側PFC (182 ms)在觀看故意傷害行為的場景時先后被激活。該研究揭示了大腦在道德認知較早階段的快速信息加工時空特征(Decety & Cacioppo, 2012)。Proverbio等人(2011)采用高生態效度的圖片材料, 發現在合作性互動場景誘發的N2 (160~280 ms)波幅比情感性互動場景更負。王益文等人(2014)及Wang等人(2015)在以往研究的基礎上, 采用ERP技術和生態效度更高的真人照片作為刺激材料, 發現了在刺激呈現后較早階段(170~270 ms)友好意圖誘發的N2成分要顯著負于敵對意圖, 而晚期階段(270~500 ms)敵對意圖誘發的P3成分要顯著正于友好意圖, 在700~800 ms進一步發現互動意圖(友好和敵對意圖)和無互動意圖的LPC波幅存在顯著差異。LPC通常是指刺激呈現約300 ms 之后出現的一個正ERP成分, 可能反映大腦對心理理論不同層次水平的加工(Wang et al., 2012)。Liu, Sabbagh, Gehring和Wellman (2004)發現在大腦的左側前額區, 理解他人心理狀態的LPC成分(約在刺激呈現后800 ms)與理解物理世界有顯著差異。王益文、鄭玉瑋、沈德立、崔磊和閆國利(2012)采用四字成語做為材料考察解讀互動心理和單個人心理的差異, 發現在700~800 ms時, 在額中區的LPC只與解讀互動心理有關。盡管上述研究進一步豐富了社會意圖領域的研究, 并對大腦理解友好意圖和敵對意圖(定義可詳見黃亮、鄧兆鑫、任翰林、林國耀、王益文(2018)的研究)的時空動態加工機制進行了直接的探察, 但以往研究多采用頭部掩蔽或黑白照片, 實驗材料的生態效度有待進一步提高。此外, 王益文等人(2014)的研究中對中性無互動意圖的設置可能導致:(1)被試可能通過簡單的物理視覺線索(人物背對彼此)就能夠將無互動和互動意圖區分開來, 不涉及到心理意圖加工; (2)在日常生活中兩人由初始時相向而立到接下來背對彼此站立, 容易被觀察者知覺為一種不友好的負性社交場景(如, 兩人不喜歡彼此或發生了沖突分歧)。其行為研究結果發現在正確率和反應時上均存在意圖條件的主效應, 也進一步表明三種條件可能在任務難度上存在顯著差異, 無法完全排除ERP成分受任務難度的影響。另外, 王益文等人(2014)認為發生在腦內較早階段的N2成分僅與友好意圖的理解有關聯, 而行為結果卻顯示理解友好意圖的反應時顯著長于其它條件, 兩者之間似乎并不能完全相互支持。盡管他們給予了較恰當的解釋, 認為是兩種指標反映的心理加工階段不同導致, 但上述問題有待于進一步闡釋說明。
為考察大腦理解友好、敵對互動和中性無互動三種意圖的動態時空加工時程, 研究記錄了被試觀看兩個人物肢體動作表達的三類意圖的彩色照片并完成意圖推理任務時的腦電變化, 并進行腦溯源分析其空間特征。每個試次中包括兩張照片, 先后分別是動作準備及執行照片。值得說明的是, 本研究中的中性無互動意圖是嚴格參照Walter等人(2004)、Wang等人(2012)及王益文、黃亮等人(2012)的操作定義進行實踐, 包含兩個個體不相關聯的私人意圖內涵。基于以往的研究(Proverbio et al., 2011; Wang et al., 2015; 王益文, 黃亮等, 2012; 王益文等, 2014), 研究者假設在動作執行照片呈現后的較早加工階段, 中性意圖誘發的N250成分會顯著負于友好意圖, 且友好意圖誘發的N250成分也會顯著負于敵對意圖, 出現中性、友好和敵對意圖加工的初步分離; 而在加工晚期, 理解敵對意圖時誘發的P300成分可能顯著正于友好意圖和中性意圖, 理解敵對和友好互動意圖的LPC成分也會顯著正于中性意圖, 大腦理解不同性質的社會互動意圖的動態時空特征不同。
2 方法
2.1 被試
20名在校大學生(10女10男, 年齡范圍為19~25歲, 平均年齡為22.5歲), 右利手, 視力或者矯正視力正常, 沒有精神病史, 在實驗前均簽署了知情同意書, 實驗后給予一定的報酬。
2.2 實驗材料
實驗范式參考王益文等人(2014)及Wang等人(2015)的研究范式, 本研究采用兩張照片(動作準備及執行照片)先后呈現的方式, 使照片中人物的動作產生運動變化的景象, 要求被試在第二張動作執行照片呈現時先理解清楚后再盡快判斷照片中兩個人物的活動場景(Decety & Cacioppo, 2012; Ortigue et al., 2010)。為保證實驗材料的生態效度, 本研究的實驗材料是采用數碼相機拍攝, 演員為兩名男性在校研究生(年齡均為27歲, 身高分別為178 cm和176 cm)和兩名女性在校研究生(年齡分均為24歲, 身高分別為165 cm和158 cm), 同性別的演員處于同一場景, 照片中左邊的人物始終作為動作的發起者。實驗條件分為三種:友好意圖、敵對意圖、中性無互動意圖。第一種條件描述的是兩名人物友好合作的場景, 例如一個人拿著礦泉水瓶然后遞給另外一個人, 另外一個人伸手去接(見圖1a); 第二種條件描述的是兩名人物敵對沖突的場景, 例如一個人拿著礦泉水瓶然后砸向另外一個人, 另外一個人抬手去擋(見圖1b); 第三種條件作為控制條件, 描述的是兩名人物之間不存在互動、各自獨立活動的場景, 例如一個人拿著礦泉水瓶, 然后該人將礦泉水瓶放在自己嘴邊, 另外一個人則自己看著手機(見圖1c)。每種實驗條件下拍攝了70組照片, 每組照片由2張構成, 因而每種條件下共有140張照片。由于三種條件下相對應的每組照片中的第一張動作準備照片是相同的, 僅在于第二張動作執行照片不同, 因此三種條件下總共280張照片(相關示例請見圖1)。為了防止面部情緒和眼睛注視等線索對被試理解人物動作意圖的干擾, 所有照片經Photoshop對人物面部進行了馬賽克處理, 照片大小(4 cm × 3 cm)、像素(472×354)統一, 顏色為彩色照片, 照明度、對比度和飽和度一致。
首先隨機選取10名在校大學生(男女人數各半, 年齡在19~26歲之間), 要求他們判斷三種條件下每組照片(含動作準備和執行照片)中兩個人物的活動場景:如果認為左邊人物對右邊人物是友好的, 請按J鍵; 如果認為左邊人物對右邊人物是無意圖的(兩人之間不存在互動), 請按K鍵; 如果認為左邊人物對右邊人物是敵意的, 請按L鍵。每一組照片判斷結束后, 緊接著要求他們如實根據自身的感受對做出該判斷的難易程度進行5點評定(“1”代表非常容易, “2”代表有些容易, “3”代表一般, “4”代表比較困難, “5”代表非常困難)。剔除反應時超過2000 ms和正確率低于60%的照片, 最后篩選出符合要求的每種條件下的照片各40組, 選擇其中10組作為練習實驗材料, 剩余的30組照片(男性照片和女性照片各15組)作為正式實驗材料。對三種條件下的任務難度等級評定進行單因素方差分析, 結果發現不存在條件的顯著差異,(2, 117) = 2.03,= 0.136。該結果說明三種不同的意圖條件不存在任務難度的顯著差異。然后另外隨機選取10名在校大學生(男女生人數各半, 年齡在18~22歲之間), 要求他們對三種條件下的每組照片中兩個人物活動的友好或沖突程度做5點評定(“1”代表非常友好, “2”代表有些友好, “3”代表說不定, “4”代表有些沖突, “5”代表非常沖突)。經單因素方差分析發現, 三種不同條件下的照片友好或沖突的程度差異極其顯著,(2, 117) = 1111.00,< 0.001。進一步事后分析發現, 友好條件下照片的得分(1.48 ± 0.24)要顯著小于中性條件下照片的得分(3.00 ± 0.16),< 0.001, 以及顯著小于敵對條件下照片的得分(4.21 ± 0.35),<0.001, 并且中性條件下照片的得分顯著小于敵對條件下照片的得分,< 0.001。以上分析表明三種條件下的照片描述的友好或沖突程度有很好的區分度。所有參與實驗評定的被試均未參加正式實驗。
特別需要說明, 在日常社會交往中, 人們不僅通過觀察對方的身體語言來獲得其運動意圖信息, 而且常常將對方的面部表情和身體語言結合起來判斷對方的意圖。許多研究(van Wount & Sanfey, 2008; Carter & Pelphrey, 2008; Vrticka et al., 2009; Güro?lu et al., 2008)發現, 個體的不同效價的面部表情會影響到被試對個體意圖的判斷并影響到被試隨后的決策行為。本研究中, 我們對表演者的面部做了馬賽克處理, 減少了不同效價的面部表情以及眼睛注視等線索對身體語言理解的影響, 從而避免了其他因素對實驗結果的污染。另外, 本研究采用彩色人物照片和添加女性動作演員, 在實驗材料的生態效度和對無關變量的控制等方面都改進了以往的研究(王益文, 黃亮等, 2012; 王益文等, 2014)。除此之外, 所有照片的物理屬性匹配很好, 不同實驗條件下照片中的人物和物體、以及人物身體朝向基本是完全一致的, 排除了被試的腦電數據差異是由于不同物理屬性和任務難度差異所造成的。
2.3 實驗設計
被試內單因素三水平(意圖水平:友好意圖、敵對意圖和中性意圖)的腦認知實驗設計。行為學指標為正確率和反應時, 電生理學指標詳見2.6部分。
2.4 實驗程序
實驗流程見圖1。在每個試次中, 被試首先會看到提醒其實驗已開始的“+”注視點500 ms, 然后是一張持續1000 ms的動作準備照片, 在400~600 ms的隨機間隔之后, 呈現1500 ms的動作執行照片。在400~600 ms隨機間隔后進入下一個試次。實驗總試次為270次, 三種意圖條件下各90次, 呈現順序是偽隨機。整個實驗分兩段進行, 中間讓被試充分休息。實驗任務為要求被試在呈現動作執行照片時, 思考前后兩張照片的內在聯系, 先理解準確后再盡快判斷。判斷方式為按鍵反應, 為避免左、右手混淆, 均用右手反應。如果你認為照片中的左邊人對右邊人是友好的, 請按1鍵; 如果你認為照片中左邊人對右邊人沒有意圖, 請按2鍵; 如果你認為照片中左邊人對右邊人是敵意的, 請按3鍵。被 試需要在1500 ms內做出反應, 按鍵方式在被試間平衡, 超過1500 ms視為未反應。正式實驗前, 被試先進行一個簡短的練習實驗以熟悉實驗程序, 練習實驗與正式實驗的材料不同。
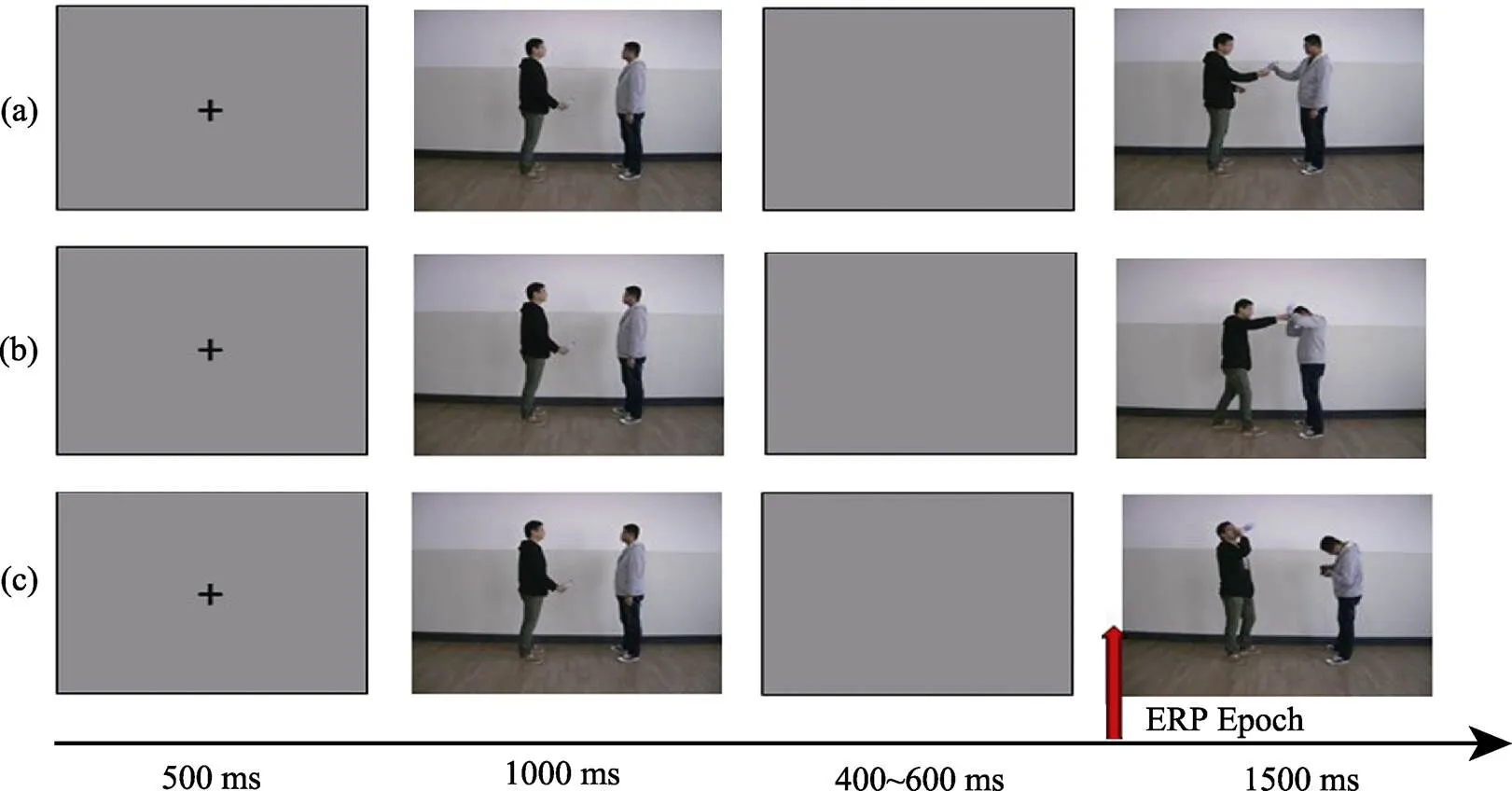
圖1 實驗流程圖示
2.5 腦電記錄
采用國際10~20系統擴展的64導電極帽, 以NeuroScan系統記錄EEG信號。腦電記錄時所有電極參考置于左乳突的一只參考電極, 離線分析時再次以置于右乳突的有效電極進行再參考, 即從各導聯信號中減去1/2該參考電極所記錄的信號, 轉化為以雙側乳突的平均值為參考。同時記錄雙眼外側的水平眼電(HEOG)和左眼上下眶的垂直眼電(VEOG)。濾波帶通為0.05~100 Hz, AC采樣, 采樣頻率為1000 Hz/導, 所有電極與頭皮之間阻抗都小于5 k?。對數據進行離線分析, 矯正眼電偽跡, 自動排除其他波幅大于± 75 μV的偽跡信號。
2.6 ERP數據處理與統計分析
在本研究中, 僅對動作執行照片呈現時的腦電數據進行分析和疊加, 并且根據照片呈現時的行為反應數據, 對正確反應的EEG進行疊加平均。分析時程為動作執行照片呈現后800 ms, 該刺激呈現前200 ms作為基線。本研究主要比較三類不同動作執行照片所誘發的ERPs成分的波幅的差異。采用SPSS 16.0軟件包錄入數據并進行統計分析。據已有研究(Wang, Ling, Yuan, Huang, & Shen, 2010; Wang et al., 2012; 王益文等, 2014)和本研究目的, 選擇9個電極點F3/FZ/F4、C3/CZ/C4、P3/PZ/P4, 進行進一步的數據分析。圖2顯示了20名被試在三種條件下的ERP總平均波形。從圖上可以看出, 三種條件(友好意圖、敵對意圖、中性意圖)下均誘發明顯的N250、P300和LPC成分。N250和P300的測量指標是峰值, 其時間窗口分別為170~270 ms和270~450 ms。此外, 從450~750 ms以每100 ms為間隔測量LPC的平均波幅。對在上述9個電極點上測得的ERPs成分的波幅進行3(意圖水平:友好意圖、敵對意圖、中性意圖) × 3(電極左右分布:左半球、中線、右半球) × 3(電極前后分布:額區、中央區、頂區)三因素重復測量方差分析。所有主效應和交互作用的值均采用Greenhouse-Geisser法校正, 事后配對比較采用Bonferroni法校正。正式實驗中每個實驗條件的照片組重復3次, 即每個條件下90個試次。去偽跡后, 友好意圖、敵對意圖和中性意圖三種條件下ERP總平均的有效疊加次數分別為70, 71和72。用sLORETA軟件對友好和敵對意圖條件下的N250和P300進行溯源定位, 考察大腦理解友好和敵對互動意圖的空間特征。

圖2 友好、敵意、中性意圖條件下ERP總平均波形圖
3 結果
3.1 行為結果
友好、敵對和中性三種條件的平均正確率見表1。采用單因素重復測量方差分析發現, 不存在正確率的顯著差異,(2, 38) = 3.40,> 0.05。該分析說明三種條件在任務難度上是基本相當的, 與材料篩選中對實驗任務難度的評定是相一致的。
友好、敵對和中性三種條件的平均反應時見表1。采用單因素重復測量方差分析發現, 存在反應時的顯著差異,(2, 38) = 42.74,< 0.001,= 1741.66, η= 0.69。事后分析發現, 友好和中性條件的反應時均顯著長于敵對條件,s < 0.001, 而友好與中性條件的反應時差異不顯著,> 0.05。

表1 三種意圖動作分類任務的正確率和反應時(M± SD)
3.2 ERP結果
3.2.1 N250
對N250的峰值進行3(意圖水平)×3(電極左右分布)×3(電極前后分布)的三因素重復測量方差分析(見表2), 發現意圖水平的主效應差異極其顯著,(2, 38) = 15.97,< 0.001,= 6.59, η= 0.46。事后比較分析發現中性條件的波幅(?2.72 ± 0.79 μV)比友好(?1.69 ± 0.68 μV)和敵對條件(?1.24 ± 0.82 μV)均更負,s < 0.05, 而友好與敵對條件的波幅無顯著差異,0.05 (見圖3a左側)。
統計分析結果還發現意圖水平與電極前后分布的交互作用顯著,(4, 76) = 4.07,< 0.05,= 3.16, η= 0.18。簡單效應分析發現條件在全腦上均存在顯著差異,s < 0.05。事后進一步分析發現, 在大腦額區, 中性條件的波幅比友好和敵對條件均更負,s < 0.05, 且友好條件的波幅也比敵對條件也更負,< 0.05。在大腦中央區, 中性條件的波幅比友好條件更負,< 0.05, 中性條件的波幅比敵對條件也更負,< 0.001, 友好與敵對條件的波幅則無顯著差異,0.05。在大腦頂區, 僅發現中性條件的波幅比友好條件更負,< 0.01, 未發現其它顯著差異。統計分析結果還發現意圖水平與電極左右分布的交互作用,(4, 76) = 3.37,< 0.05,= 0.91, η= 0.15。簡單效應分析發現條件在全腦上均存在顯著差異,s < 0.001。事后進一步分析發現, 在大腦全腦上均發現中性條件的波幅比友好和敵對條件均更負,s < 0.05, 而友好和敵對條件的波幅則無顯著差異,s0.05。

表2 N250和P300的波幅方差分析結果
注: 事后比較中1代表友好意圖, 2代表敵對意圖, 3代表中性意圖。***< 0.001, **< 0.01, *< 0.05
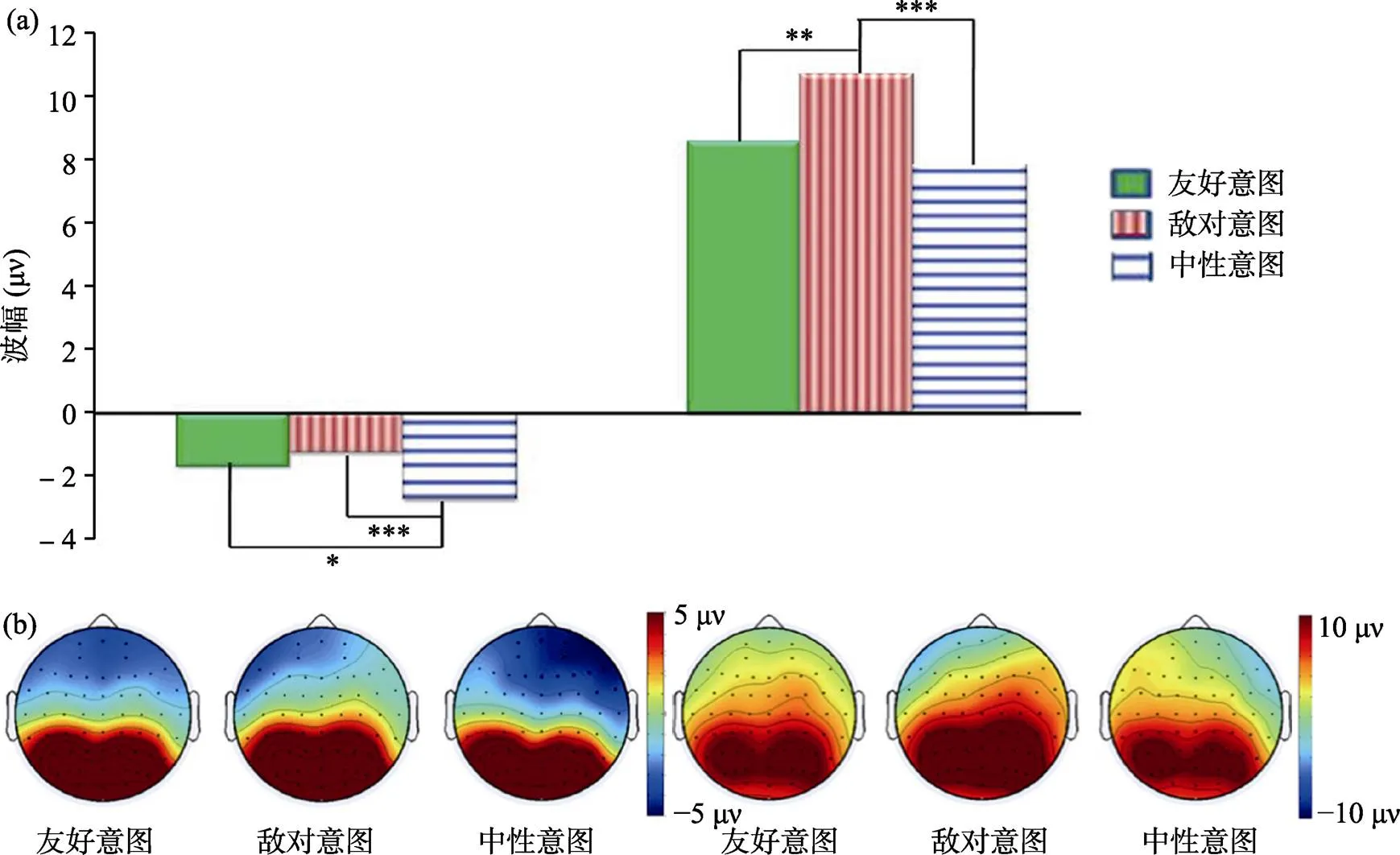
圖3 三種條件下N250波幅(a左側)和地形圖(b左側)以及P300的波幅(a右側)和地形圖(b右側)
值得注意的是, 統計分析結果顯示意圖水平與電極前后、左右分布的三重交互作用極其顯著,(8, 152) = 8.61,< 0.001,= 1.01, η= 0.31。該結果表明友好、敵對和中性三種條件在某些電極分布上存在顯著差異。簡單效應分析發現三種條件在大腦的F3、FZ、F4、C3、Cz、C4、P3和PZ點均發現存在顯著差異, 僅在P4點未發現有顯著差異。簡單簡單效應分析發現, 在左側額區(F3)和左側中央區(C3), 僅發現中性條件的波幅比敵對條件更負,s < 0.05, 未發現其它顯著差異。在額中區(FZ), 中性條件的波幅比友好和敵對條件均更負,s < 0.05, 且友好條件的波幅比敵對條件也更負,= 0.042。在F4、CZ、C4、P3和PZ, 均發現中性條件的波幅比友好和敵對條件都更負,s < 0.05, 而友好與敵對條件的波幅則無顯著差異,s0.05。
綜上可知, 在大腦額區(尤其在FZ點)發現了3種意圖條件的兩兩分離, 中性條件的N250波幅比友好和敵對條件均更負, 且友好條件的N250波幅比敵對條件也顯著更負。
3.2.2 P300
對P300的峰值進行了三因素重復測量方差分析(見表2), 發現意圖水平的主效應差異極其顯著,(2, 38) = 19.85,< 0.001,= 27.84, η= 0.51。事后分析發現, 敵對條件的波幅(10.73 ± 0.06 μV)比友好(8.62 ± 0.74 μV)和中性條件(7.86 ± 0.77 μV)均更正,s < 0.01, 友好與中性條件的波幅也達到邊緣顯著差異,= 0.069 (見圖3a右側)。
統計分析結果還發現意圖水平與電極前后分布的交互作用顯著,(4, 76) = 12.18,< 0.001,= 6.51, η= 0.39。簡單效應分析發現, 條件在全腦上均有顯著差異,(2, 38) = 6.90/14.90/27.84,s < 0.01。事后進一步分析發現, 在大腦額區, 友好和敵對條件的波幅均比中性條件更正,s < 0.05, 友好與敵對條件則無顯著差異,0.05。在大腦中央區和頂區, 均發現敵對條件的波幅比友好和中性條件都更正,s < 0.01, 而友好與中性條件則無顯著差異,s0.05。統計分析結果還發現意圖水平與電極左右分布的交互作用,(4, 76) = 15.23,< 0.001,= 2.64, η= 0.45。簡單效應分析發現, 條件在全腦上均存在顯著差異,s < 0.01。事后進一步分析發現, 在大腦左半球, 僅發現敵對條件的波幅比中性條件更正,< 0.05, 未發現其它顯著差異。在大腦中線, 敵對條件的波幅比友好和中性條件均更正,s < 0.01, 友好與中性條件則無顯著差異,= 0.531。在大腦右半球, 敵對條件的波幅比友好和中性條件均更正,s < 0.001, 且友好條件比中性條件也更正,< 0.01。該結果表明在大腦右半球, 從中性、友好到敵對三種條件的激活程度呈現出階梯遞增的現象。
值得注意的是統計分析結果顯示條件與電極前后、左右分布的三重交互作用極其顯著,(8, 152) = 6.87,< 0.001,= 1.26, η= 0.27。簡單效應分析發現三種條件在大腦FZ、F4、Cz、C4、P3、PZ和P4等點均發現存在顯著差異, 僅在F3和C3等點未發現有顯著差異。簡單簡單效應分析卻未發現在額中區(FZ)存在條件的顯著差異,0.05。在中央區(CZ)、左側頂區(P3)、中部頂區(PZ)和右側頂區(P4), 敵對條件的波幅比友好和中性條件均更正,s < 0.01, 友好與中性條件的波幅則無顯著差異,s0.05。在右側額區(F4)和右側中央區(C4), 敵對條件的波幅比友好和中性條件均更正,s < 0.05, 友好條件的波幅比中性條件也更正,s < 0.05。該趨勢與上面提到的右半球優勢效應基本一致。
綜上可知, 在大腦右半球(尤其在F4和C4點)發現了3種意圖條件的兩兩分離, 敵對條件的P300波幅比友好和中性條件顯著更正, 且友好條件的P300波幅比中性條件也顯著更正。
3.2.3 LPC
對450~750 ms的平均波幅, 以每100 ms為間隔, 做三因素重復測量方差分析(見表3)。統計分析結果發現在450~550 ms時間窗口, 存在意圖水平的主效應差異,(2, 38) = 12.39,< 0.001,= 20.32, η= 0.40。事后分析發現, 友好條件(6.14 ±0.76 μV)和敵對條件(6.94 ± 0.95 μV)的波幅均比中性條件(4.65 ± 0.78 μV)更正,< 0.01, 而友好與敵對條件則無顯著差異,0.05。
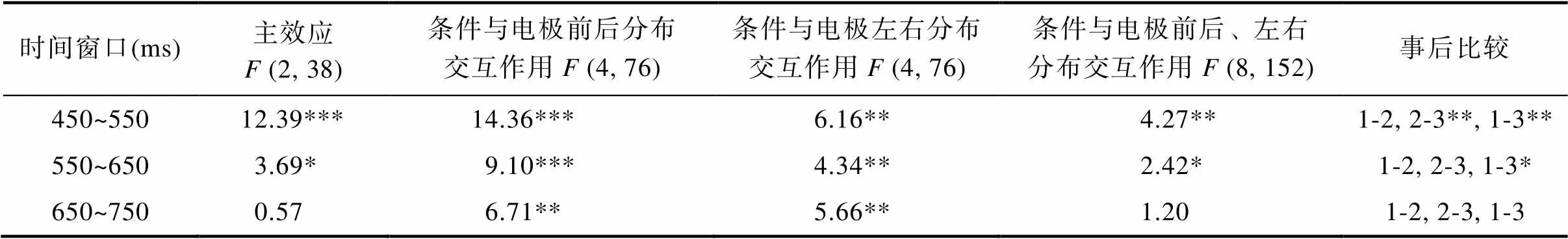
表3 450~750 ms LPC波幅方差分析結果
注: 事后比較中1代表友好意圖, 2代表敵對意圖, 3代表中性意圖。***< 0.001, **< 0.01, *< 0.05
統計分析結果還發現存在意圖水平與電極前后分布的交互作用,(4, 76) = 14.36,< 0.001,= 4.74, η= 0.43。簡單效應分析發現在全腦區均存在條件的顯著差異,s < 0.001。事后進一步分析發現, 在大腦額區和中央區, 友好和敵對條件的波幅均比中性條件更正,s < 0.05, 而友好與敵對條件則無顯著差異,s0.05。在大腦頂區, 敵對條件的波幅比友好和中性條件均更正,s < 0.01, 而友好與中性條件則無顯著差異,0.05。統計分析結果還發現存在意圖水平與電極左右分布的交互作用,(4, 76) = 6.16,< 0.01,= 2.97, η= 0.25。簡單效應分析發現在大腦中線和右半球存在條件的顯著差異,s < 0.001。事后進一步分析發現, 在大腦的中線, 僅發現敵對條件的波幅比中性條件更正,< 0.01, 未發現其它顯著差異。在大腦的右半球, 敵對條件的波幅比友好和中性條件均更正,s < 0.05, 友好比中性條件的波幅也更正,< 0.001。
統計分析結果還發現存在意圖水平與電極左右、前后分布的三重交互作用,(8, 152) = 4.27,< 0.01,= 1.25, η= 0.184。簡單效應分析發現三種條件在大腦的F3、FZ、F4、Cz、C4、P3、PZ和P4均發現存在顯著差異, 僅在C3未發現有顯著差異。簡單簡單效應分析發現在左側額區(F3)和額中區(FZ), 僅發現友好條件的波幅比中性條件更正,s < 0.01, 未發現其它顯著差異。在中央區(CZ), 僅發現敵對條件的波幅比中性條件更正,< 0.01, 未發現其它顯著差異。在右側額區(F4)和左側頂區(P3), 友好和敵對條件的波幅均比中性條件更正,s < 0.001, 而友好與敵對條件的波幅則無顯著差異,s0.05。在右側中央區(C4), 友好和敵對條件的波幅均比中性條件更正,< 0.01, 敵對條件的波幅比友好條件也更正,< 0.01。在中部頂區(PZ)和右側頂區(P4), 敵對條件的波幅比友好和中性條件均更正,s < 0.01, 而友好與中性條件的波幅則無顯著差異,s0.05。
統計分析結果發現在550~650 ms時間窗口, 存在意圖水平的主效應差異,(2, 38) = 3.69,< 0.05,= 33.57, η= 0.16。事后分析僅發現友好條件的波幅(6.78 ± 0.75 μV)比中性條件(5.29 ± 0.82 μV)更正,< 0.05, 而友好條件的波幅與敵對條件(6.54 ± 0.90 μV)無顯著差異,0.05, 敵對條件與中性條件的波幅也無顯著差異,0.05。
統計分析結果還發現存在意圖水平與電極前后分布的交互作用,(4, 76) = 9.10,< 0.001,= 5.78, η= 0.32。簡單效應分析發現在大腦額區和中央區存在顯著差異,s < 0.01。事后進一步分析發現, 在大腦額區, 僅發現友好條件的波幅比中性條件更正,< 0.01, 未發現其它顯著差異。在大腦中央區, 友好和敵對條件的波幅均比中性條件更正,s < 0.05, 友好與敵對條件則無顯著差異,= 1.000。統計分析結果還發現存在意圖水平與電極左右分布的交互作用,(4, 76) = 4.34,< 0.01,= 2.95, η= 0.19。簡單效應分析發現僅在大腦右半球存在顯著差異,< 0.001。事后進一步分析發現, 在大腦右半球, 友好和敵對條件的波幅均比中性條件更正,s < 0.05, 友好與敵對條件則無顯著差異,0.05。
統計分析結果還發現存在意圖水平與電極左右、前后分布的三重交互作用,(8, 152) = 2.42,< 0.05,= 1.41, η= 0.11。簡單效應分析發現三種條件在大腦F3、FZ、F4、C4等點均發現存在顯著差異。簡單簡單效應分析發現, 在左側額區(F3)僅發現友好條件的波幅比中性條件更正,< 0.05, 未發現其它顯著差異。在右側額區(F4)和右側中央區(C4), 友好和敵對條件的波幅均比中性條件更正,s < 0.01, 友好與敵對條件的波幅則無顯著差異,s0.05。在額中區(FZ)未發現條件存在任何顯著差異,0.05。
統計分析結果發現在650~750 ms時間窗口, 意圖水平的主效應不顯著(見表3),(2, 38) = 0.57,0.05,= 31.66, η= 0.03。盡管統計分析結果發現存在意圖水平與電極前后分布、意圖水平和電極左右分布的兩個交互作用,(4, 76) = 6.71/5.66,s < 0.01,= 5.07/3.30, η= 0.26/0.23, 但進一步分析卻未發現任何有統計學意義的顯著差異。
綜上可知, 在450~750 ms的時間窗口, 在大腦右半球(C4點等)發現了3種意圖條件的LPC成分呈現出由存在兩兩差異到互動與中性條件的差異再到不同條件間的差異完全消失這種發展變化趨勢。
3.3 腦地形圖
值得關注的是, 從170~270 ms三種條件的N250波幅存在的顯著差異, 這種差異在在三種條件的腦地形圖中的到印證, 如圖3(b左)所示。該圖是根據三種條件在270~500 ms時間窗口內的峰值做出, 顯示出與統計分析發現的條件與電極左右分布或條件與電極前后分布的顯著交互作用一致, 在全腦分布上, 中性條件比友好和敵對條件的激活程度都更小, 而友好和敵對條件的激活程度則沒有顯著差異。從270~500 ms三種條件的P300波幅存在顯著差異。這種差異在三種條件的腦地形圖中得到印證, 如圖3(b右)所示。該圖是根據三種條件在270~500 ms時間窗口內的峰值做出, 顯示出與統計分析發現的條件與電極左右分布或條件與電極前后分布的顯著交互作用一致, 在大腦頂區, 敵對條件比友好和中性條件被更顯著激活, 而友好和中性條件的激活區域則沒有顯著差異; 在大腦的右半球, 從中性、友好到敵對三種條件呈現一種階梯性激活增強的趨勢。
3.4 sLORETA結果
先用 sLORETA 軟件分別對中性條件和友好條件、以及中性條件和敵對條件的N250原始波進行溯源分析, 發現抑制的腦區分別位于Brodmann 6區(BA6, 見圖4a)和Brodmann 2區(BA2, 見圖4b), 進行比較后均未發現活動存在顯著差異的腦區。然后, 對友好條件和敵對條件兩種條件下的N250原始波進行溯源分析, 激活的腦區都主要位于Brodmann 10區(BA10), 額葉區的額中回(圖4c), 對兩種條件下激活的腦區進行比較后未發現存在顯著差異的腦區。對P300原始波進行了溯源分析, 發現友好條件和敵對條件下, 激活的腦區都主要位于BA45區, 腦島(圖4d), 對兩種條件下激活的腦區進行比較后發現, 在額葉、下葉(sublobar)和邊緣葉等區域的39個體素上發現顯著的負性電流差異(最大值為0.96,< 0.01), 其中64%的個體素位于額葉的額中回, 額下回, 額內側回等區域, 18%的個體素位于下葉的腦島等區域, 另外18%的個體素位于前扣帶回等區域。

圖4 對中性意圖與友好、敵對意圖條件下的N250進行溯源分析的最大抑制腦區分別為BA6區(a)和BA2區(b); 對友好意圖和敵對意圖兩種條件下N250和P300進行溯源的最大激活腦區分別位于BA10區(c)和BA45區(d)
4 討論
4.1 N250反映大腦對互動意圖中動作目標不同社會屬性的初步分類
本研究采用ERP技術考察大腦理解雙人互動背景下基于肢體表達的友好和敵對互動意圖的動態時空特征。本研究對正確率的分析結果顯示三種意圖條件無顯著差異, 反應時分析結果顯示友好意圖和中性意圖的反應時均顯著長于敵對意圖, 友好意圖和中性意圖的反應時沒有顯著差異, 而王益文等人(2014)的研究則發現友好意圖的正確率顯著低于無互動意圖, 友好意圖的反應時顯著長于敵對意圖和無互動意圖, 敵對意圖與無互動意圖的反應時無顯著差異。本研究結果與王益文等人的研究結果不完全一致。究其原因在于兩者在中性無互動意圖內涵的建構上不同, 本研究中的中性無互動意圖是嚴格參照Walter等人(2004)、Wang等人(2012)及王益文、黃亮等人(2012)的操作定義進行實踐, 克服了王益文等人(2014)研究中被試僅憑實驗材料中人物身體朝向(背對彼此)這一外在物理特征來理解中性意圖的不足, 且排除了ERP成分受到任務難度等因素影響的潛在可能性。
ERP結果顯示分布在大腦額區的友好意圖的N250波幅比敵對意圖顯著更負, 且中性意圖的N250波幅比友好意圖也顯著更負。換言之, 從敵對意圖、友好意圖到中性意圖的N250波幅呈現逐步更負的趨勢。王益文、黃亮等人(2012)發現私人(中性無互動)意圖的N250波幅比交際(互動)意圖顯著更負。王益文等人(2014)進一步將互動意圖按動作目標的不同社會屬性區分為友好和敵對意圖, 并發現友好意圖的N2波幅比敵對意圖顯著更負。綜合上述兩項系列研究發現, 從敵對意圖到友好意圖再到中性意圖可能呈現一種階梯性更負趨勢, 本研究結果與前人研究結果是基本一致的。Proverbio等人(2011)發現高生態背景下合作性互動動作誘發的N2 (160~280 ms)波幅比情感性互動動作顯著更負。Decety和Cacioppo (2012)采用高密度EEG研究技術發現故意傷害性動作知覺在182 ms左右激活了腹內側PFC, 反映了道德認知早期的快速信息加工階段。本研究中敵對意圖動作和友好意圖動作均誘發了額區的N250成分, 且出現了兩者之間的分離, 可能表明N250反映了大腦對互動意圖中不同動作目標社會屬性的分類理解。至于王益文等人(2014)則發現友好意圖的N2波幅顯著大于敵對意圖以及與無互動意圖的差異也達到邊緣顯著, 敵對意圖與無互動意圖的N2波幅則沒有顯著差異, 本研究結果與前人研究結果不一致。王益文等人對無互動意圖的設置可能導致被試能通過簡單的物理視覺線索來理解該實驗條件, 而完全不涉及到心理意圖加工; 另外, 在日常生活中兩人背對彼此站立容易被觀察者知覺為一種不友好的負性社交場景, 因而在王益文等人的研究中理解無互動意圖與敵對意圖需要耗費的認知神經資源均較少且無顯著差異。根據該思路, 研究者認為N250成分可能反映了對互動意圖動作目標的不同社會屬性的分類加工, 能夠較早的區分敵對、友好和中性意圖, 且反應時結果也顯示對敵對意圖表現出較早的理解, 而不是Wang等人(2014)提出的N2僅與友好意圖的理解有關。此外, 已有研究發現, 相比動作量少的圖片, 包含較多動作的身體刺激會誘發較大的N170波幅(Borhani, Borgomaneri, Làdavas, & Bertini, 2016)。在本研究中, 中性意圖包含兩個不相關聯的人物身體動作, 與友好和敵對意圖相比, 包含的動作內容更多, 且友好意圖在動作內容上或模糊程度上也比敵對意圖更甚。因此N250在三種不同條件上的差異也可能反映了動作內容的復雜性或互動情境的模糊性。至于在時間進程上與前人研究不完全一致, 可能是因為實驗材料與任務不相同。
認知神經科學研究已經表明, 推測他人的意圖需要依賴額?頂動作觀察網絡和一個假定的社會腦網絡(Centelles et al., 2011)。多數研究者認為以上兩個腦網絡系統在動作理解中扮演著不同卻互補的角色(Isoda, 2016), 然而還有一些研究者認為兩者扮演的是完全獨立的角色(Catmur, 2015)。前者對偶發性社會動作進行一般性的加工, 而后者對由于異常的運動特性所導致的意圖理解進行加工(Georgescu et al., 2014)。本研究中, 在大腦前額區域發現友好和敵對意圖的皮層電流密度差異。確切的說, 在大腦的額中區(FZ點)發現友好和敵對意圖的N250波幅和皮層電流密度存在顯著差異。另外, 對友好和敵對意圖的N250成分進行溯源分析的結果發現, 兩種條件下的最大激活區定位于BA10區, 額葉區的額中回, 但激活程度不存在顯著差異。近來, 許多神經影像學研究(Maranesi, Livi, Fogassi, Rizzolatti, & Bonini, 2014; Yoder & Decety, 2014)發現背外側PFC在理解單人抓握動作或雙人互動動作時進行早期自上而下的預測加工中都被顯著激活。Amoruso, Finisguerra和Urgesi (2018)認為背外側PFC可能在對他人基于情境的動作進行預期加工時扮演一般普遍性的角色。另外一項fMRI研究發現額下回在理解手部動作傳遞的互動意圖時被顯著激活(M?tt?nen, Farmer, & Watkins, 2016)。額中回作為鄰近背外側mPFC和額下回的一個腦區, 本研究結果再次表明前額皮層在理解肢體表達的友好和敵對意圖動作時扮演重要角色。
4.2 P300反映大腦對互動意圖的不同社會情感意義的評價加工
在270~450 ms, ERP結果發現在大腦右半球, 敵對意圖的P300波幅比友好和中性意圖更正, 且友好意圖比中性意圖的P300波幅也更正。該結果與前人研究結果較一致。相關研究發現在加工威脅性的面孔或有威脅性的提示詞語時誘發的P300 / LPC波幅更正(Moser, Huppert, Duval, & Simons, 2008; Weymar, Bradley, Hamm, & Lang, 2013)。敵對意圖在本研究中含有威脅性的信息, 誘發了更強的P300波幅。王益文等人(2014)也發現, 在大腦的頂區、中線和右半球, 敵對意圖的P3 峰值比友好意圖和中性無互動意圖更正。上述研究均表明P300成分可能反映大腦對負性信息更強的注意和評價加工。然而, Wu等人(2014)發現, 與中性刺激相比, 負性刺激在女性的經期晚期(黃體期)比經期早期(卵泡期)誘發了顯著的N2 (150~300 ms)。本研究結果與其不太一致, 可能是由于刺激材料和實驗任務不同所致。Wieser, McTeague和Keil (2011)認為高情緒效價的刺激, 比如社會威脅線索, 會在大腦中得到優先并持續的加工。本研究中, 與友好意圖相比, 敵對意圖的行為反應時顯著的更短且誘發了更大的P300波幅。研究結果表明個體在理解具有負性社會情感意義的敵對意圖時投入了較多的認知神經資源, 可能包含了注意、感知和認知評價等復雜心理加工過程。個體對威脅性信息保持高度警覺性和深層次的加工有利于人類生存, 符合進化學的觀點。
許多fMRI腦成像研究(Sinke et al., 2010; Decety & Cacioppo, 2012)均發現被試的額?頂動作觀察網絡和負責加工道德違犯的大腦區域(TPJ、旁扣帶皮層、內側眶額皮層、杏仁核)在觀看威脅性電影片段或圖片時都被顯著激活, 且額?頂動作觀察網絡與這些區域的神經連接性也顯著增強。Watanabe等人(2012)發現前扣帶皮層/腹內側PFC和背內側PFC在健康被試基于非言語線索進行的友好或敵對判斷時均被強烈激活。本研究發現與前人的研究結果部分一致。許多社會認知神經科學研究表明P300成分主要分布在頂區, 該區域在加工個體動作時的重要性得到了一致性的證據。在腦地形圖中, 意圖條件主效應的差異在270~450 ms時間窗口內的也得到印證, 顯示出在大腦頂區, 敵對意圖比友好和中性意圖都被更顯著激活。對友好和敵對意圖的P300成分進行溯源分析的結果發現, 兩種條件下的最大激活區位于BA45區, 下葉的腦島, 且在額葉的額中回、額下回、額內側回等區域, 下葉的腦島等區域, 以及前扣帶回等區域發現敵對意圖的激活強度顯著正于友好意圖。許多關于情緒的腦成像研究(Gilead, Katzir, Eyal, & Liberman, 2016)表明腦島在加工基本情緒信息, 包括憤怒等負性情緒中扮演重要的角色。腦島是負責評價和情緒加工的腦神經網絡重要組成部分之一, 在整合情緒和認知加工中起著廣泛的作用(Berntson et al., 2011)。本研究結果也表明腦島等區域可能在對友好和敵對意圖的社會情感意義進行評價加工起著重要作用。意圖條件在270~450 ms時間窗口內的腦地形圖中激活的差異反映出在大腦右半球, 從中性意圖、友好意圖至敵對意圖呈現一種階梯性激活增強的趨勢, 與認知心理學中關于右半球在情緒加工中起重要作用的論斷也是一致的。在本研究中, 敵對意圖作為一種威脅性的信號, 可能不可避免地誘發了被試的某種負性情緒(如不愉悅)。友好和敵對意圖在激活腦區的顯著差異, 可能表明了大腦理解不同性質的互動意圖在300 ms左右時開始進一步出現空間上的分離, 對負性敵對意圖的社會情感意義表現出優勢評價加工。該結論一方面對前人(Bahnemann, Dziobek, Prehn, Wolf, & Heekeren, 2010)提出的意圖理解兩階段加工模型進行了補充, 另一方面為心理理論的分層認知框架(HMCF)的假設模型(Wang et al., 2011, 2012)提供了更豐富的神經電生理學證據。LPC成分在550~650 ms發生互動意圖與中性意圖的分離, 表明晚期階段的LPC成分可能與更復雜的社會認知加工相關, 可能反映了大腦對互動意圖后續的理解加工, 這與前人關于LPC (700~800 ms)的發現(Wang et al., 2011, 2012; 王益文等, 2014)是一致的。至于在時間進程上不完全重合, 可能是由于實驗材料的不同造成的。
由于自閉癥群體對互動情境下的雙人肢體運動知覺存在損傷(Kaiser, & Pelphrey, 2012), 本研究將有助于為探查其社會互動缺陷的機制提供一個較好的范式。當然本研究中的ERP波幅還可能會受到不同的動作運動學特性的影響, 因此在今后的研究中應該思考如何更好地加以控制, 對本研究結果的推論也應當更加謹慎。此外, 在部分關于動作意圖的研究(Adenzato et al., 2017; 黃亮等, 2018)中發現意圖理解的性別差異, 以及意圖影響肢體運動特征的新視角(Ansuini, Cavallo, Bertone, & Becchio, 2015), 值得我們在未來研究中深入探討。
5 結論
綜上所述, 本研究采用ERP技術, 主要從N250和P300 (時間進程)及其溯源分析(空間角度)考察了大腦理解雙人肢體運動表達的友好和敵對互動意圖的動態時空加工過程。結果表明:
在腦內早期階段, N250成分表明大腦在250 ms左右就對敵對、友好和中性意圖從動作目標上進行了分類加工, 三種意圖條件的N250成分出現了兩兩分離。溯源分析發現該成分主要源于額中回。從行為反應時可以發現大腦對負性敵對意圖表現出了最早的理解加工。
在腦內晚期階段, P300成分表明大腦在300 ms左右對敵對、友好和中性意圖的社會情感性進一步進行了評價加工, 三種意圖條件的P300成分也出現了兩兩分離。溯源分析發現該成分主要源于腦島。大腦右半球的加工優勢效應表現出了對負性敵對意圖的持續性評價加工。
Adenzato, M., Brambilla, M., Manenti, R., de Lucia, L., Trojano, L., Garofalo, S., .... Cotelli, M. (2017). Gender differences in cognitive theory of mind revealed by transcranial direct current stimulation on medial prefrontal cortex., 41219.
Amoruso, L., Finisguerra, A., Urgesi, C. (2018). Contextualizingaction observation in the predictive brain: Causal contributions of prefrontal and middle temporal areas.,, 68–78.
Ansuini, C., Cavallo, A., Bertone, C., & Becchio, C. (2015). Intentions in the brain: The unveiling of Mister Hyde.,(2), 126–135.
Bahnemann, M., Dziobek, I., Prehn, K., Wolf, I., & Heekeren, H. R. (2010). Sociotopy in the temporoparietal cortex: Common versus distinct processes.,(1), 48–58.
Berntson, G. G., Norman, G. J., Bechara, A., Bruss, J., Tranel, D., & Cacioppo, J. T. (2011). The insula and evaluative processes.,(1), 80–86.
Blakemore, S. J., & Decety, J. (2001). From the perception of action to the understanding of intention.,(8), 561–567.
Borhani, K., Borgomaneri, S., Làdavas, E., & Bertini, C. (2016). The effect of alexithymia on early visual processing of emotional body postures., 1–8.
Buon, M., Jacob, P., Margules, S., Brunet, I., Dutat, M., Cabrol, D., & Dupoux, E. (2014). Friend or foe? Early social evaluation of human interactions.(2), e88612.
Cacioppo, J. T., Berntson, G. G., & Decety, J. (2010). Social neuroscienceanditsrelationshiptosocialpsychology.(6), 675–685.
Cacioppo, S., Juan, E., & Monteleone, G. (2017). Predicting intentions of a familiar significant other beyond the mirror neuron system., 155.
Carter, E. J., & Pelphrey, K. A. (2008). Friend or foe? Brain systems involved in the perception of dynamic signals of menacing and friendly social approaches.,(2), 151–163.
Catmur, C. (2015). Understanding intentions from actions: Directperception, inference, and the roles of mirror and mentalizing systems., 426–433.
Centelles, L., Assaiante, C., Nazarian, B., Anton, J.-L., & Schmitz,C. (2011). Recruitment of both the mirror and the mentalizing networks when observing the social interactions depicted by point-lights: A neuroimaging study.,(1), e15749.
Decety, J. & Cacioppo, S. (2012). The speed of morality: A high-density electrical neuroimaging study.,(11), 3068–3072.
Gan, T., Shi, R., Liu, C. & Luo Y. J. (2018). Cathodal transcranial direct current stimulation on the right temporo-parietal junction modulates the helpful intention processing.(1), 36–46.
[甘甜, 石睿, 劉超, 羅躍嘉. (2018). 經顱直流電刺激右側顳頂聯合區對助人意圖加工的影響.(1), 36–46.]
Georgescu, A. L., Kuzmanovic, B., Santos, N. S., Tepest, R., Bente, G., Tittgemeyer, M., & Vogeley, K. (2014). Perceiving nonverbal behavior: Neural correlates of processing movement fluency and contingency in dyadic interactions.,(4), 1362–1378.
Gilead, M., Katzir, M., Eyal, T., & Liberman, N. (2016). Neural correlates of processing "self-conscious" vs. "basic" emotions.,, 207–218.
Grafton, S. T. (2009). Embodied cognition and the simulation of action to understand others., 97–117.
Güro?lu, B., Haselager, G. J. T., van Liershout, C. F. M., Takashima, A., Rijpkema, M., & Fernández, G. (2008) Why are friends special? Implementing a social interaction simulation task to probe the neural correlates of friendship.(2), 903–910.
Huang, L., Deng, Z. X., Ren, H. L., Lin, G. Y., & Wang Y. W. (2018). The role of stimulus ecologicalness and genders in understanding kind versus hostile intentions.(2), 435–440.
[黃亮, 鄧兆鑫, 任翰林, 林國耀, 王益文. (2018). 材料生態性和性別因素對理解友好與敵對意圖的影響.,(2), 435–440.]
Isoda, M. (2016). Understanding intentional actions from observers' viewpoints: A social neuroscience perspective., 1–9.
Kaiser, M. D., Pelphrey, K. A. (2012). Disrupted action perception in autism: Behavioral evidence, neuroendophenotypes, and diagnostic utility.,(1)25–35.
Liberman, Z., Kinzler, K. D., & Woodward, A. L. (2014). Friends or foes: Infants use shared evaluations to infer others' social relationships.(3), 966–971.
Liu, D., Sabbagh, M. A., Gehring, W. J., & Wellman, H. M. (2004). Decoupling beliefs from reality in the brain: An ERP study of theory of mind.,(6), 991–995.
Malle, B. F., & Holbrook, J. (2012). Is there a hierarchy of social inferences? The likelihood and speed of inferring intentionality, mind, and personality.,(4), 661–684.
Maranesi, M., Livi, A., Fogassi, L., Rizzolatti, G., & Bonini, L. (2014) Mirror neuron activation prior to action observation in a predictable context.,(45), 14827–14832.
Moser, J. S., Huppert, J. D., Duval, E., & Simons, R. F. (2008). Face processing biases in social anxiety: An electrophysiological study.,(1), 93–103.
M?tt?nen, R., Farmer, H., & Watkins, K. E. (2016). Neural basis of understanding communicative actions: Changes associated with knowing the actor’s intention and the meanings of the actions.,, 230–237.
Ortigue, S., Sinigaglia, C., Rizzolatti, G., & Grafton, S. T. (2010). Understanding actions of others: The electrodynamics of the left and right hemispheres. A high-density EEG neuroimaging study.,(8), e12160.
Proverbio, A. M., Riva, F., Paganelli, L., Cappa, S. F., Canessa, N., Perani, D., & Zani, A. (2011). Neural coding of cooperative vs. affective human interactions: 150 ms to code the action's purpose.,(7), e22026.
Sinke, C. B. A., Sorger, B., Goebel, R., & de Gelder, B. (2010). Tease or threat? Judging social interactions from bodily expressions.,(2), 1717–1727.
van Wount, M., & Sanfey, A. G. (2008). Friend or foe: The effect of implicit trustworthiness judgments in social decision-making.,(3), 796–803.
Vrticka, P., Andersson, F., Sander, D., & Vuilleumier, P. (2009). Memory for friends or foes: The social context of past encounters with faces modulates their subsequent neural traces in the brain.(5), 384–401.
Walter, H., Adenzato, M., Ciaramidaro, A., Enrici, I., Pia, L. & Bara, B. G. (2004). Understanding intentions in social interaction: The role of the anterior paracingulate cortex.,(10), 1854–1863.
Wang, Y. W., Huang, L., Lin, C., Zhang, Z., Liang, F., & Shen, D. (2012). Spatio-temporal brain dynamics of understandingsocial versus private intentions: An electrical neuroimaging study.,(4), 733–743.
Wang, Y. W., Huang, L., Xu, S., Yuan, B., Xu, Y. J., & Li, H. Y. (2012). ERP time course of understanding private versus communicative intentions.(12), 1618–1627.
[王益文, 黃亮, 徐晟, 袁博, 徐艷嬌, 李洪玉. (2012). 理解私人意圖與交際意圖的ERP證據.(12), 1618–1627.]
Wang, Y. W., Huang, L., Zhang, Z., Song, J., & Bai, L. Y. (2014). Kindness or hostility? Electrical brain dynamics of understanding interactive intentions of other people.,(7), 736–746.
[王益文, 黃亮, 張振, 宋娟, 白麗英. (2014). 友好還是敵對?理解互動意圖的大腦動態時程.(7), 736–746.]
Wang, Y. W., Huang, L., Zhang, W., Zhang, Z., & Cacioppo, S. (2015). Spatio-temporal dynamics of kind versus hostile intentions in the human brain: An electrical neuroimaging study.(3), 253–267.
Wang, Y. W., Lin, C. D., Yuan, B., Huang, L., & Shen, D. L. (2010). Person perception precedes theory of mind: An event related potential analysis.,(1), 238–246.
Wang, Y. W., Zheng, Y. W., Lin, C. D., Wu, J., & Shen, D. L. (2011). Electrophysiological correlates of reading the single-and interactive-mind.,, 64.
Wang, Y. W., Zheng, Y. W., Shen, D. L., Cui, L., & Yan, G. L. (2012). Reading another’s mind and reading the interactive mind from Chinese idioms: Evidience from eye-movements and ERPs.,(1), 100–111.
[王益文, 鄭玉瑋, 沈德立, 崔磊, 閆國利. (2012). 解讀成語中的他人心理與互動心理: 來自眼動和ERP的證據.,(1), 100–111.]
Watanabe, T., Yahata, N., Abe, O., Kuwabara, H., Inoue, H., Takano, Y., ... Yamasue, H. (2012). Diminished medial prefrontal activity behind autistic social judgments of incongruent information.(6), e39561.
Weymar, M., Bradley, M. M., Hamm, A. O., & Lang, P. J. (2013). When fear forms memories: Threat of shock and brain potentials during encodingand recognition.(3), 819–826.
Wieser, M. J., McTeague, L. M., & Keil, A. (2011). Sustained preferential processing of social threat cues: Bias without competition.(8), 1973–1986.
Wu, H., Chen, C., Cheng, D., Yang, S., Huang, R., Cacioppo, S., & Luo, Y-J. (2014). The mediation effect of menstrual phase on negative emotion processing: Evidence from N2.(3), 278–288.
Yoder, K. J., & Decety, J. (2014). The good, the bad, and the just: Justice sensitivity predicts neural response during moral evaluation of actions performed by others.(12), 4161–4166.
Young, L., & Saxe, R. (2009). Innocent intentions: A correlation between forgiveness for accidental harm and neural activity.(10), 2065–2072.
Brain spatio-temporal dynamics of understanding kind versus hostile intentions based on dyadic body movements
HUANG Liang; YANG Xue; HUANG Zhihua; WANG Yiwen
(Institute of Applied Psychology, Minnan Normal University, Zhangzhou 363000, China)(Institute of Psychological and Cognitive Sciences, Fuzhou University, Fuzhou 350116, China)(Center for China Social Trust Research, Fuzhou University, Fuzhou 350116, China)(College of Mathematics and Computer Science, Fuzhou University, Fuzhou 350116, China)
Previous social neuroscience studies focused mainly on the neural networks that sustain an understanding of a single individual’s neutral or negative intentions. However, few studies have explored those of different types of social interactive intentions performed by two or a number of individuals and their whole body movements. In addition, the spatio-temporal dynamics of understanding the kind and hostile intentions in the human brain also remain unclear.
In order to address the above issue, the electroencephalograph (EEG) technique was employed to explore the dissociative neural correlates of understanding kind versus hostile intention. Twenty healthy participants were recruited for the experiment. Their behavioral data (accuracy and reaction time) and electrical brain activities were recorded while they were watching colorful photos depicting two actors’ actions and performing an intention inference task (IIT). There were three different types of action intentions: kindness, hostility and non-interactiveness (neutrality). The ERP data was analyzed using the Scan and sLoreta software in an off-line way.
The Univariate Analysis of Variance (ANOVA) with repeated measures of mean accuracy showed no significant difference among three conditions, while a main effect of condition existed for reaction time. The reaction times of hostile intention were shorter than those of the kind and neutral intentions, while no significant difference was found between the latter two conditions. The ERP data were analyzed using a three-way repeated measure ANOVA. The ANOVA factors were intention condition (kindness, hostility and neutrality), laterality (left, midline and right areas) and caudality (frontal, central and parietal areas). Electrophysiological results showed, over the frontal area, a significantly more negative amplitude of N250 (170~270 ms) for neutral intention compared to kind and hostile intentions, and the N250 amplitudes for kind intention were also more negative than those for hostile intention, especially on the FZ electrode site. The source localization showed maximum activation in Broadman 10 (BA10), in the vicinity of middle frontal gyrus, for N250 for kind and hostile intentions. At the later stage (270~450 ms), the peak amplitudes of the P300 for hostile intention were more positive than those for the kind and neutral intentions, and the P300 amplitudes for kind intention were also more positive than those for neutral intention over the central, parietal areas as well as the right hemisphere. The maximum activation for P300 of kind and hostile intentions was found in BA45, located in the vicinity of insula, and a stronger activity existefor hostile intention compared to kind intention.
These findings show that there is a spatio-temporal dynamic dissociation between kind and hostile intentions understanding in the brain. Altogether, the current study provides electrophysiological evidence underlying the kind, hostile interactive intentions and non-interactive (neutral) intention understanding, and suggests a prioritized and sustained processing for hostile interactive intention. The study enriches the contents of the two-stage intention-understanding model and the putative Hierarchical & Multi-level Cognitive Framework (HMCF) in Theory of Mind.
Theory of Mind; kind intention; hostile intention; dyadic interaction; ERP
10.3724/SP.J.1041.2019.00557
2017-12-20
* 國家自然科學基金面上項目(31771238)研究成果之一, 并受福建省閩江學者支持計劃、福建省社會科學規劃青年項目(FJ2018C030)、福建省自然科學基金項目(2019J01753)、2017年福建省高校杰出青年科研人才培育計劃和漳州市2018年度中青年領軍人才訪學進修計劃資助。
王益文, E-mail: wangeven@126.com
B845
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
