背鰭缺失對胭脂魚幼魚游泳能力與反捕食能力的影響
2019-05-21 08:58:56周龍艷付世建李秀明
淡水漁業(yè) 2019年3期
周龍艷,付世建,付 成,凌 鴻,李秀明
(重慶師范大學進化生理與行為學實驗室,重慶市動物生物學重點實驗室,重慶401331)
游泳運動是魚類逃逸、捕食和繁殖等行為實現(xiàn)的主要方式,為魚類的生存、生長和繁衍提供基本保證[1]。魚類游泳行為復雜多變,且存在顯著的種間和種內分化。因此,需要選擇合適的指標對魚類的游泳能力進行客觀評價。一直以來,臨界游泳速度(critical swimming speed,Ucrit)和最大勻加速游泳速度(constant acceleration test speed,Ucat)被研究者廣泛應用于魚類游泳能力的評價[2]。Ucrit作為魚類有氧運動能力的重要評價指標[3],主要受到最大有氧代謝能力(maximum metabolic rate, MMR)的限制。而Ucat作為魚類無氧運動能力的重要評價指標[3]。二者對于魚類在自然環(huán)境條件下逃避捕食、穿越激流以及尋找適宜生境等活動密切相關,具有重要的生態(tài)學意義。另外,最近的研究發(fā)現(xiàn)盡管運動時間短(不足1 s)且移動距離有限,快速啟動逃匿過程(fast-start response)的運動性能對魚類有效捕獲獵物和逃避敵害至關重要[4]。因此,該過程的最大速度、最大加速度、旋轉角度以及旋轉率(后二者為靈活性的指標)可能與魚類的生存適合度緊密關聯(lián)[5]。
魚鰭是魚類游泳的重要器官之一,主要包括胸鰭、腹鰭、臀鰭、背鰭以及尾鰭。不同魚鰭在魚類游泳過程中發(fā)揮著截然不同的作用。作為游泳運動的重要部位,尾鰭與魚類運動能力的關系一直受到研究者們的廣泛關注[6-7]。而背鰭對魚類游泳能力的影響卻鮮有涉及。胭脂魚(Myxocyprinusasiaticus)屬鯉形目(Cypriniformes)胭脂魚科或稱亞口魚科(Catostomidae),該科魚類全世界約有 13 屬70 種,絕大多數(shù)種類分布于北美洲,但是胭脂魚在我國為特有種,分布于我國的長江和閩江流域[8]。因此,胭脂魚在魚類系統(tǒng)分類和動物地理學研究上有著極為重要的科學價值。相關的調查報道指出,目前胭脂魚在閩江已近絕跡,長江種群的資源也日趨枯竭,目前已經被列為國家二類水生野生保護動物[9]。不同生長階段的胭脂魚體形差異明顯。幼魚階段體型奇特,體較高,呈三角形,背鰭寬展而高大,有“一帆風順”的寓意[10-11]。本研究主要探討背鰭在胭脂魚幼魚游泳運動和反捕食過程中的作用,為其資源保護提供重要的參考資料。
1 材料與方法
1.1 實驗魚的來源與馴養(yǎng)
胭脂魚幼魚(Myxocyprinusasiaticus)于2017年11月購自重慶市北碚區(qū),體質量(2.30±0.12) g, 體長(4.59±0.09) cm,共124尾,在實驗室自凈化循環(huán)控溫水槽中馴養(yǎng)21 d,曝氣24 h后的自來水作為實驗用水。馴養(yǎng)期間每天10: 00采用商業(yè)顆粒飼料飽足投喂一次,30 min后用虹吸管排除殘餌與糞便;水槽日換水量約為水體(200 L)的10%,水溫控制在(25±1)℃,并用充氧泵供氧。馴養(yǎng)結束后,挑選身體健康、體重、體長接近的實驗魚作為實驗對象。
捕食者烏鱧(Ophiocephalusargus)購于重慶市永川區(qū)漁場。購回的烏鱧放入(25±1)℃的水槽中馴養(yǎng)14 d,期間用曝氣24 h后的自來水馴養(yǎng)。
1.2 實驗方案
1.2.1 實驗設計
剪鰭處理:將實驗魚輕度麻醉(3-氨基苯甲酸乙酯甲基磺酸鹽, MS-222,50 mg/L)后用剪刀進行剪背鰭處理(圖 1)。切除背鰭處理后的實驗魚放回水體恢復2 d[5]。
游泳能力測定:選取實驗魚60尾用于3種游泳能力的測定,每種游泳能力取20尾測定,其中對照組10尾背鰭缺失組10尾。
捕食死亡率:實驗設4個重復,每個重復選取實驗魚16尾用于捕食死亡率的測定,其中8尾進行背鰭切除,另外8尾作為對照組。
1.2.2Ucrit的測定
采用實驗室自行研制的游泳代謝測定儀測定Ucrit[12]。具體操作為首先將單尾實驗魚放入測定儀中適應2 h,適應過程的水流速度為4. 0 cm/s(約1 倍體長/秒,Body Length,1 BL/s);隨后以4 cm/s的速度增量提高水流速度至 8、12、16和20 cm/s等,各水流速度下的持續(xù)游泳歷時均為20 min[1]。重復上述操作直至實驗魚表現(xiàn)出力竭狀態(tài)即停止實驗。力竭的判斷標準為實驗魚被水流沖至游泳管尾端篩板,且身體呈卷曲狀態(tài)的時間不低于20 s[13]。計算公式為:
Ucrit=[V+(t/ΔT)ΔV]
(1)
式中:V為順利完成設定時間的最大游泳速度(力竭前一檔速度);t為未能完成當前速度梯度下設定時間的實際持續(xù)游泳時間(t<20 min);ΔT為各速度梯度下的持續(xù)時間(即20 min);ΔV為各速度梯度的速度增量(即 4 cm/s)。
運動代謝率在測定Ucrit的過程中進行測定。具體操作為每2 min測定 1 次游泳代謝儀封閉水體的溶氧值,以溶氧值隨時間變化斜率的絕對值和水體的體積計算每尾實驗魚的運動過程的耗氧率(MO2)
作為代謝率的指標。實驗結束后將實驗魚取出,對游泳代謝儀重新密閉并進行 20 min 的耗氧率測定,用以消除細菌耗氧對實驗魚MO2的影響[13],計算公式為:
MO2=(St-S0)×3.45× 60/(W× 1 000)
(2)
式中MO2(mgO2/(kg·h))代表耗氧率,St為每檔期間(20 min)溶氧值隨時間變化的斜率,S0為細菌耗氧的溶氧值變化斜率,3.45 為游泳代謝儀體積(L), 60 為 60 min。W表示實驗魚的體重(g)。
1.2.3Ucat的測定
Ucat測定的設備與Ucrit基本相同,主要改進之處為調節(jié)流速的方式改變?yōu)橥ㄟ^智能化調節(jié)輸出電源的頻率而改變電機的轉速。具體操作如下:將實驗魚轉入游泳代謝測定儀中馴化適應2 h,流速為4 cm/s。馴化結束之后,將測定儀中的水流速度以恒定的加速度(10 cm/s2)持續(xù)均勻增加水流速度,實驗魚力竭時的水流速度即為Ucat [14]。
1.2.4 快速啟動游泳能力的測定
實驗前將直徑 1 mm 的白色圓形紙片固定于實驗魚背部中央皮膚表層,作為實驗魚的質心標記點[15],以便進行圖像分析和參數(shù)計算。拍攝整個快速啟動過程并計算快速啟動指標。實驗結束后,用圖像處理軟件(ACD see v 8. 0)對拍攝的圖像進行處理并采用軟件(tpsdig2)進行數(shù)據采集和分析。通過計算,分別求得實驗魚質心的最大線加速度(Amax)、速度(Vmax)以及旋轉率和旋轉角度,參數(shù)計算詳見相關參考文獻[4,16]。
1.2.5 捕食死亡率的測定
將對照組和背鰭缺失組各8尾實驗魚一同轉移至提前放置有2尾捕食者的捕食測定水槽中(長1.5 m×寬0.8 m×高0.5 m)。在實驗過程中,每3 h觀察一次捕食情況,當一半的胭脂魚(即8尾)被捕食時,停止實驗。設置4個重復。捕食死亡率的計算為被捕食的對照組或背鰭缺失組個體占總被捕食個體的百分比[17]。
1.2 統(tǒng)計分析
實驗數(shù)據采用SPSS 17進行統(tǒng)計分析。統(tǒng)計值均以平均值±標準誤(Mean±SE)表示,顯著水平為P<0.05。對照組和背鰭缺失組之間各參數(shù)的差異使用t檢驗進行統(tǒng)計分析。背鰭缺失和游泳速度對運動代謝率(MO2)的影響采用重復度量的雙因素方差分析。
2 結果
2.1 背鰭缺失對胭脂魚幼魚Ucrit及其代謝率、Ucat和快速啟動能力相關參數(shù)的影響
對照組與背鰭缺失組的Ucrit及各代謝參數(shù)均無顯著性差異(表 1)。隨著水流速度的增加,對照組和背鰭缺失組的MO2均顯著上升,但對照組和背鰭缺失組在同一游泳速度下的MO2均無顯著差異(圖2),暗示其組間游泳效率無顯著差異。對照組與背鰭缺失組胭脂魚幼魚的Ucat沒有顯著性差異,分別為(53.12±3.02)和(52.92±1.52) cm/s(表 1)。對照組和背鰭缺失組的Vmax分別為(1.12±0.08),(1.11±0.05) mm/ms;Amax分別為(0.22±0.02),(0.19±0.01) mm/ms2;旋轉角度分別為(54.91±3.3)(°),(59.66±7.33)(°);旋轉率分別為(1 390.10±171.03,(1 762.51±266.83)(°)/s。對照組與背鰭缺失組快速啟動各個參數(shù)均無顯著差異(表 1)。
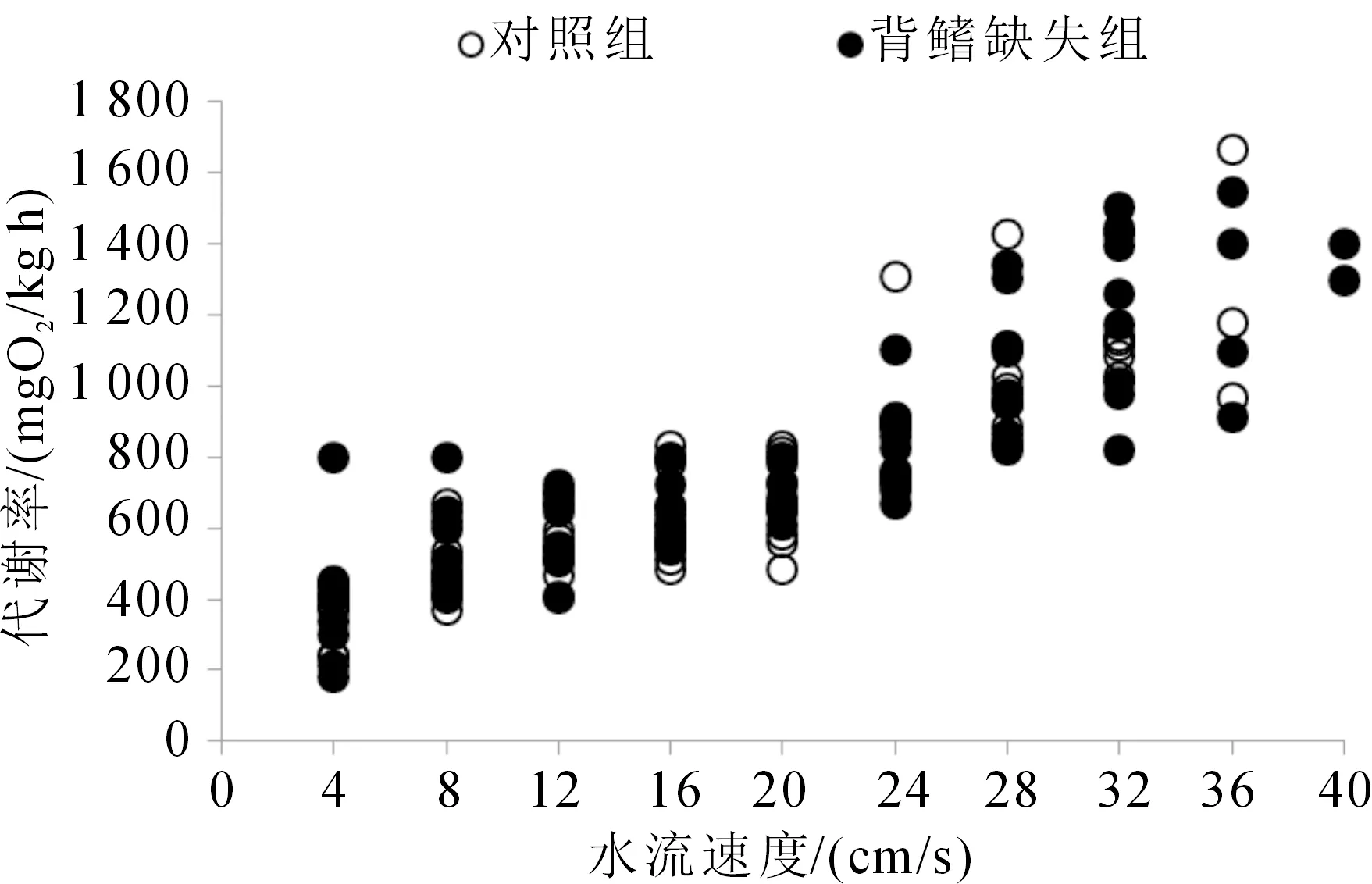
圖2 背鰭缺失對胭脂魚幼魚游泳過程中耗氧率的影響
2.2 背鰭缺失對胭脂魚幼魚在捕食者存在條件下存活率的影響
從圖3可以看出,對照組胭脂魚幼魚在捕食者存在條件下的存活率為62.5%,而背鰭缺失組胭脂魚幼魚在捕食者存在條件下的存活率為40.6%。背鰭缺失對胭脂魚幼魚在捕食者存在條件下存活率具有顯著影響,背鰭缺失組胭脂魚幼魚的存活率顯著低于對照組(P<0.05)。
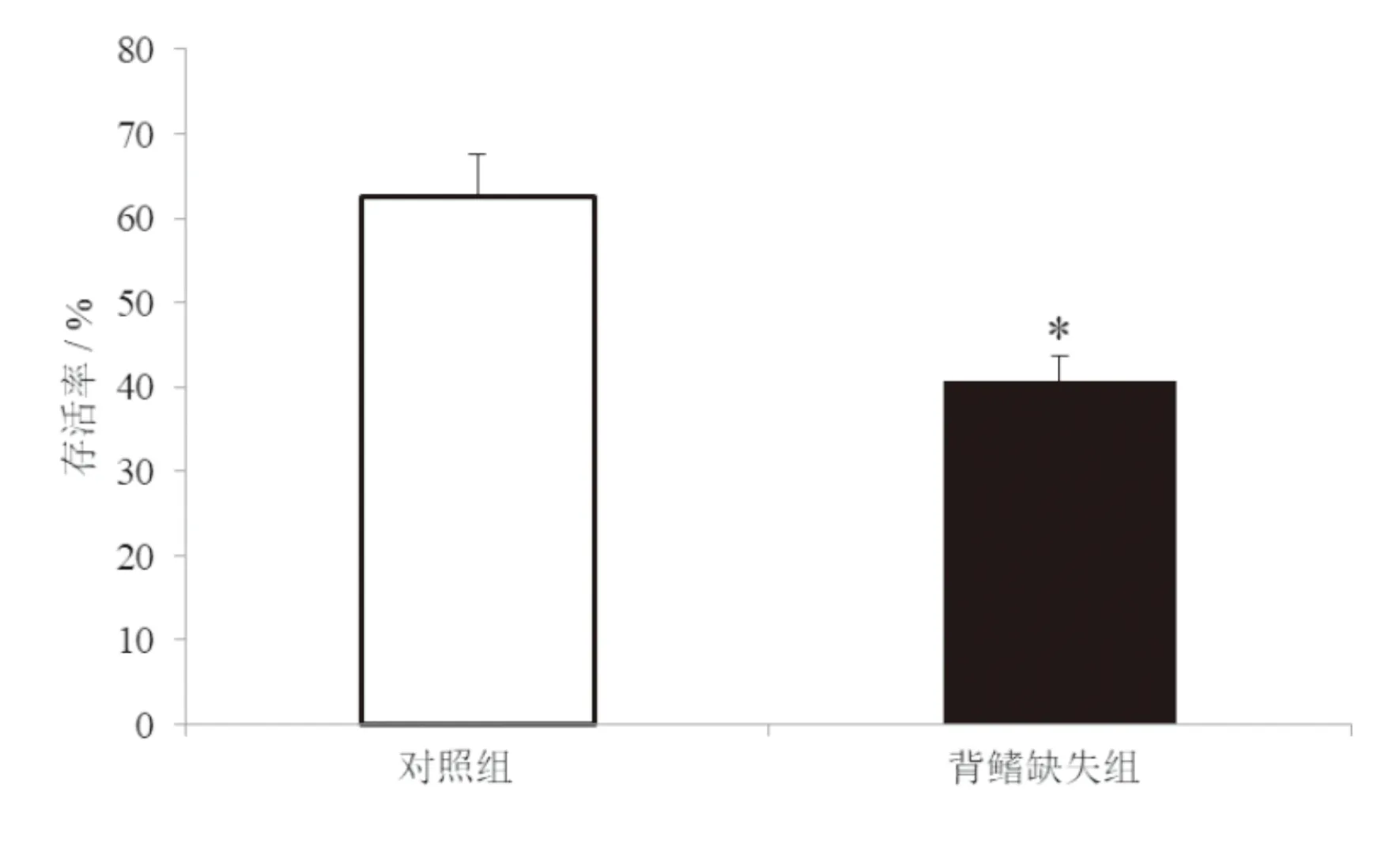
圖3 背鰭缺失對胭脂魚幼魚反捕食能力的影響
3 討論
魚鰭在魚類游泳過程中起著重要的作用,不同類型、大小和形態(tài)的魚鰭在魚類運動過程中的作用不盡相同[18]。尾鰭作為運動系統(tǒng)的重要器官, 在游泳過程中具有推進及導向作用,因此大部分研究主要關注魚類尾鰭與運動能力的關系[5-7]。尾鰭的缺失導致鳊(Parabramispekinensis)、金魚(Carassiusauratus)、中華倒刺鲃(Spinibarbussinensis)和鯉(Cyprinuscarpio)幼魚的Ucrit分別下降40%、35%、49% 和 32%[7,19-20]。由于胭脂魚幼魚具有特殊的背鰭形態(tài),可能對其游泳能力產生一定的影響。不過,相關研究還未見報道。本研究通過比較對照組和背鰭缺失組胭脂魚幼魚之間Ucrit發(fā)現(xiàn),背鰭缺失對胭脂魚幼魚持續(xù)游泳速度沒有顯著的影響(表 1)。這與唐魚(Tanichthysalbonubes)切除背鰭后的研究結果相似[20]。此外,由于魚類的游泳能力與其能量代謝能力密切相關,一些研究者也對鰭缺失后魚體耗氧率進行了研究[6,19]。不過,以往的研究發(fā)現(xiàn)尾鰭缺失并沒有對紅大馬哈魚(Oncorhnchusnerka)和鳊的MMR產生顯著影響[6,19]。本研究發(fā)現(xiàn),背鰭缺失處理后胭脂魚幼魚的RMR、MMR和MS等代謝參數(shù)也都沒有產生顯著性變化(表 1)。由此表明,背鰭的缺失并沒有對胭脂魚幼魚游泳代謝能力產生影響(圖 1),這可能是其游泳能力沒有產生顯著變化的原因之一。總體而言,魚類背鰭在游泳過程的作用遠小于尾鰭,胭脂魚進化過程中形成的獨特的背鰭形態(tài)及其生態(tài)學作用值得進一步研究。
魚類生存能力與其游泳能力密切相關,特別是魚類非穩(wěn)定游泳能力(通常用Ucat和快速啟動過程中的Vmax、Amax、旋轉角度以及旋轉率等指標表示)在魚類捕食和避敵過程中發(fā)揮著重要的作用[3-5]。研究發(fā)現(xiàn),尾鰭缺失導致鯽(Carassiusauratus)和寬鰭鱲(Zaccoplatypus)快速啟動能力都顯著下降,最大線速度和旋轉半徑分別下降39% 和62%[5]。本研究發(fā)現(xiàn),盡管背鰭缺失對胭脂魚幼魚非穩(wěn)定游泳能力指標(Ucat、Vmax、Amax、旋轉角度和旋轉率)沒有顯著影響(表 1),卻導致其捕食者存在條件下的存活率顯著下降 35%(圖 2)。由此表明,背鰭可能對胭脂魚幼魚的反捕食能力具有重要的作用,但其作用并非通過影響其非穩(wěn)定游泳能力的方式表現(xiàn)出來,具體機制值得進一步研究。先前的研究表明捕食者的口裂大小與獵物的大小密切相關,口裂大小是影響魚類捕食獵物的一個重要因素[21-23]。因此,胭脂魚幼魚高高的背鰭可能是其形態(tài)上針對捕食者口裂限制的反捕食策略之一。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
發(fā)明與創(chuàng)新(2022年30期)2022-10-03 08:40:56
中學生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
