鮑魚腸道海帶降解菌株的篩選、鑒定及粗酶學性質研究
2019-05-23 03:36:22姚艷艷劉心田常麗榮錢浩王曉輝車建鋒李長青
食品與發酵工業 2019年9期
姚艷艷,劉心田,常麗榮*,錢浩,王曉輝,車建鋒,李長青
1(威海長青海洋科技股份有限公司,國家海產貝類工程技術研究中心,山東 榮成,264300) 2(威海市漁業技術推廣站,山東 威海,264200)
海帶(LaminariajaponicaAresch)是一種重要食品、肥料、飼料及工業原料,有著很高的經濟價值。據統計,2017年,我國經濟海藻養殖總量達223.5萬t,其中海帶(鮮品)年產量達148.7萬t[1]。而目前國內對海帶的加工方式還是以自然曬干、烘干、鹽漬等簡單加工為主,無論哪種方式,都面臨著環境污染嚴重、高耗能、勞動密集、附加值低等嚴峻問題。
近年來,海帶提取物相關技術、產品陸續涌現,尤其是在農業[2]、食品[3]、化妝品[4]、保健品[5]、醫藥[6]等領域表現出了廣闊的應用前景。工業上,海帶的提取加工方法主要有化學法、物理法、生物法等[7-8]。化學法和物理法存在能耗高、污染環境的缺點;而生物發酵降解技術是利用微生物產生的褐藻膠裂解酶、纖維素酶、蛋白酶、淀粉酶等來降解藻體,是海帶提取加工產業的創新應用,具有反應條件溫和、節約能耗、綠色環保等特點,是海帶及其他經濟海藻提取加工的新技術。目前,國內外對海帶的生物降解方法仍以生物酶解技術為主[9-10],存在降解得率低、資源浪費大、酶制劑成本高等問題。只有少量文獻報道利用復合菌株對海帶進行發酵降解[11-12],而已報道的產酶菌株種類較少,且產酶較單一,產酶能力低[13-15],難以滿足工業生產的需求。
皺紋盤鮑主要以海帶為食,其腸道內含有多種益生菌及產褐藻膠裂解酶的菌株[16-18],但從鮑魚腸道內篩選應用于降解海帶的微生物尚未見報道。本試驗從鮑魚腸道中,以篩選產褐藻膠裂解酶菌株為出發點,進一步篩選能夠對海帶進行發酵降解的海洋源菌株,最大程度地提取和利用以褐藻膠為主的海帶有效成分,提高海帶降解得率。
1 材料與方法
1.1 材料與試劑
成熟鮑魚:威海長青海洋科技股份有限公司;褐藻膠裂解酶:1 000 U/mL,國家海產貝類工程技術研究中心;復合酶:20 000 U/mL,諾維信創新與發展中心;血液瓊脂基礎,脫纖維羊血:青島高科園海博生物技術有限公司;細菌基因組DNA提取試劑盒:寶日醫生物技術(北京)有限公司。
褐藻酸鈉,MgSO4·7H2O,FeSO4·7H2O,(NH4)2SO4,KH2PO4,K2HPO4,蛋白胨,酵母粉,可溶性淀粉,FePO4,KCl,CuSO4,MgCl2:分析純,國藥化學試劑有限公司。
1.2 儀器與設備
AL104分析天平,梅特勒-托利多儀器有限公司;L535-1低速離心機,湖南湘儀實驗室儀器開發有限公司;UV2800S紫外分光光度計,上海舜宇恒平科學儀器有限公司,HFsafe-1200生物安全柜,上海力申科學儀器有限公司;DHZ大容量恒溫振蕩培養箱,上海精宏實驗設備有限公司;SHP-250型生化培養箱,上海森信實驗儀器有限公司;STTIK高壓滅菌鍋,施都凱儀器設備(上海)有限公司;BIOER XP基因擴增儀,杭州博日科技有限公司;SC80S凝膠成像系統,上海山富科學儀器有限公司。
1.3 培養基
富集培養基:褐藻酸鈉5 g,蛋白胨2 g,MgSO4·7H2O 0.01 g,FeSO4·7H2O 0.5 g,過濾海水1 000 mL,調節初始pH至7.2,121 ℃滅菌20 min。
篩選培養基:褐藻酸鈉5 g,(NH4)2SO42 g,KH2PO43 g,K2HPO47 g,NaCl 30 g,MgSO4·7H2O 0.1 g,FeSO4·7H2O 0.05 g,瓊脂15 g,蒸餾水1 000 mL,調節初始pH至7.0,121 ℃滅菌20 min。
液體發酵培養基:見參考文獻[19]。
血平板:配制100 mL的血液瓊脂基礎培養基于121 ℃滅菌15 min,冷卻至50 ℃,無菌操作加入10 mL無菌的脫纖維羊血,搖勻,傾注平板,凝固后,4 ℃保存備用。
蛋白酶培養基:脫脂奶粉30 g,瓊脂15 g,陳海水1 000 mL,調節培養基初始pH至7.6,121 ℃滅菌20 min。
淀粉酶培養基、纖維素酶培養基:見參考文獻[20]。
1.4 菌株的篩選
1.4.1 菌種富集及初篩
取鮮活的鮑魚腸道作為實驗樣品,用研缽研磨后,加入裝有100 mL富集培養基的三角燒瓶中,于25 ℃下富集培養16 h,將富集后的樣品梯度稀釋至10-4,取各梯度下樣品100 μL涂布篩選培養基,25 ℃下培養2~5 d。
1.4.2 溶血素的產生
將2216E培養基過夜培養的待測菌株點種于血平板上,25 ℃培養24 h,根據菌落周圍的溶血圈情況,來判斷溶血素的產生。
1.4.3 產酶能力篩選
用接種環挑取純化后的菌株單菌落,分別點種于蛋白酶培養基、淀粉酶培養基和纖維素酶培養基,25 ℃培養24~48 h。菌落周圍產生透明圈的菌株為陽性,不產透明圈的菌株為陰性。
淀粉酶培養基顯色:輕輕刮下菌苔,用蒸餾水或1 mol/L NaCl溶液沖洗培養基表面,用盧戈氏碘液染色5 min。
纖維素酶培養基顯色:輕輕刮下菌苔,用蒸餾水或1 mol/L NaCl溶液沖洗培養基表面,用0.1%剛果紅染液染色30 min,再用1 mol/L NaCl溶液洗脫1~2次,每次浸泡20 min。
1.5 菌種鑒定
1.5.1 菌株菌落形態觀察
菌株在篩選平板上25 ℃培養48 h后,觀察平板上菌落形態,并進行革蘭氏染色,觀察細菌形態。
1.5.2 16S rRNA分子鑒定
以提取菌株的總DNA為模板,以27F和1 492R為引物[21],進行Touchdown-PCR擴增反應,反應條件為94 ℃預變性5 min,94 ℃變性30 s,60 ℃下退火30 s,72 ℃延伸1 min 30 s,每個循環降低1 ℃,共進行15個循環;再進行94 ℃變性30 s,45 ℃下退火30 s,72℃延伸1 min 30 s,共進行25個循環;最后在72 ℃下延伸10 min。凝膠回收目的片段后,送樣測序。
將得到的測序結果在EzBioCloud網站上進行序列比對,并找出與之同源性接近的不同菌株序列,應用MeGA 5.0軟件進行聚類與同源性分析。遵循鄰接法(neighbor-joining method)原則,并通過自舉分析(boostrap)進行置信度檢測,自舉數據集為1 000次,構建分支系統發育樹。
1.6 褐藻膠裂解酶酶活力的測定
25 mL比色管中加入2 mL含1.5% NaCl的0.5% 褐藻酸鈉溶液,在30 ℃水浴中放置10 min,再加入10~100 μL酶液,振蕩混勻后于30 ℃水浴中反應30 min,加入3 mL DNS試劑混勻,沸水浴5 min后立即冷卻至室溫,定容至25 mL,在540 nm處檢測光吸收值。根據葡萄糖標準曲線確定產生還原糖的量。酶活力單位定義為:30 ℃下每分鐘產生1 μg的還原糖所需要的酶量。發酵液酶活力定義為:1 mL發酵液含酶活單位(U/mL)。
1.7 菌體生長和產酶曲線的繪制
在液體發酵培養基中接種3次傳代后的菌株,在預實驗最佳的培養條件下培養,在菌株生長過程中每間隔5 h取樣1次,測定發酵液的OD600值及酶活力值大小,繪制出菌株的生長和產酶曲線。
1.8 褐藻膠裂解酶酶學性質研究
利用硫酸銨分級鹽析法制備粗酶,并用20 mmol/L Tris-HCl緩沖液(pH 7.5)溶解。
最適溫度的測定:在25、30、35、40、45、50 ℃下測定酶活力,其中底物濃度5‰,反應pH 7.0。
最適pH的測定:在pH 6.5、6.8、7.0、7.3、7.5、8.0、8.5下測定酶活力,其中底物濃度5‰,酶解溫度是35 ℃。
金屬離子對酶活力的影響:在反應體系中加入K+(KCl)、Fe2+(FeSO4·7H2O)、Cu2+(CuSO4)、Na+(NaCl)、Mg2+(MgCl2)、Ca2+(CaCl2)下測定酶活力,其中底物濃度5‰,酶解溫度是35 ℃,反應pH 7.5。
1.9 海帶降解得率
選取100目海帶粉,海帶粉與水的料液質量比為1∶12,在菌株產生的褐藻膠裂解酶最佳酶解條件下,利用菌株發酵液、褐藻膠裂解酶、復合酶對海帶發酵降解6 h,并計算海帶降解得率。
海帶降解得率是指干海帶發酵后溶于水中的可溶性固形物占干海帶的百分含量,按公式(1)計算:
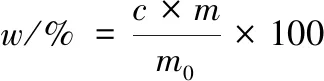
(1)
式中:w,海帶發酵得率,%;c,海帶發酵上清液的可溶性固形物的百分含量,%;m,發酵后所得海帶發酵上清液質量,g;m0,發酵用的干海帶質量,g。
2 結果與分析
2.1 海帶降解菌株的篩選
通過以褐藻酸鈉為唯一碳源的選擇性培養基初篩,從鮑魚腸道中篩選得到100多株菌,其中有12株是產褐藻膠裂解酶的菌株,通過搖床振蕩發酵培養,測定不同菌株發酵上清液的酶活力大小,結果如表1所示。其中YY7菌株的酶活力最高,且經過多次傳代培養后,其遺傳穩定性及產酶活力穩定,因此,將此菌株進行菌種鑒定及酶學性質研究。
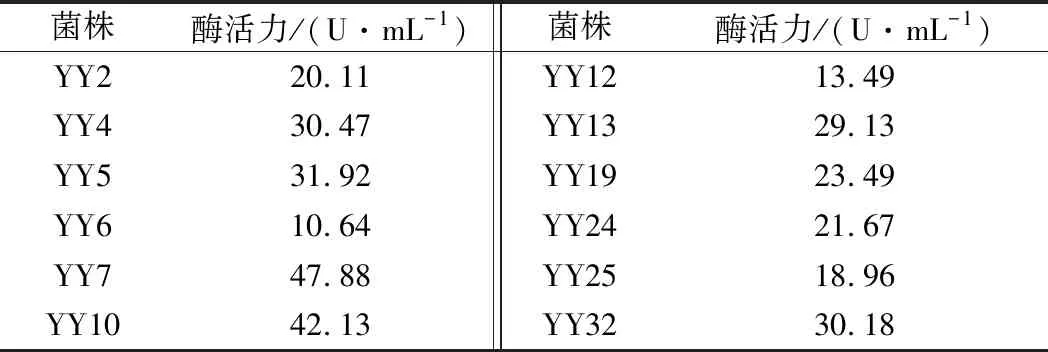
表1 不同菌株產褐藻膠裂解酶活力對比Table 1 The alginate lyase activity in different strains
2.2 菌種的鑒定
菌株YY7在篩選平板上培養48 h后,菌體周圍產生明顯水解透明圈,單菌落呈現不透明圓形,邊緣整齊,顏色微白,直徑約1.5 mm,邊緣整齊,表面光滑、濕潤,稍有隆起,顯微鏡下觀察為彎桿狀,革蘭氏陰性菌。
通過分子生物學手段,鑒定菌株YY7的16S rRNA基因序列為1 476 bp,在EzBioCloud網站上經過序列比對,發現其16S rRNA與弧菌屬Vibrioalgivorus的SA2(T)菌株的相似性最高,為99.72%,依據16S rRNA序列構建系統發育樹聚類分析(圖1),結合菌株的形態學特征觀察,初步鑒定為弧菌屬菌株,并命名為Vibriosp. YY7。
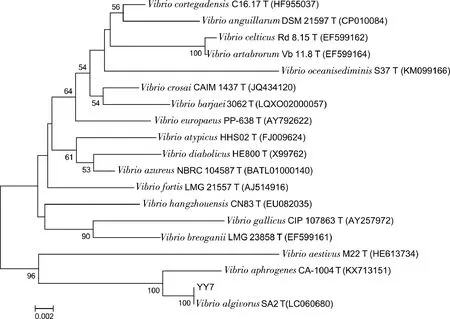
圖1 YY7菌株的16S rRNA序列系統發育樹Fig.1 Phylogenetic tree based on 16S rRNA sequenceof YY7 stains
將YY7菌株點種于血平板上培養,并沒有產生溶血現象,表明YY7菌株不具有潛在致病性。通過其他培養基鑒定YY7菌株的產酶情況后,發現YY7菌株在蛋白酶培養基(圖2-a)和淀粉酶培養基(圖2-b)上均有透明圈,說明菌株YY7還具有蛋白酶和淀粉酶活性。水產動物益生菌的篩選大多通過體外篩選產酶菌株獲得[22],因此推斷YY7菌株可能是鮑腸道內的潛在益生菌。
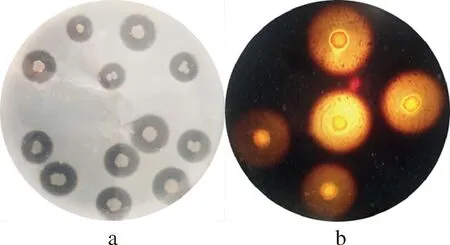
a-蛋白酶培養基; b-淀粉酶培養基圖2 YY7菌株在蛋白酶培養基和淀粉酶培養基的產酶情況Fig.2 The enzyme production of YY7 strains on protease medium and amylase medium
2.3 菌體生長曲線和產酶曲線的繪制
對YY7菌株進行生物量監測和產褐藻膠裂解酶曲線的繪制,如圖3。由圖3可見,菌體的生長隨時間的延長逐漸升高,在18 h后開始進入穩定生長期,菌株生物量接近最大值,酶活力在20 h達到最高值,為54.31 U/mL,與已報道的弧菌Vibriosp. QY107[14]、Vibriosp. SS-1[18]等菌株相比,菌株YY7的發酵產酶周期更短,生產速度較快,在工業化生產中,極大縮短了發酵時間,從而降低雜菌污染的風險,節約了生產成本,具有極大的生產優勢。
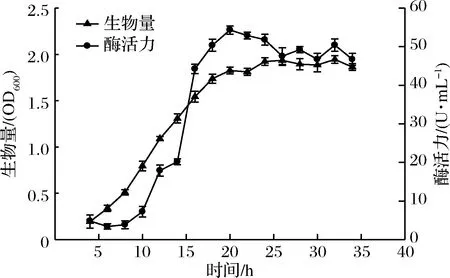
圖3 YY7菌株的生長曲線及酶活力曲線Fig.3 Growth curve and enzyme activity curve of YY7 strains
2.4 褐藻膠裂解酶酶學性質研究
2.4.1 褐藻膠裂解酶最適反應溫度的確定
將酶反應體系分別置于25、30、35、40、45、50 ℃的水浴中反應,測定不同溫度下褐藻膠裂解酶的活力,結果見圖4。從圖4可以看出,隨著溫度升高,酶活力逐漸升高,35 ℃達到最高值,40 ℃時酶活力略有降低,之后溫度越高,酶活力逐漸降低,可能是溫度越高,對酶的破壞越大,從而活力逐漸降低。故該酶的最適反應溫度為35 ℃。
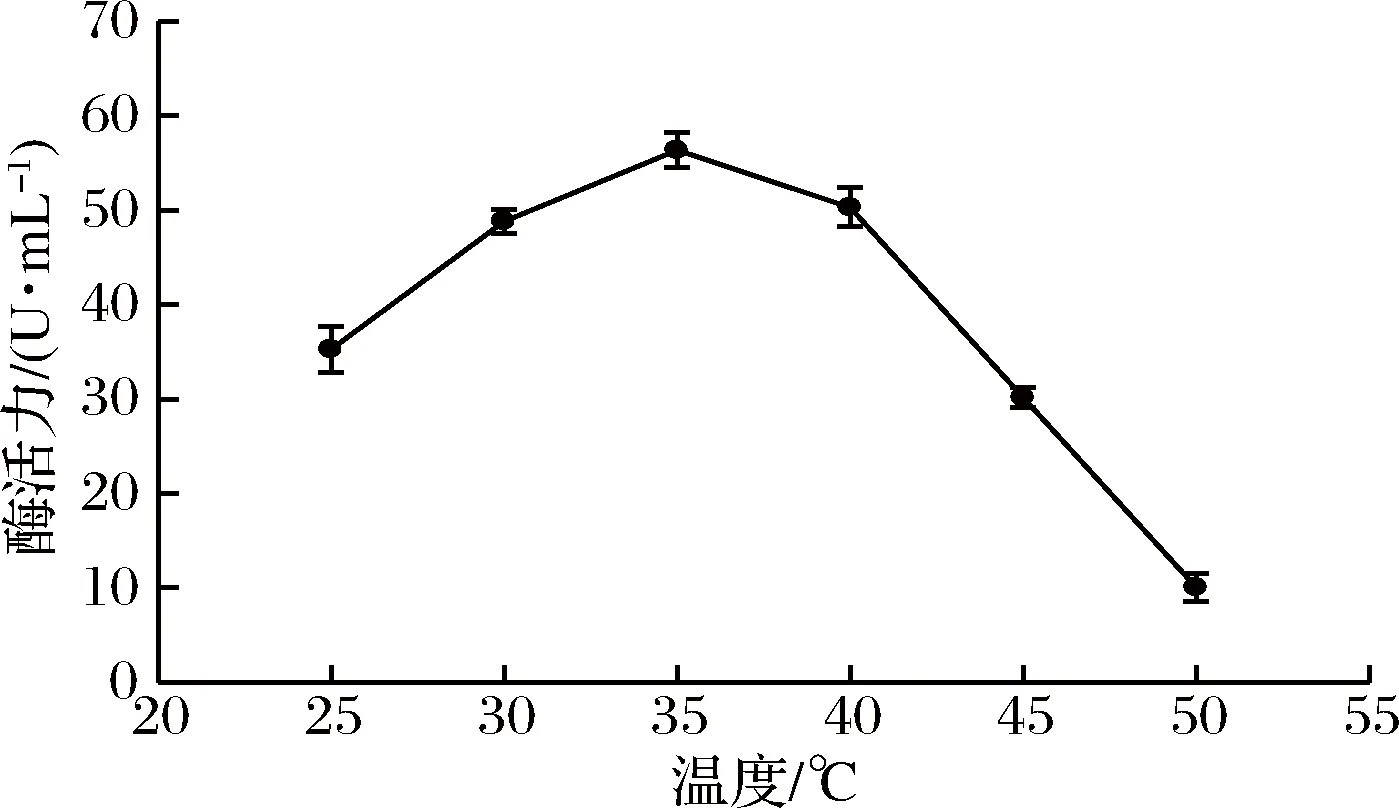
圖4 溫度對酶活力的影響Fig.4 Effect of temperature on enzyme activity
2.4.2 褐藻膠裂解酶最適pH確定
酶解反應體系中pH是影響酶的結構和穩定性的重要因素,同時也影響底物和酶的解離狀態[23]。為探究反應體系pH對褐藻膠裂解酶活力的影響,選取pH 6.5、6.8、7.0、7.3、7.5、8.0、8.5七個梯度進行酶活力測定,結果見圖5。由圖5可知,褐藻膠裂解酶活力隨著體系pH的增大呈現先增大后減小趨勢,在pH 7.5附近達到最大值,繼續增大pH,褐藻膠裂解酶活力反而下降,可能是堿性環境影響了酶與底物的結合,也可能是在堿性環境下酶蛋白的結構和穩定性都遭到了破壞。因此該酶的最適反應pH為7.5。
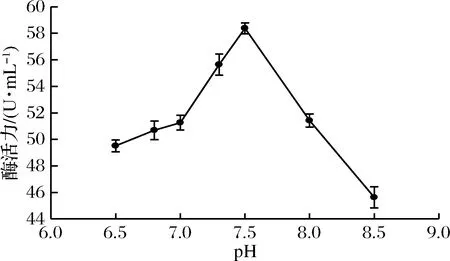
圖5 pH對酶活力的影響Fig.5 Effect of pH on enzyme activity
2.4.3 金屬離子對褐藻膠裂解酶酶解反應的影響
選取5種金屬離子研究它們對褐藻膠裂解酶活力的影響,結果見圖6。從圖6可以看出Na+、Mg2+、K+、Ca2+對酶活力都有不同程度的促進作用,此結果與李麗妍[24]的結果一致,各金屬離子對酶活力的促進作用由高到低依次為Na+>Mg2+>K+>Ca2+,Na+對酶活力的促進作用最大,可能與產酶菌株來自高鹽的海洋環境有關[24],且褐藻膠裂解酶對Na+的依賴性較高[25]。而Fe2+和Cu2+對褐藻膠裂解酶活力起抑制作用。因此,在褐藻膠裂解酶的應用過程中,可以根據需求在反應體系中添加Na+、Mg2+、K+、Ca2+,提高酶活力。
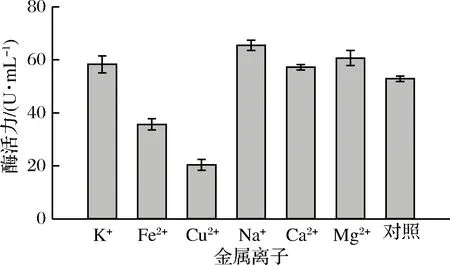
圖6 金屬離子對酶活力的影響Fig.6 Effect of metal ion on enzyme activity
2.5 海帶降解效果分析
2.5.1 海帶發酵過程中總糖及還原糖含量變化
海帶粉與水按1∶12比例攪拌均勻,將反應體系pH調至7.5±0.1后,加入1 000 U發酵液或酶,于35 ℃下酶解6 h。YY7發酵液降解海帶過程中總糖及還原糖含量的變化情況見圖7。由圖7可見,海帶在酶解發酵過程中,反應體系中的總糖和還原糖含量逐漸升高,這可能與菌株YY7發酵液中各種酶的作用有關,海帶中褐藻膠、海藻淀粉、海藻多糖等大分子物質都有可能被部分降解。
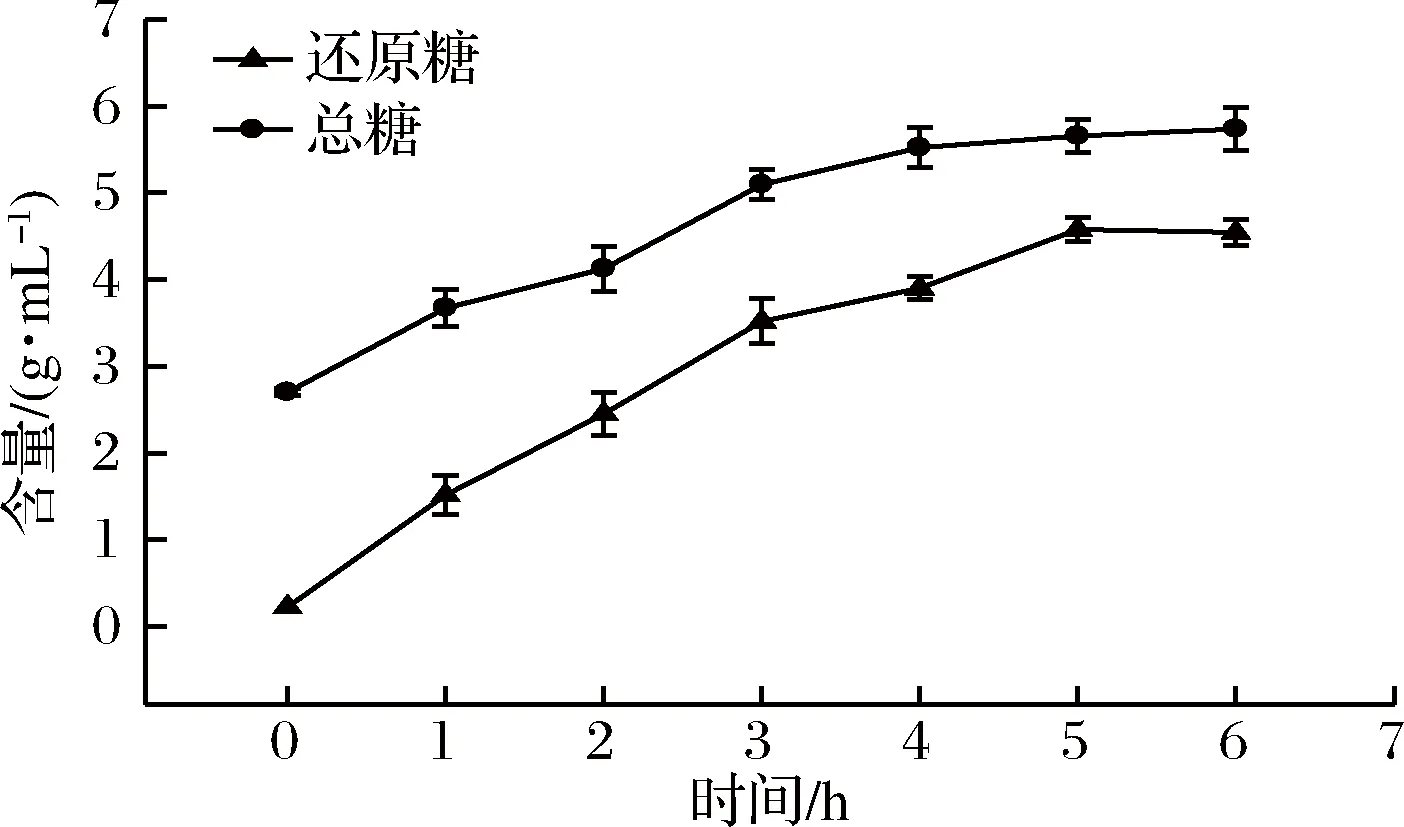
圖7 海帶發酵過程中總糖及還原糖含量變化Fig.7 Content changes of total sugar and reducing sugar during fermentation of Laminaria japonica Aresch
2.5.2 海帶降解得率對比分析
海帶降解得率反應海帶降解效果,海帶降解得率對比見圖8。與褐藻膠裂解酶和復合酶相比,YY7發酵液對海帶的降解效果最佳,海帶降解得率可達到65%以上,表明YY7發酵液對海帶降解具有較好的效果。
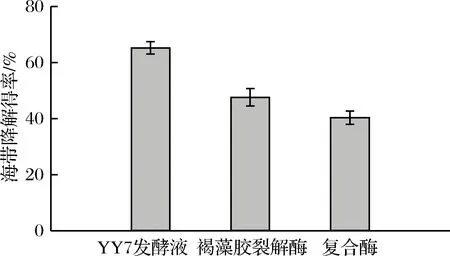
圖8 不同酶對海帶降解得率的影響Fig.8 Effect of different enzymes on degradation yield ofLaminaria japonica Aresch
2.5.3 海帶降解前后的顯微結構差異
海帶粉通過YY7發酵液降解前后,海帶粉的顯微結構發生明顯的變化,結果見圖9。
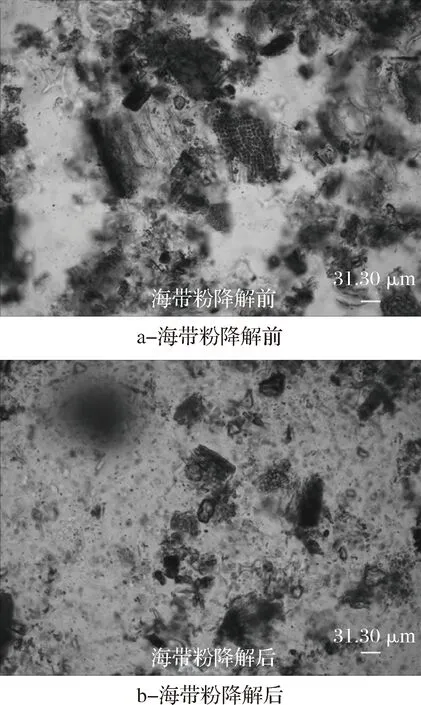
圖9 海帶粉降解前后的顯微照片Fig.9 Photomicrographs before and after degradation of kelp powder
從圖9中可以看出,海帶粉降解前,海帶的細胞結構較明顯,完整的細胞結合較緊密;而海帶粉降解后,其細胞結構明顯被降解,完整的細胞也被分散開。這可能是由于海帶粉經過YY7菌株產生的褐藻膠裂解酶、蛋白酶、淀粉酶等降解后,細胞間結合變得松散,細胞壁、細胞膜等組織結構也被部分降解。
3 結論
從皺紋盤鮑腸道中,以褐藻酸鈉為唯一碳源,篩選出產褐藻膠裂解酶菌株12株,其中酶活力最高的是YY7菌株,通過16S rRNA初步鑒定YY7菌株與弧菌屬Vibrioalgivorus的SA2(T)菌株的相似性最高,為99.72%;通過培養基鑒定發現,YY7菌株不具有溶血性,同時還產蛋白酶和淀粉酶。根據YY7菌株的生長曲線和產酶曲線可知,YY7菌株的生長速度快,其褐藻膠裂解酶活力在20 h可達到最高54.31 U/mL,最適溫度為35 ℃,最適pH 7.5,Na+、Mg2+、K+、Ca2+對褐藻膠裂解酶活力具有促進作用,Fe2+和Cu2+起抑制作用。最后利用YY7發酵液對海帶粉進行降解處理發現,海帶降解得率可達65%以上,降解效果佳。本研究結果表明,該菌株適合應用于海帶的加工利用,對于提高海帶利用率具有非常重要的作用。下一步還將繼續優化YY7菌株的發酵培養基及發酵條件,并進行放大培養,探索其在海帶精深加工的應用研究。
