茂蘭喀斯特森林自然保護區凋落葉分解動態
2019-06-03 07:14:46竇百強俞國松
水土保持通報 2019年2期
關鍵詞:差異
竇百強, 容 麗, 俞國松, 李 恒
(1.貴州師范大學 地理與環境科學學院, 貴州 貴陽 550001;2.中國科學院 地球化學研究所 環境地球化學國家重點實驗室, 貴州 貴陽 550081)
森林凋落物,也稱枯落物或有機碎屑,是指生態系統內由地上植物組分產生并歸還到地表,作為分解者的物質和能量來源,借以維持生態系統功能的所有有機物質的總稱[1-3]。其在維持土壤肥力、水土保持和促進森林生態系統養分循環等方面具有重要意義。凋落物分解是森林生態系統生物地球化學循環的重要組成部分,也是陸地生態系統初級生產力的重要物質來源,其分解速度的快慢也在很大程度上決定了森林生態系統的土壤肥力狀況以及林分生產力的高低[4-5]。西南喀斯特地區是中國生態系統較為脆弱的一個地區,喀斯特森林植被所處的生境條件大多比較惡劣,基巖裸露率高,土層淺薄,養分物質總量低,對植物生長要求較高,因而凋落葉的分解對喀斯特森林生態系統物質循環和能量轉換具有重要意義,對維持喀斯特森林生態系統的功能具有不可替代的作用。近年來,有關喀斯特森林凋落物的研究已有不少學者進行了報道,魏魯明等[6]、錢正敏等[7]對茂蘭喀斯特森林凋落物量動態進行了研究,俞國松等[8]、熊紅福等[9]研究了喀斯特地區不同演替群落的凋落物量、組成特征和月動態變化,朱雙燕等[10]、曾昭霞等[11]和盧曉強等[12]先后分析了喀斯特區原生林和次生林凋落物養分歸還特征,但針對喀斯特森林不同樹種凋落葉間養分循環特征的研究尚不多見,主要為對喀斯特區森林凋落葉生態化學計量特征的研究[13-14]。為此,本文擬通過對茂蘭喀斯特森林保護區內典型常綠落葉闊葉混交林進行為期18個月的凋落葉分解動態研究,探討喀斯特森林生態系統中凋落葉,特別是分別對常綠及落葉成分的失重率和干重殘留率動態變化以及分解速率進行探討,同時初步分析凋落葉養分釋放特征,旨在為闡明喀斯特森林生態系統凋落葉物質循環特征及生物地球化學過程提供科學依據。
1 研究區與樣地環境概況
研究區位于貴州省南部荔波縣境內的茂蘭喀斯特森林自然保護區內,處于貴州高原南部向廣西丘陵平原過渡的斜坡地帶,地理位置25°09′20″—25°20′50″N,107°52′10″—108°45′40″E。整個保護區呈東北—西南走向,區內地形以高原山地為主,為典型的喀斯特地貌,最高海拔1 078.6 m,最低海拔430 m,平均海拔500~800 m。年平均氣溫18.3 ℃,1月平均氣溫8.3 ℃,7月平均氣溫26.4 ℃,年平均積溫6 210.8 ℃,全年日照時數1 272.8 h,日照百分率29%;年降水量為1 752.5 mm,雨季集中在5—10月,年平均相對濕度為70%左右,屬中亞熱帶季風濕潤氣候。區內土壤以黑色石灰土為主,富鈣和富鹽基化,pH值為7.5~8.0,有機質和全氮含量特別豐富。土層淺薄且不連續,地面巖石裸露,持水量較低,地表水缺乏,臨時性干旱頻繁[7]。
試驗樣地位于茂蘭保護區西北部拉橋緩沖區,為典型喀斯特常綠落葉闊葉混交林,樣地內土層淺薄,生境類型主要為土面、石面、石溝和石縫,坡體下部受少量人為干擾。坡腳主要分布有:天仙果(Ficuserecta)、巴東莢迷(Viburnumhenryi)、香葉樹(Linderacommunis)、長梗羅傘(Brassaiopsisglomerulatavar.longipedicellata)、小葉欒樹(Boniodendronminus)、光皮梾木(Swidawilsoniana)、滇青岡(Cyclobalanopsisglaucoides)、牛耳楓(Daphniphyllumcalycinum)等植物。圓果化香樹(Platycaryalongipes)、短萼海桐(Pittosporumbrevicalyx)、小果潤楠(Machilusmicrocarpa)、青檀(Pteroceltistatarinowii)、光葉海桐(Pittosporumglabratum)、裂果衛矛(Euonymusdielsianus)、絲栗栲(Castanopsisfargesii)、十大功勞(Mahoniafortunei)等植物分布在中坡。而坡頂主要分布的植物有:燈臺樹(Cornuscontroversa)、香葉樹(Linderacommunis)、滇青岡(Cyclobalanopsisglaucoides)、多脈青岡(Cyclobalanopsismultiervis)、絲栗栲(Castanopsisfargesii)、山黃皮(Randiacochinchinensis)、山礬(Symplocossumuntia)、粗葉木(Lasianthuschinensis)等。
2 研究方法
2.1 凋落物處理
凋落葉的野外分解工作采用分解袋法。將收集的凋落葉按常綠葉、落葉分別裝入孔徑為1 mm,規格為10 cm×20 cm的分解袋中,每個分解袋裝入10 g烘干至恒重的葉凋落物樣品。同時,為了盡量縮小由于分解袋的隔離作用及其形成的小環境限制土壤動物的分解作用,在分解袋正反兩面各剪出一個1 cm2的孔。在茂蘭喀斯特原生林內,下坡和上坡各設置3個樣地、中坡4個樣地,共10個樣地,樣地面積10 m×10 m。于2015年8月下旬將標記好的樣袋放置于樣地林下進行分解,每個樣地按常綠葉和落葉分別放置30個分解袋,即一個樣地放置60個分解袋,下坡和上坡各放置180個分解袋,中坡放置240個分解袋,總計600個分解袋。放置時貼近地表模擬凋落葉分解的自然狀況。在2015年9月至2017年2月的分解期內,每2個月取一次分解袋,每個樣地按常綠葉和落葉各取3袋,一個樣地每次取6袋,即下坡和上坡每次各取18袋,中坡每次取24袋,每次總計取60袋,至2017年2月底共回收9次。將取回的樣品中附著的雜物清除后,于80 ℃烘箱中烘干至恒重,測定剩余凋落葉質量及其百分比[15]。
2.2 凋落葉N,C元素測定
將不同坡位凋落葉樣品混合均勻進行烘干后,分別將落葉與常綠葉樣品磨細,過60目篩后,放置于廣口瓶中備測。采用元素分析儀(PE2400-Ⅱ)測定凋落葉N,C含量。
2.3 數據分析與處理
使用Excel 2016軟件對數據進行整理后,用SPSS 23.0軟件對凋落葉失重率和養分含量進行單因素方差分析(one-way ANOVA),對凋落葉干重殘留率進行回歸分析。
凋落葉失重率計算公式為:
R=(W1-W2)/W1×100%
式中:R——凋落物失重率;W1——初始重量;W2——分解后的重量。
凋落葉分解速率采用Olson指數衰減模型[16]計算:
y=Xt/X0=e-kt
式中:Xt——時間t時的干重殘留量;X0——凋落物的初始重量;Xt/X0——凋落物干重殘留率;k——凋落物的年分解系數或腐解系數。
試驗期間茂蘭喀斯特森林區的月平均降雨量、氣溫和濕度分布如圖1所示。

圖1 茂蘭喀斯特森林區2015-2017年平均氣候狀況
3 結果與分析
3.1 凋落葉失重率及季節動態變化
計算出凋落葉的失重率,取平均值制成折線圖(圖2)。由圖2可知,在2015年9月到2017年2月的分解期中,落葉樹種和常綠樹種凋落葉的分解速度均呈現“快—慢—快”的周期變化。凋落葉的失重率變化呈雙峰型,第1次峰值期出現在分解期的第2個月(2015年10月),落葉樹種和常綠樹種凋落葉失重率分別為16.8%和15.4%,上坡、中坡和下坡的凋落葉失重率分別達到11%,16.38%和20.83%。但隨后的6個月(2015年11月至2016年4月)凋落葉的分解速度相對減慢,到第8個月(2016年4月)時失重率最低,之后凋落葉分解速度開始再次加快。落葉樹種和常綠樹種凋落葉的第2次峰值期出現在分解期的第10個月(2016年6月),其失重率分別為12.57%和13.33%。上坡和下坡的第2次峰值期出現在分解期的第10個月(2016年6月),上坡和下坡的凋落葉失重率分別為12.65%和17.72%;而中坡的第2次峰值期出現在分解期的第12個月(2016年8月),失重率為13.9%。由此可見,凋落葉分解的高峰期出現在春夏兩季,而秋冬兩季分解速度緩慢。
方差分析的結果表明,落葉樹種和常綠樹種凋落葉失重率無顯著性差異(p=0.971>0.05),不同坡位凋落葉的失重率也無顯著性差異(p=0.225>0.05)。
由圖1可知,茂蘭喀斯特森林區氣候雨熱同期,夏季高溫多雨,而冬季低溫少雨。夏季6月—8月的平均降雨量、氣溫及濕度為全年最高,其中7月份的平均降雨量為269 mm,平均濕度達79%,8月平均氣溫最高為26.5 ℃。冬季12月至翌年2月的平均降雨量、氣溫及濕度為全年最低,其中1月份的平均降雨量僅11.7 mm,平均濕度為68%,平均氣溫最低為6.5 ℃。本研究中,凋落葉的失重率峰值期有兩個,分別為2015年10月和2016年6—8月,其月平均氣溫均在20 ℃以上,平均濕度在75%左右,溫暖潮濕的環境有利于凋落葉的快速分解。而凋落葉失重率的低谷期為11月至翌年4月,低溫少雨,分解較慢。這種氣候的季節差異使凋落葉的分解呈現出“快—慢—快”的周期變化,春夏季節凋落葉的分解速度加快,而秋冬季節凋落葉的分解速度減慢。

圖2 研究區凋落葉失重率動態變化
3.2 凋落葉干重殘留率動態變化
3.2.1 不同樹種凋落葉干重殘留率變化 如圖3所示,凋落葉干重殘留率隨時間的推移總體呈不斷下降的趨勢。凋落葉干重殘留率越低,表明其分解速度越快,而且不同坡位的凋落葉分解均有明顯的階段性。上坡凋落葉分解前期(前8個月)常綠樹種凋落葉分解速度比落葉樹種凋落葉分解速度快,分解中期(第8—14個月)落葉樹種凋落葉分解速度快于常綠樹種凋落葉分解速度,分解后期(第14—18個月)落葉樹種凋落葉分解速度趨于平緩,而常綠樹種凋落葉分解速度再次超過落葉樹種凋落葉分解速度,落葉樹種凋落葉和常綠樹種凋落葉最終干重殘留率分別為56%和51%。中坡和下坡凋落葉分解前期(前8個月)分解速度較快,分解中期(第8—14個月)分解速度趨緩,分解后期(第14—18個月)分解速度再次加快。中坡和下坡落葉樹種凋落葉最終干重殘留率分別為42%和27%,均低于常綠樹種凋落葉在中坡和下坡的最終干重殘留率(45%和33%),說明中坡和下坡的落葉樹種凋落葉分解速度比常綠樹種凋落葉分解速度快,且中坡和下坡凋落葉分解過程中落葉樹種凋落葉分解速度始終快于常綠樹種凋落葉分解速度。
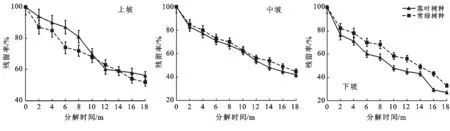
圖3 研究區不同樹種凋落葉殘留率動態變化
3.2.2 不同坡位凋落葉干重殘留率變化 由圖4可知,茂蘭喀斯特森林區凋落葉在經過18個月的分解期后,落葉樹種凋落葉分解后的最終干重殘留率由小到大依次為:下坡(27%)<中坡(42%)<上坡(56%);常綠樹種凋落葉分解后的最終干重殘留率由小到大依次為:下坡(33%)<中坡(45%)<上坡(51%)。而且在凋落葉分解過程中,不同坡位凋落葉殘留率大小始終為:上坡>中坡>下坡。由此可知,茂蘭喀斯特森林區同生活型樹種的凋落葉分解速度在不同坡位存在差異,分解速度為:下坡﹥中坡﹥上坡。此外,常綠樹種凋落葉分解速度在不同坡位的差異同落葉樹種凋落葉相比較小。
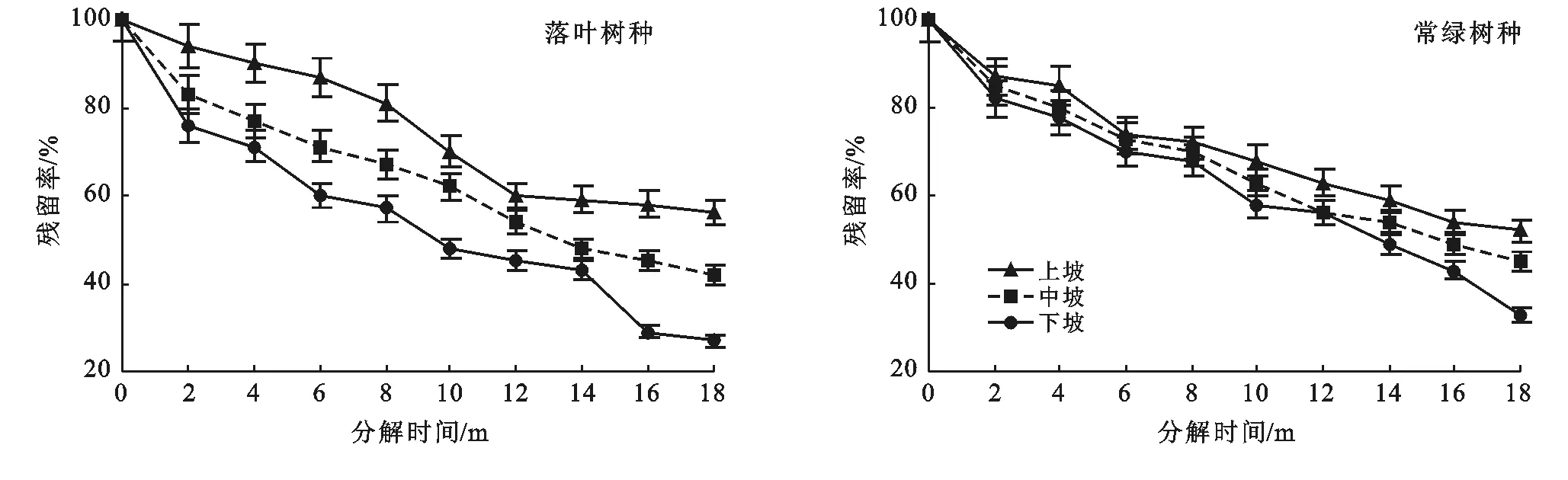
圖4 研究區不同坡位凋落葉殘留率動態變化
3.3 凋落葉分解時間預測
為比較不同類型凋落葉在不同坡位分解速率的差異,利用Olson指數衰減模型對凋落葉分解50%和95%所需時間進行估測(表1)。由表1可以看出,不同坡位落葉樹種與常綠樹種凋落葉分解模型的判定系數均很高,擬合效果較好(R2>0.9,p<0.01)。落葉樹種凋落葉和常綠樹種凋落葉的年分解速率分別為0.548,0.492。
落葉樹種凋落葉上、中、下坡的年分解系數分別為0.417,0.501,0.727,均大于常綠樹種凋落葉上坡(0.411)、中坡(0.455)和下坡(0.610)的年分解系數。而且不同坡位落葉樹種凋落葉分解50%和95%所需時間分別為0.95~1.66 a和4.13~7.19 a,均少于常綠樹種凋落葉分解50%(1.14~1.69 a)和95%(4.92~7.30 a)所需時間,所以落葉樹種凋落葉和常綠樹種凋落葉在不同坡位的分解速度均表現為:下坡>中坡>上坡,且落葉樹種凋落葉分解速度快于常綠樹種凋落葉。從Olson指數衰減模型計算得出的結論來看,與上述研究結果相一致,符合試驗實際情況。
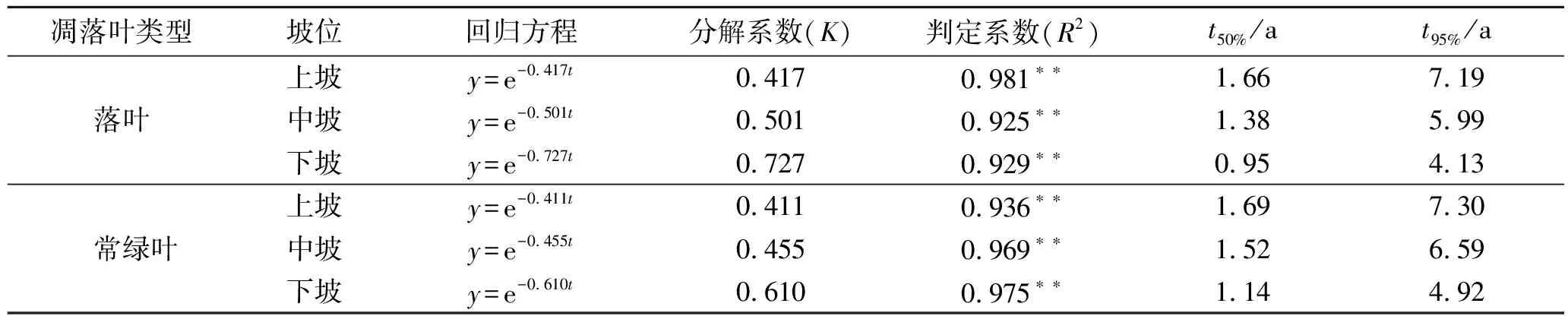
表1 凋落葉殘留率隨時間的指數回歸方程
注:*p<0.05,**p<0.01;t50%為分解50%所需時間,t95%為分解95%所需時間。
3.4 凋落葉養分動態變化
由圖5可知,落葉和常綠葉的初始N含量分別為1.28%和1.13%,而初始C含量分別為42.21%和45.54%。在分解過程中,落葉的N含量始終比常綠葉的N含量高,而落葉的C含量始終低于常綠葉。落葉和常綠葉的N含量前期均表現出富集現象,隨后分別在第14個月和第16個月N含量達到一個最高值后開始釋放,在試驗后期凋落葉N元素釋放階段,落葉N元素釋放速率大于常綠葉N元素釋放速率。而落葉和常綠葉的C含量波動性較大,無規律,但總體隨分解時間的推移而減少,經過18個月的分解后,落葉和常綠葉的最終C含量分別為38.12%和41.88%,落葉C元素釋放速率大于常綠葉C元素釋放速率。落葉和常綠葉的初始C/N比值分別為33.1,40.2,隨著分解時間增加,落葉和常綠葉的C/N比值均呈下降趨勢,其最終C/N比值分別為20.47,24.16。方差分析表明,落葉樹種凋落葉和常綠樹種凋落葉的N含量差異極顯著(p=0.001<0.01),C含量差異顯著(p=0.014<0.05),C/N比值差異極顯著(p=0.002<0.01)。
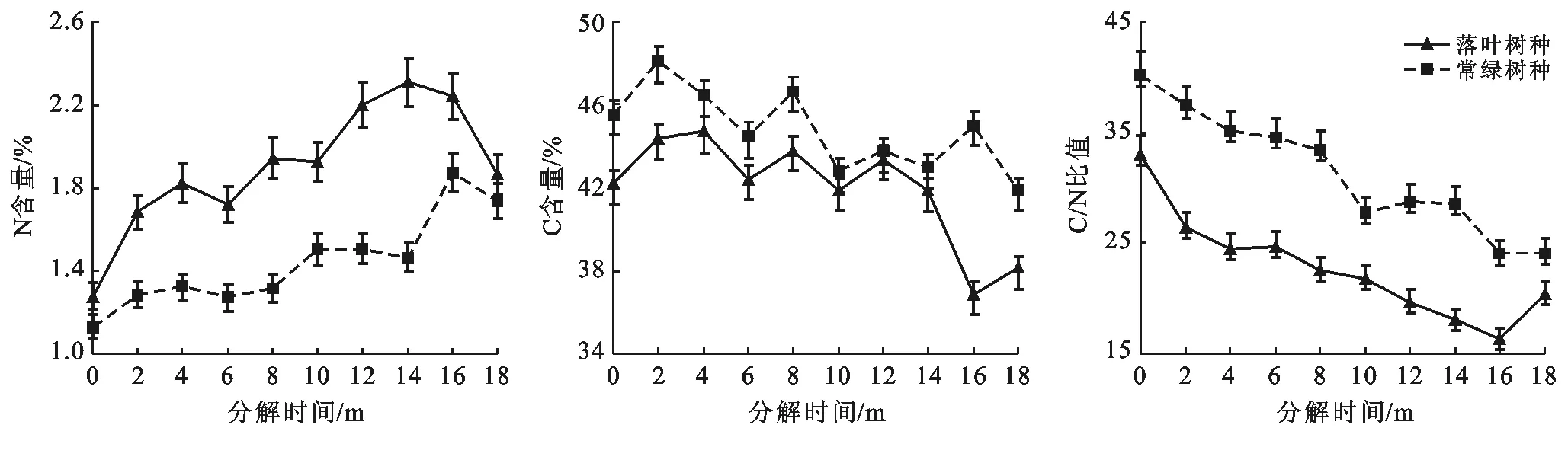
圖5 研究區凋落葉N,C含量及C/N比動態變化
4 討 論
4.1 不同樹種凋落葉分解的季節變化動態及差異
由以上結果可知,落葉樹種和常綠樹種凋落葉間的失重率差異并不顯著,且二者的分解均呈現出較明顯的季節性變化差異,落葉樹種和常綠樹種凋落葉分解的高峰期均出現在春夏兩季,而秋冬兩季分解速度相對緩慢。茂蘭喀斯特森林區氣候對凋落葉分解的影響主要體現在季節變化所引起的溫度和濕度差異。結合圖1,茂蘭喀斯特森林區5—9月高溫多雨,濕度較大,是凋落葉失重率在一年中的高峰期。春夏季由于氣溫升高、降水量增加改變了土壤動物的生境,提高了微生物活性,加快了有機質和凋落葉分解以及養分元素循環的進行[17]。而秋冬季節則由于氣溫下降限制了微生物的活性,凋落葉的分解速度減慢。同時,本研究發現,春夏季凋落葉分解率比秋冬季凋落葉分解率高6%~8%,與Moore等[18]研究得出的隨著氣溫升高,凋落物分解率會增加4%~7%的結論相近。本研究中,凋落葉的分解率呈現“快—慢—快”的周期變化,這可能是由于分解初期可溶性有機物的淋洗和易分解碳水化合物的分解,但到一定階段,難分解的纖維素和單寧等物質的積累,導致分解速度變慢,隨著時間推移,組織結構崩潰,分解速度再次加快[19]。
凋落葉殘留率越低,表明其分解速度越快。本研究發現,落葉樹種凋落葉最終干重殘留率低于常綠樹種凋落葉最終干重殘留率,落葉樹種凋落葉分解速度比常綠樹種凋落葉分解速度快。在凋落葉分解過程中,不僅受外界氣候因子的影響,也受凋落葉自身理化性質的作用,這是不同樹種凋落葉分解速率不同的關鍵原因[20]。一般的落葉樹種葉片為比較舒展、輕薄的紙質葉片,而常綠樹種的葉片通常為革質葉片,表面有一層蠟質保護膜,比落葉樹種的葉片要厚一些,所以微生物一般不易在很短的時間內對常綠樹種凋落葉進行分解,而且也不利于降雨淋溶和土壤動物的機械破壞,因此落葉樹種凋落葉分解速度比常綠樹種凋落葉分解速度快。
4.2 不同樹種凋落葉分解的養分動態及養分釋放速率
不同樹種凋落葉的養分組成不同,對其分解速率和養分釋放均有一定影響。在凋落物分解過程中,N是制約凋落物分解的重要因素之一[21]。一般認為,N含量高的凋落物其分解速率也會比較高,凋落物初始N含量與分解速率呈正相關關系[20]。本研究中,初始N含量:落葉>常綠葉,分解速率:落葉>常綠葉。落葉與常綠葉分解過程中的N元素釋放模式均為富集—釋放模式,N含量的動態變化呈先上升后下降的趨勢。這主要是因為凋落物在分解初期,微生物必須從外部獲取N源以滿足自身分解的需要,故分解初期N的絕對含量趨于升高[22]。落葉與常綠葉的初始C含量分別為42.21%和45.54%,較高的C含量是由于植物葉片需合成有機物質,從而維持其生長和代謝。凋落葉C含量隨分解時間的推進呈波動性變化,差異顯著,這可能是由凋落葉初始C含量和試驗樣本量差異造成的。C/N是凋落物分解的一個重要預測指標,本研究中,C/N比值隨凋落葉的分解呈不斷下降的趨勢,這是由于凋落葉分解到后期,難分解物質相對增加,分解速度變緩,C/N比就會逐漸降低[23]。有研究表明,凋落物中C含量及C/N比值越低分解失重及元素釋放就越高[5]。本研究中,落葉的C含量及C/N比值在分解過程中始終低于常綠葉,落葉元素釋放速率大于常綠葉元素釋放速率。不同樹種凋落葉中養分元素釋放速率的差異,與凋落葉中養分元素的初始含量和葉片結構特征有關。
4.3 不同樹種凋落葉分解時間預測
由于不同地區林分特征、生境條件及凋落物質量不同,即使是同一氣候帶,其凋落物分解速率也會存在明顯的差異。本研究中,茂蘭喀斯特森林區不同坡位落葉樹種與常綠樹種凋落葉分解95%所需時間分別為4.13~7.19 a和4.92~7.30 a,高于中亞熱帶東部武夷山常綠闊葉林(3.01 a)[19]和浙江天童常綠闊葉林(2~4.5 a)[24],略高于神農架常綠落葉闊葉混交林(4.54 a)[25]。水熱環境等因素的差異是導致上述區域森林凋落葉分解速率差異的主要原因[26]。而與同區域的喀斯特森林相比,低于黔中喀斯特次生林(3.1~10 a)[27]和貴州普定喀斯特人工林(5~10 a)[28],說明凋落葉分解速率也因不同林分而異。由此可知,茂蘭喀斯特森林區凋落葉分解速率比中亞熱帶東部常綠闊葉林和常綠落葉闊葉混交林低,但高于同區域喀斯特次生林與人工林。
5 結 論
由于不同季節水熱條件的差異,茂蘭喀斯特森林區不同樹種凋落葉分解率均呈現較明顯的季節性變化差異,表現為春夏季凋落葉分解率大于秋冬季凋落葉分解率,分解趨勢表現出“快—慢—快”的節奏。此外,受不同凋落葉初始質量和葉片結構特征差異的影響,落葉樹種凋落葉的分解速率和養分釋放速率均大于常綠樹種凋落葉,因此,落葉樹種凋落葉對維持林地土壤肥力,促進茂蘭喀斯特森林生態系統物質循環具有積極作用。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
