Mfge8基因敲除純合子小鼠的定向繁殖和鑒定及其與自身免疫疾病相關(guān)性分析
2019-06-05 01:50:46楊文強(qiáng)崔天盆
中國比較醫(yī)學(xué)雜志 2019年5期
楊文強(qiáng),黃 偉,馬 威,崔天盆
(武漢市第一醫(yī)院中心實驗室,武漢 430022)
乳脂球表皮生長因子8(milk fat globule EGF factor Ⅷ,MFG-E8)是一種乳汁脂肪小球表面的親脂性糖蛋白,最初在小鼠乳腺上皮細(xì)胞被發(fā)現(xiàn)[1],生理情況下在其他細(xì)胞也有表達(dá),如角質(zhì)形成細(xì)胞、脾細(xì)胞、單核細(xì)胞、腹腔巨噬細(xì)胞、樹突狀細(xì)胞、神經(jīng)膠質(zhì)細(xì)胞和抗原提呈細(xì)胞[2]等,未成熟的樹突狀細(xì)胞和未分化的巨噬細(xì)胞表達(dá)較多的MFG-E8,主要以外泌體的方式分泌,隨著細(xì)胞成熟,MFG-E8的表達(dá)降低。MFG-E8主要在吞噬細(xì)胞與凋亡細(xì)胞之間起橋連作用,增強(qiáng)了凋亡細(xì)胞的吞噬清除[3],維持機(jī)體的內(nèi)環(huán)境穩(wěn)態(tài)。Uchiyama等[4]的研究證實MFG-E8可加速糖尿病傷口愈合,促進(jìn)血管生成,凋亡細(xì)胞附近M2吞噬細(xì)胞的浸潤,進(jìn)而抑制傷口處炎性細(xì)胞因子;在結(jié)腸炎動物模型中,接受了重組人MFG-E8治療的小鼠中性粒細(xì)胞和凋亡細(xì)胞浸潤明顯減少,細(xì)胞因子和趨化因子的表達(dá)也顯著降低[5];MFG-E8通過控制間充質(zhì)干細(xì)胞中VGEF和ET-1的表達(dá),增強(qiáng)巨噬細(xì)胞向M2型極化,加速血管生成,從而促進(jìn)黑色素瘤的生成[6]。MFG-E8可能作為一種新型生物標(biāo)志物來治療人類類風(fēng)濕性關(guān)節(jié)炎(RA),因為Albus等人[7]發(fā)現(xiàn)Mfge8基因敲除小鼠關(guān)節(jié)中中性粒細(xì)胞數(shù)量增加,并伴隨有破骨細(xì)胞的活化和成骨細(xì)胞的破壞,導(dǎo)致系統(tǒng)性骨質(zhì)流失;MFG-E8的缺失也會導(dǎo)致小鼠的脾腫大,形成無數(shù)的生發(fā)中心,由于在生發(fā)中心中清除凋亡B細(xì)胞的能力下降,導(dǎo)致腎小球腎炎自身抗體的產(chǎn)生,進(jìn)一步發(fā)展為自身免疫病[8];在系統(tǒng)性紅斑狼瘡(SLE)動物模型中,MFG-E8可以下調(diào)中性粒細(xì)胞表面CXCR2的表達(dá),同時使得凋亡的中性粒細(xì)胞被吞噬,降低SLE小鼠體內(nèi)早期炎癥反應(yīng)[9]。本課題組自主構(gòu)建以C57BL/6小鼠為背景的Mfge8基因敲除小鼠并穩(wěn)定繁殖鑒定,得到的純合子小鼠旨在自身免疫性疾病中作進(jìn)一步研究。
1 材料和方法
1.1 實驗動物
Mfge8基因敲除雜合子小鼠在南京模式動物研究院構(gòu)建[SCXK(蘇)2015-0001],背景鼠為C57BL/6小鼠,其中雄性11只,雌性21只,體重16~20 g,8周齡。穩(wěn)定繁殖后在武漢市第一醫(yī)院SPF級動物實驗室[SYXK(鄂)2014-0030]內(nèi)擴(kuò)群使用,取純合子和野生型小鼠各20只,雌雄各半,其中12周齡純合子和野生型各8只,體重20~24 g,36周齡純合子和野生型各12只,體重24~28 g。本研究經(jīng)由武漢市第一醫(yī)院實驗動物福利倫理委員會批準(zhǔn)(批準(zhǔn)號:2017010),嚴(yán)格按實驗動物使用的3R原則給予人道的關(guān)懷。
1.2 主要試劑與儀器
Simgen動物組織DNA試劑盒、蛋白酶K(杭州新景生物試劑開發(fā)有限公司),2× Quick Taq? HS DyeMix(上海東洋紡生物科技有限公司),6× Loading buffer、DNA Marker(北京索萊寶科技有限公司),瓊脂糖(美國Bio-West公司),GoldView核酸染料(上海賽百盛基因技術(shù)有限公司),羅氏TUNEL檢測試劑盒(上海羅氏診斷產(chǎn)品有限公司),抗核抗體IgG檢測試劑盒、抗內(nèi)皮細(xì)胞抗體IgG檢測試劑盒(歐蒙杭州醫(yī)學(xué)實驗診斷有限公司)。臥式滅菌器YX-600 W(上海三申器械有限公司),恒溫混勻儀、低溫高速離心機(jī)(德國Eppendorf公司),PCR儀(西安天隆科技有限公司),電泳儀(北京市六一儀器廠),化學(xué)發(fā)光成像儀(美國Bio-Rad公司),熒光顯微鏡BX51(日本Olympus公司)。
1.3 實驗方法
1.3.1 小鼠的飼養(yǎng)與繁殖
按照SPF級動物飼養(yǎng)標(biāo)準(zhǔn)進(jìn)行飼養(yǎng),屏障環(huán)境內(nèi)溫度控制在20~22℃,濕度40%~70%,12 h/12 h晝夜交替,小鼠飼喂已輻照消毒的繁殖飼料(北京華阜康公司)和去離子水,實行自由采食和飲水,小鼠墊料和籠盒均經(jīng)過高溫高壓滅菌處理,每周更換籠盒墊料兩次。因引進(jìn)小鼠有限,繁殖初期按1只雄鼠與1只雌鼠進(jìn)行合籠,小鼠性成熟期為8周左右,母鼠妊娠期21 d左右,子代小鼠10 d左右剪尾鑒定和剪腳趾編號,仔鼠21 d斷乳分籠,繁殖出F2代子鼠以后挑選合適基因型按1只雄鼠與2只雌鼠進(jìn)行合籠,逐漸擴(kuò)大繁殖。
1.3.2 小鼠的基因型鑒定
由于所構(gòu)建的小鼠均為雜合子Mfge8基因敲除小鼠,繁殖后子代可能出現(xiàn)野生型(Mfge8+/+)、雜合子(Mfge8+/-)和純合子(Mfge8-/-)3種表型,故需對子代進(jìn)行基因型鑒定。
(1)抽提鼠尾基因組DNA:剪取小鼠尾尖0.5~1.0 cm,放入1.5 mL EP管中,參照Simgen動物組織DNA試劑盒說明書抽提DNA,TE溶解后測OD值,于4℃保存。
(2)PCR擴(kuò)增反應(yīng)及瓊脂糖凝膠電泳進(jìn)行基因型鑒定:①引物設(shè)計:針對Mfge8基因敲除小鼠的基因型設(shè)計引物:上游引物:P1 5’-GTGGGCAAGTG CATCTGAGTAC-3’,下游引物:P2 5’-GAGCGATCC TATCTCAAAACCAA-3’,用于檢測Mfge8基因敲除小鼠中包含LoxP插入位點的等位基因片段(擴(kuò)增大小 620 bp)。針對野生型小鼠設(shè)計引物:上游引物:P3 5’-TTGCCAACAGGCTTGATGGATAT-3’,下游引物:P4 5’-GACAGTACGGAACAGCGAAGGTA-3’,用于檢測野生型小鼠的Mfge8等位基因的片段(擴(kuò)增大小453 bp)引物由上海生工生物工程股份有限公司合成,為干粉狀。②PCR擴(kuò)增:10 μL反應(yīng)體系:用微量移液槍依次加入DNA模板1 μL、PCR Master Mix 5 μL、上游引物1 μL、下游引物1 μL和ddH2O 2 μL,充分混勻后將PCR管置于PCR反應(yīng)儀上進(jìn)行擴(kuò)增,擴(kuò)增條件:預(yù)變性94℃、3 min;變性94℃、30 s,退火56℃、40 s,延伸68℃、40 s,循環(huán)35次;末次循環(huán)后延伸68℃、10 min。4℃保存。③瓊脂糖凝膠電泳:樣品孔分別取PCR擴(kuò)增產(chǎn)物7 μL與6× Loading buffer 1 μL混勻,對照孔加Marker 5 μL作對照,在以GoldView為分子量標(biāo)記物的1.5%瓊脂糖凝膠中以120 V由負(fù)極向正極電泳30 min后,于凝膠成像中觀察拍照。

圖1 子代1日齡仔鼠Figure 1 One-day-old progeny mice
1.3.3 利用TUNEL法檢測小鼠肺組織中凋亡細(xì)胞
取12周齡Mfge8-/-和野生型小鼠左肺組織,放置于4%多聚甲醛溶液中固定,石蠟包埋切片,肺組織經(jīng)過脫蠟和抗原修復(fù)后,用羅氏TUNEL試劑盒顯色,在熒光顯微鏡下觀察拍照。
1.3.4 小鼠血清中抗核抗體(ANA)檢測
抗核抗體是一組具有多種細(xì)胞核成分的自身抗體,由于自身免疫疾病譜型范圍廣和自身抗體呈多樣性,ANA雖不能作為自身免疫疾病的診斷性指標(biāo),但對自身免疫病有一定的初篩作用[10]。本研究將36周齡的Mfge8-/-和野生型小鼠血清按1∶10稀釋后分別與猴肝組織作為基質(zhì)的生物薄片溫浴,洗滌后再與異硫氰酸熒光素標(biāo)記的羊抗小鼠IgG溫浴,再洗滌后在熒光顯微鏡下觀察結(jié)果。
1.3.5 小鼠血清中抗內(nèi)皮細(xì)胞抗體(AECA)檢測
抗內(nèi)皮細(xì)胞抗體最初在系統(tǒng)性紅斑狼瘡、類風(fēng)濕性關(guān)節(jié)炎等自身免疫疾病患者血清中獲得,其出現(xiàn)在多種自身免疫性疾病中,尤其是與血管炎相關(guān)的疾病,在系統(tǒng)性紅斑狼瘡的血管炎中表現(xiàn)尤為突出[11]。本研究將36周齡的Mfge8-/-和野生型小鼠血清按1∶10稀釋后分別與人臍靜脈內(nèi)皮細(xì)胞(HUVEC)作為基質(zhì)的生物薄片溫浴,洗滌后再與異硫氰酸熒光素標(biāo)記的羊抗小鼠IgG溫浴,再洗滌后熒光顯微鏡下觀察結(jié)果。
1.4 統(tǒng)計學(xué)方法
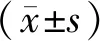
2 結(jié)果
2.1 小鼠繁殖情況
初次生產(chǎn)的母鼠會出現(xiàn)食仔現(xiàn)象,每胎平均產(chǎn)仔4~13只,如圖1所示,成活率大于90%。
2.2 小鼠基因型鑒定結(jié)果
利用PCR擴(kuò)增小鼠基因組DNA中因敲除而插入的loxp位點基因及Mfge8基因片段,并利用瓊脂糖凝膠電泳檢測擴(kuò)增片段的長度,用以鑒定子代小鼠的基因型。結(jié)果如圖2所示,擴(kuò)增片段1、2、5、6、9、10、12、13、14號為雜合子(Mfge8+/-);3號為野生型(Mfge8+/+);4、7、8、11號為純合子(Mfge8-/-)。
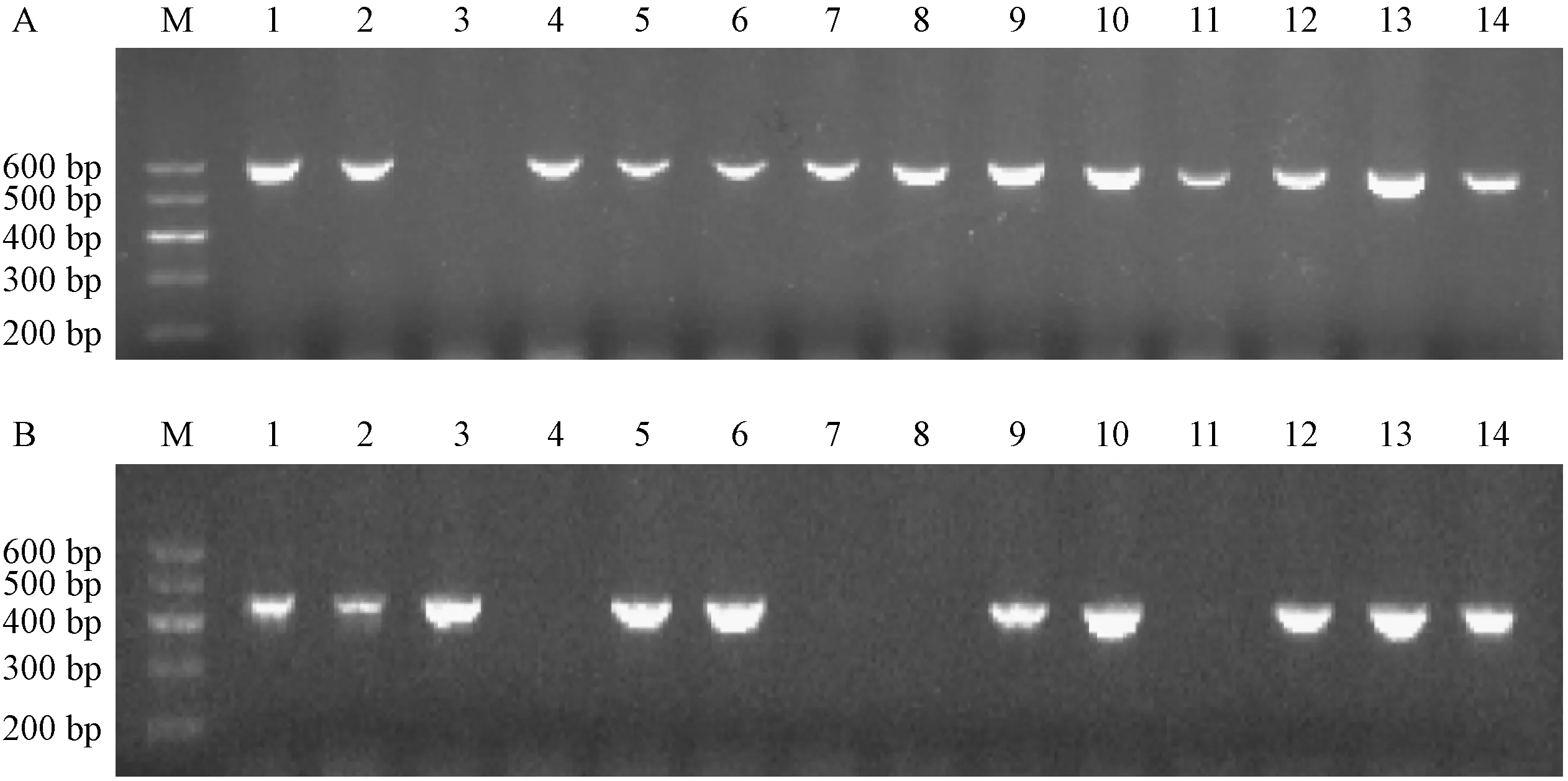
注:A:引物P1和P2擴(kuò)增620 bp片段;B:引物P3和P4擴(kuò)增453 bp片段。M為DNA Marker,1~14代表小鼠編號。圖2 PCR及瓊脂糖凝膠電泳鑒定子代小鼠基因型Note. A, Amplification of 620 bp fragment with primers P1 and P2. B, Amplification of 453 bp fragment with primers P3 and P4. M, DNA marker; 1-14, the mice number.Figure 2 Identification of progeny genotype by PCR and agarose gel electrophoresis
2.3 肺組織中凋亡細(xì)胞檢測結(jié)果
利用TUNEL法檢測12周齡WT和Mfge8-/-小鼠肺組織中凋亡細(xì)胞含量。在放大100倍視野下的觀察結(jié)果,凋亡細(xì)胞被染綠色顆粒的數(shù)量Mfge8-/-小鼠明顯比WT小鼠要多,因此12周齡Mfge8-/-小鼠肺組織中凋亡細(xì)胞數(shù)量明顯多于WT小鼠,如圖3所示。
2.4 小鼠血清中ANA檢測結(jié)果
36周齡WT和Mfge8-/-小鼠血清中IgG型抗核抗體(ANA)以猴肝Hep-2細(xì)胞為基質(zhì)利用間接免疫熒光法在100倍和400倍視野下觀察結(jié)果,Mfge8-/-小鼠在100倍和400倍兩個視野下細(xì)胞核被標(biāo)記的熒光強(qiáng)度明顯高于WT小鼠,因此36周齡Mfge8-/-小鼠血清中ANA抗體呈現(xiàn)陽性,而WT小鼠血清中ANA抗體呈現(xiàn)陰性,如圖4所示。
2.5 小鼠血清中AECA檢測結(jié)果
36周齡WT和Mfge8-/-小鼠血清中IgG型抗內(nèi)皮細(xì)胞抗體(AECA)以人臍靜脈內(nèi)皮細(xì)胞(HUVEC)為基質(zhì)利用間接免疫熒光法在400倍視野下觀察結(jié)果,Mfge8-/-小鼠內(nèi)皮細(xì)胞外周被標(biāo)記的熒光強(qiáng)度明顯高于WT小鼠,因此36周齡Mfge8-/-小鼠血清中AECA抗體呈現(xiàn)陽性,而WT小鼠血清中AECA抗體呈現(xiàn)陰性,如圖5所示。
3 討論
由于構(gòu)建的Mfge8基因敲除小鼠均為雜合子,所以其子代可能出現(xiàn)Mfge8+/+、Mfge8+/-和Mfge8-/-三種表型,我們采用PCR用兩組引物擴(kuò)增小鼠DNA,然后進(jìn)行瓊脂糖凝膠電泳,Mfge8-/-表型的小鼠擴(kuò)增產(chǎn)物只有一條帶,即620 bp,Mfge8+/+表型的小鼠也只有一條帶,即453 bp,而Mfge8+/-表型的小鼠同時會出現(xiàn)620 bp和453 bp兩條帶。
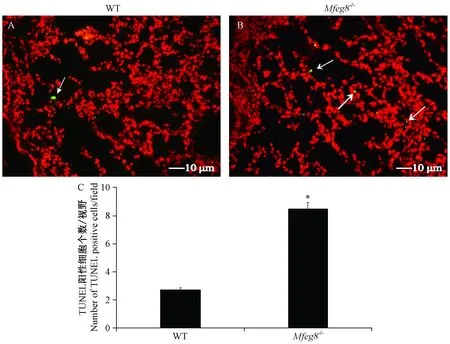
注:A:WT小鼠肺組織中凋亡細(xì)胞(× 100);B:Mfge8-/-小鼠肺組織中凋亡細(xì)胞(× 100)。箭頭所指為凋亡細(xì)胞。C:隨機(jī)視野下TUNEL陽性細(xì)胞個數(shù)統(tǒng)計(×100),與WT小鼠相比,*P<0.05。圖3 TUNEL染色檢測肺組織中凋亡細(xì)胞Note. A, Apoptotic cells in the lung tissue of a WT mouse (× 100). B, Apoptotic cells in the lung tissue of a Mfge8-/- mouse (× 100). Arrows indicate apoptotic cells. C, Average number of TUNEL positive cells in random view fields (×100). Compared with the WT mice,*P<0.05.Figure 3 Detection of apoptotic cells in lung tissues by TUNEL staining

注:A:WT小鼠血清中ANA(× 100);B:Mfge8-/-小鼠血清中ANA(× 100);C:WT小鼠血清中ANA(× 400);D:Mfge8-/-小鼠血清中ANA(× 400)。圖4 血清中ANA檢測結(jié)果Note. A, ANA in the serum of WT mice(× 100). B, ANA in the serum of Mfge8-/- mice(× 100). C, ANA in the serum of WT mice(× 400). D, ANA in the serum of MFGE8-/- mice(× 400).Figure 4 Analysis of ANA serum levels
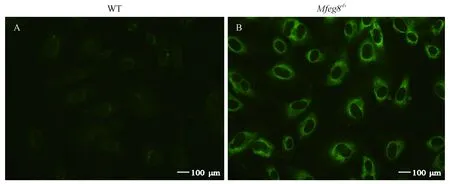
注:A:WT小鼠血清中AECA(× 400);B:Mfge8-/-小鼠血清中AECA(× 400)。圖5 血清中AECA檢測結(jié)果Note. A, AECA in the serum of WT mice. B, AECA in the serum of Mfge8-/- mice.Figure 5 Analysis of AECA serum levels
Hanayama等[3]研究表明,與野生型動物相比,不能分泌MFG-E8的動物巨噬細(xì)胞對凋亡細(xì)胞的吞噬能力減弱,由于缺乏足夠的清除凋亡細(xì)胞的能力,不分泌MFG-E8的成年動物更易引發(fā)自身免疫性疾病。抗核抗體(ANA)的產(chǎn)生細(xì)胞凋亡有兩種不同的理論,一種理論認(rèn)為由細(xì)胞凋亡抑制引起;另一種理論認(rèn)為與細(xì)胞過度凋亡有關(guān),在凋亡過程中產(chǎn)生的凋亡小體是一種重要的自身抗體來源,推測細(xì)胞凋亡增加時,吞噬細(xì)胞不能完全清除凋亡小體,未被吞噬的凋亡小體膜破裂,細(xì)胞核物質(zhì)增多[12],MFG-E8在吞噬細(xì)胞與凋亡細(xì)胞中的橋梁作用,可能會推動這種進(jìn)程,進(jìn)而誘導(dǎo)Mfge8-/-小鼠機(jī)體產(chǎn)生ANA;正常細(xì)胞膜上的磷脂呈不對稱分布,中性磷脂分布在細(xì)胞膜的外層,磷脂酰絲氨酸(PS)分布在細(xì)胞膜的內(nèi)層,當(dāng)細(xì)胞凋亡時,磷脂的不對稱分布被破壞,PS從細(xì)胞膜內(nèi)側(cè)向外側(cè)轉(zhuǎn)移,PS的暴露是內(nèi)皮細(xì)胞凋亡的最早期特征[13],MFG-E8包含兩個不同的功能結(jié)構(gòu)域,一個是有兩個表皮生長因子樣結(jié)構(gòu)域的氨基端(N端),另一個是羧基端(C端)的兩個與凝血因子Ⅴ和Ⅷ結(jié)構(gòu)域同源的C樣結(jié)構(gòu)域,能結(jié)合細(xì)胞膜的陰離子磷脂雙層,其中第二個C樣結(jié)構(gòu)域含有一個氨基的置換,可以使MFG-E8與PS膜有更強(qiáng)的親和力[14],暴露PS的凋亡內(nèi)皮細(xì)胞在MFG-E8的作用下更易被吞噬細(xì)胞識別并吞噬,反之,MFG-E8的缺乏不能及時清除凋亡的內(nèi)皮細(xì)胞,AECA也會隨即升高。血清中抗核抗體和抗內(nèi)皮細(xì)胞抗體均顯著增加(圖4和圖5),抗核抗體(ANA)對自身免疫性疾病有重要診斷價值[15],其中ANA的靶抗原為細(xì)胞死亡后釋放的細(xì)胞核內(nèi)不同生化成分,包括核酸、細(xì)胞核蛋白和核糖體蛋白,此外,抗內(nèi)皮細(xì)胞抗體(AECA)在臨床上主要用炎性疾病的判斷標(biāo)準(zhǔn),最主要的就是各種血管炎[16],在系統(tǒng)性紅斑狼瘡的活動期患者的血清中也發(fā)現(xiàn)高滴度的AECA,但是是否作為自身抗體還有待進(jìn)一步研究,MFG-E8的缺失促進(jìn)了炎癥的發(fā)展,進(jìn)而產(chǎn)生了大量的AECA。
MFG-E8是一種多結(jié)構(gòu)域蛋白,能介導(dǎo)不同類型的細(xì)胞間相互作用,其中在介導(dǎo)精卵結(jié)合、附睪上皮細(xì)胞的維持[17]、腸上皮細(xì)胞的維持與修復(fù)[18]、促進(jìn)乳腺分支形態(tài)發(fā)生[19]、促進(jìn)血管形成[20]、促進(jìn)樹突狀細(xì)胞外泌體功能和增強(qiáng)吞噬清除凋亡細(xì)胞等功能顯著。系統(tǒng)性紅斑狼瘡患者體內(nèi),低水平的MFG-E8能增強(qiáng)吞噬作用,高水平的MFG-E8減弱吞噬作用并呈劑量依賴性,其原因可能是過量的MFG-E8分別與吞噬細(xì)胞和凋亡細(xì)胞結(jié)合,打破了兩種細(xì)胞間MFG-E8的橋梁關(guān)系,從而引起吞噬障礙,引發(fā)慢性自身免疫疾病[21];文獻(xiàn)報道女性SLE患者的內(nèi)含子6發(fā)生一個少見突變,導(dǎo)致內(nèi)含子6中的一個隱匿外顯子拼接成截斷的MFG-E8,其異常糖基化和唾液酸化產(chǎn)生C端截短的MFG-E8,導(dǎo)致SLE發(fā)生[22]。Mfge8基因敲除小鼠由于其生發(fā)中心內(nèi)TBMs及FDCs吞噬凋亡細(xì)胞能力障礙,凋亡細(xì)胞過量積累,T細(xì)胞及B細(xì)胞過度活化,解剖Mfge8基因敲除老年小鼠呈現(xiàn)脾腫大;病理學(xué)檢測發(fā)現(xiàn)脾生發(fā)中心及T淋巴細(xì)胞區(qū)域增大,且TBMs增大伴有大量凋亡細(xì)胞在胞內(nèi)未被降解,因此,Mfge8基因敲除脾生發(fā)中心內(nèi)TBMs不能有效清除凋亡B細(xì)胞;血清學(xué)檢測老年Mfge8基因敲除小鼠體內(nèi)自身抗體明顯升高,表現(xiàn)為SLE樣自身免疫性疾病[8,23]。
本文通過構(gòu)建Mfge8基因敲除小鼠,檢測分析自發(fā)誘導(dǎo)自身免疫性疾病相關(guān)重要指標(biāo),為模擬人SLE等其它自身免疫疾病的發(fā)生發(fā)展提供一個有效的動物模型,同時為進(jìn)一步研究MFG-E8在自身免疫疾病中的發(fā)病機(jī)制提供了可靠的工具基礎(chǔ)。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫(yī)科大學(xué)學(xué)報(2021年2期)2021-03-29 05:31:08
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
云南中醫(yī)學(xué)院學(xué)報(2014年3期)2014-07-31 18:57:34
