異養(yǎng)鞭毛蟲對銅綠微囊藻的生長影響
2019-06-12 03:40:46羅曉霞李長玲何雪怡陶德琴杜振雄
廣東海洋大學學報 2019年3期
關鍵詞:實驗
羅曉霞,李長玲,何雪怡,陶德琴,杜振雄
?
異養(yǎng)鞭毛蟲對銅綠微囊藻的生長影響
羅曉霞1,2,3,李長玲1,2,3,何雪怡1,陶德琴1,杜振雄1
(1.廣東海洋大學水產學院//2. 廣東省藻類養(yǎng)殖及應用工程技術研究中心,廣東 湛江,524088; 3.廣東海洋大學深圳研究院,廣東 深圳,518000)
【】研究異養(yǎng)鞭毛蟲(sp.)在不同密度銅綠微囊藻()中的生長狀況,探討其對銅綠微囊藻的攝食及抑制作用。【】實驗共設6個銅綠微囊藻密度,分別是0、250′104、500′104、750′104、1 000′104和2 000′104cells/mL,研究異養(yǎng)鞭毛蟲與微囊藻的種群數量變化。【】異養(yǎng)鞭毛蟲主要行異養(yǎng)生活。在銅綠微囊藻密度為250′104~2 000′104cells/mL時,異養(yǎng)鞭毛蟲對銅綠微囊藻的抑制率為98.00%~99.69%。單個鞭毛蟲對微囊藻的攝食率為13~49 cells/d。異養(yǎng)鞭毛蟲的密度和生長率隨銅綠微囊藻密度的升高而增加,在銅綠微囊藻密度為500′104~1 000′104cells/mL時,異養(yǎng)鞭毛蟲密度為21′104~24′104cells/mL、生長率為0.6~0.74 d-1。但是,銅綠微囊藻密度過高也不利于異養(yǎng)鞭毛蟲的種群增長,在2 000′104cells/mL密度時,異養(yǎng)鞭毛蟲的生長速率顯著低于其它密度組。異養(yǎng)鞭毛蟲能有效抑制銅綠微囊藻種群增長,其生長的最適微囊藻密度為500′104~1 000′104cells/mL。
銅綠微囊藻;異養(yǎng)鞭毛蟲;生長率;抑制率;攝食率
目前,我國許多大型湖泊如太湖、巢湖、滇池等頻繁暴發(fā)藍藻水華。藍藻水華不僅使湖泊生態(tài)系統(tǒng)的服務功能喪失,降低生物多樣性,破壞生態(tài)系統(tǒng)的平衡,而且還會釋放出對水生動物、水禽、家畜和人類等有毒害作用的藻毒素[1-2]。其中,微囊藻水華營養(yǎng)價值低[3],具有毒素[4]及群體的結構[5-6],不利于浮游動物的生長[7],使浮游植物和浮游動物之間的傳遞效率下降,藍藻水華越趨嚴重,這也是藍藻水華難以調控的主要原因之一。
富營養(yǎng)化水體中,微食物網被認為是連接藍藻水華——浮游動物的中間環(huán)節(jié)[8-11]。異養(yǎng)鞭毛蟲(Heterotrophic nano-flagellates, HNF)隸屬原生生物界、原生動物亞界、肉鞭門、鞭毛亞門,是微食物網中重要組成部分[12]。已有研究報道異養(yǎng)鞭毛蟲可以吞噬有機碎屑、細菌、綠藻、硅藻及藍藻[13-15]。鞭毛蟲通過吞噬微囊藻,減少微囊藻生物量,進而促進自身種群數量增長[16]。張雪等[17]提出鞭毛蟲sp.具有降解微囊藻藻毒素的作用。有研究[18-19]表明某些異養(yǎng)鞭毛蟲具有營養(yǎng)提升的作用,能重新合成長鏈多不飽合脂肪酸LCn-3PUFAs和甾醇。Luo等[9]認為在富營養(yǎng)化水體中,鞭毛蟲在藍藻水華→浮游動物碳流途徑中起關鍵營養(yǎng)提升作用,從而促進浮游動物的種群增長。因此,異養(yǎng)鞭毛蟲控制有害藻類種群數量的作用不可忽視,可通過主動利用異養(yǎng)鞭毛蟲控制藍藻水華,實現水體的生物調控。
不同異養(yǎng)鞭毛蟲具有不同吞噬能力。筆者在前期實驗中發(fā)現異養(yǎng)鞭毛蟲(sp.)在微囊藻的腐爛過程中大量增殖,且能有效抑制微囊藻的種群增長[9]。Cui等[20]研究了溫度、光照及擾動對鞭毛蟲攝食銅綠微囊藻()的影響,但是對于不同微囊藻濃度下,銅綠微囊藻與鞭毛蟲交互作用下的種群動態(tài)變化仍然缺乏研究數據。本研究把該種異養(yǎng)鞭毛蟲單種分離,研究異養(yǎng)鞭毛蟲在不同銅綠微囊藻密度下種群增長情況,同時研究其對微囊藻的攝食及抑制作用。研究結果有助于揭示鞭毛蟲參與自然界微囊藻水華調控的作用機制,同時探索大量培養(yǎng)鞭毛蟲的條件,為其在生物控藻上的應用提供理論依據。
1 材料與方法
1.1 實驗動物的采集與處理
異養(yǎng)鞭毛蟲(sp.)培養(yǎng)于1 000 mL錐形瓶,溫度20 ~ 25 ℃、照度1 300 ~ 1 700 lx,所投餌料為銅綠微囊藻()。
1.2 藻種及培養(yǎng)基
銅綠微囊藻()藻種(FACHB-905,產毒種)購于武漢水生生物研究所,采用BG11培養(yǎng)液培養(yǎng)。培養(yǎng)液用超純水(Diect-Q3,Milllipore)配制,經高壓滅菌后供培藻用。選處于指數生長期的藻進行實驗。
1.3 培養(yǎng)實驗
實驗培養(yǎng)容器是250 mL錐形瓶。銅綠微囊藻密度設置0、250×104、500×104、750×104、1 000×104和2 000×104cells/mL等6個梯度,每個梯度設3個平行。分別在每個錐形瓶里加入對應濃度的180 mL藻液,再加入20 mL異養(yǎng)鞭毛蟲,使每個實驗瓶中鞭毛蟲密度達5×104cells/mL[21-22];同時,每個微囊藻密度均設置2個對照組,設置相同密度銅綠微囊藻180 mL,添加20 mL BG11培養(yǎng)液,不添加異養(yǎng)鞭毛蟲。實驗在光照培養(yǎng)箱(MGC-250,上海一恒)進行。實驗培養(yǎng)條件:溫度為(25±1)℃,光照強度為1 500 lx,光照周期為L∶D=12∶12。分別在實驗1 d、2 d、3 d、4 d、6 d時采樣,每個錐形瓶取10 mL,加魯哥氏液固定后,用血球計數板計數,并記錄好異養(yǎng)鞭毛蟲和銅綠微囊藻數量。
1.4 計算方法
異養(yǎng)鞭毛蟲的生長率()計算公式如下:
(lnN- ln0)/, (1)
式中,0表示實驗開始時異養(yǎng)鞭毛蟲的密度(cells/ mL),N表示實驗結束時異養(yǎng)鞭毛蟲的密度(cells/ mL),表示實驗的持續(xù)時間(d)。
異養(yǎng)鞭毛蟲對銅綠微囊藻種群數量的抑制率(Inhibition Ratio,IR):
IR = (1 –M/c)×100%, (2)
式中,M表示實驗結束時處理組銅綠微囊藻的密度(cells/ mL),c表示實驗結束時對照組銅綠微囊藻的密度(cells/ mL),表示實驗的持續(xù)時間(d)。
異養(yǎng)鞭毛蟲對銅綠微囊藻的攝食率(Ingestion rate,)[23]:
(0–M)/{ [( N–0)/(lnN–ln0)]×}, (3)
式中,0表示實驗開始時處理組銅綠微囊藻的密度(cells/mL),M表示實驗結束時處理組銅綠微囊藻的密度(cells/mL),0表示實驗開始時異養(yǎng)鞭毛蟲的密度(cells/mL),N表示實驗結束時異養(yǎng)鞭毛蟲的密度(cells/mL),表示實驗的持續(xù)時間(d)。
1.5 數據處理方法
數據用SPSS 17.0 進行單因素方差分析, 原始數據符合方差齊性, 運用Duncan’s HSD (<0.05)方法。
2 結果
2.1 不同銅綠微囊藻密度條件下異養(yǎng)鞭毛蟲及銅綠微囊藻數量變化趨勢
在實驗期間(1~6 d),除了0密度對照組外,其它對照組的銅綠微囊藻密度均急劇上升,其數量比初始密度增長2 ~ 3.8倍(圖1)。處理組中,隨著異養(yǎng)鞭毛蟲數量的增長,微囊藻的數量急劇下降,表明異養(yǎng)鞭毛蟲能顯著抑制銅綠微囊藻數量的增長;然而,隨著微囊藻數量的減少,異養(yǎng)鞭毛蟲的數量也持續(xù)減少。0密度處理組異養(yǎng)鞭毛蟲數量僅有輕微增長,隨后快速下降。
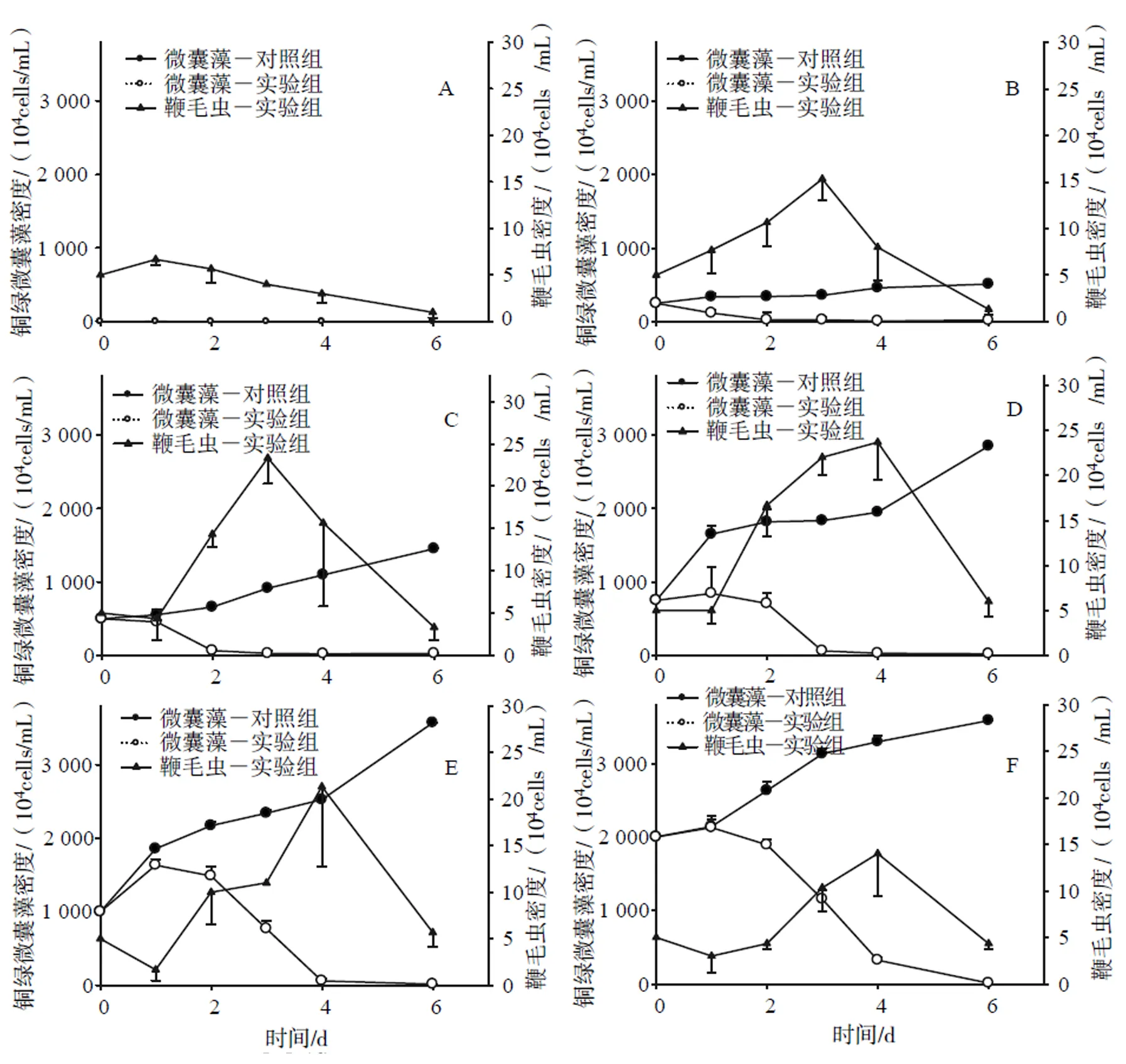
(A) 0 cells/mL;(B) 250×104 cells/mL;(C) 500×104 cells/mL; (D) 750×104 cells/mL; (E)1 000×104 cells/mL; (F) 2 000×104 cells/mL
2.2 不同銅綠微囊藻密度下異養(yǎng)鞭毛蟲的生長
不同微囊藻密度對異養(yǎng)鞭毛蟲數量有顯著影響(< 0.05)(圖2)。各密度組異養(yǎng)鞭毛蟲種群數量均呈先增加后減少的趨勢。當微囊藻密度為0時,異養(yǎng)鞭毛蟲僅在實驗1 d數量有輕微增長,隨后在2 d開始下降;其它密度組異養(yǎng)鞭毛蟲數量在1 ~ 4 d均有明顯增長。實驗1 d,0 ~ 750×104cells/mL密度組異養(yǎng)鞭毛蟲數量差異不顯著,但是顯著高于高密度組(1 000×104cells/mL、2 000×104cells/mL)。2 d、3 d時,500×104cells/mL、750×104cells/mL密度組的異養(yǎng)鞭毛蟲數量顯著高于0cells/mL密度組及高密度組(1 000×104cells/mL、2 000×104cells/mL)。4 d、6 d時,密度為750×104cells/mL、1 000×104cells/mL組的異養(yǎng)鞭毛蟲數量顯著高于低密度組(0、250×104cells/mL),但是500×104~ 2 000×104cells/mL密度組間鞭毛蟲數量無顯著差異。因此,銅綠微囊藻密度較低時,異養(yǎng)鞭毛蟲數量增長較快,但是當銅綠微囊藻數量過低不足以支持異養(yǎng)鞭毛蟲的生長時,異養(yǎng)鞭毛蟲的數量也隨之下降。高密度的銅綠微囊藻抑制異養(yǎng)鞭毛蟲的增長速度,但是高密度的銅綠微囊藻能支持異養(yǎng)鞭毛蟲數量的持續(xù)增長,其中異養(yǎng)鞭毛蟲數量的高峰值出現在密度組500×104~ 1 000×104cells/mL,異養(yǎng)鞭毛蟲數量達21×104~24×104cells/mL。然而,密度最高的2 000×104cells/mL組的異養(yǎng)鞭毛蟲數量增長較慢,且鞭毛蟲最高密度低于500×104~ 1 000×104cells/mL密度組,說明銅綠微囊藻密度過高也不利于異養(yǎng)鞭毛蟲生長。因此,異養(yǎng)鞭毛蟲生長的最適微囊藻密度范圍為500×104~ 1 000×104cells/mL。
銅綠微囊藻密度對異養(yǎng)鞭毛蟲的生長率有顯著影響(< 0.05)(圖3)。實驗1 d,低密度組0~ 250×104cells/mL的異養(yǎng)鞭毛蟲的生長率為正增長,而密度較高的500×104~ 2 000×104cells/mL組均為負增長,其中,高密度組(1 000×104cells/mL、2 000×104cells/mL)的生長率顯著小于低密度組的生長率(0、250×104cells/mL),說明高密度銅綠微囊藻對異養(yǎng)鞭毛蟲生長有一定抑制作用。2 d時,250×104~ 1 000×104cells/mL密度組的生長率顯著高于最低的0密度及最高的2 000×104cells/mL密度;3 d時,密度為500×104cells/mL、750×104cells/mL時,異養(yǎng)鞭毛蟲生長率顯著高于其它組,其中0密度組的異養(yǎng)鞭毛蟲生長率出現負增長;4 d時,750×104cells/mL、1 000×104cells/mL密度組生長率顯著高于密度較低的0~250×104cells/mL。6 d時,0、250×104cells/mL、500×104cells/mL、2 000×104cells/mL密度組的生長率均出現負增長。因此,銅綠微囊藻密度較低時,異養(yǎng)鞭毛蟲種群生長快,較高密度的銅綠微囊藻開始抑制異養(yǎng)鞭毛蟲種群的增長,但是異養(yǎng)鞭毛蟲在高密度銅綠微囊藻中獲得更加持久的種群生長率,其中,異養(yǎng)鞭毛蟲在銅綠微囊藻密度為500×104~1 000×104cells/mL時維持了較高生長率,達0.65 ~ 0.74 d-1。
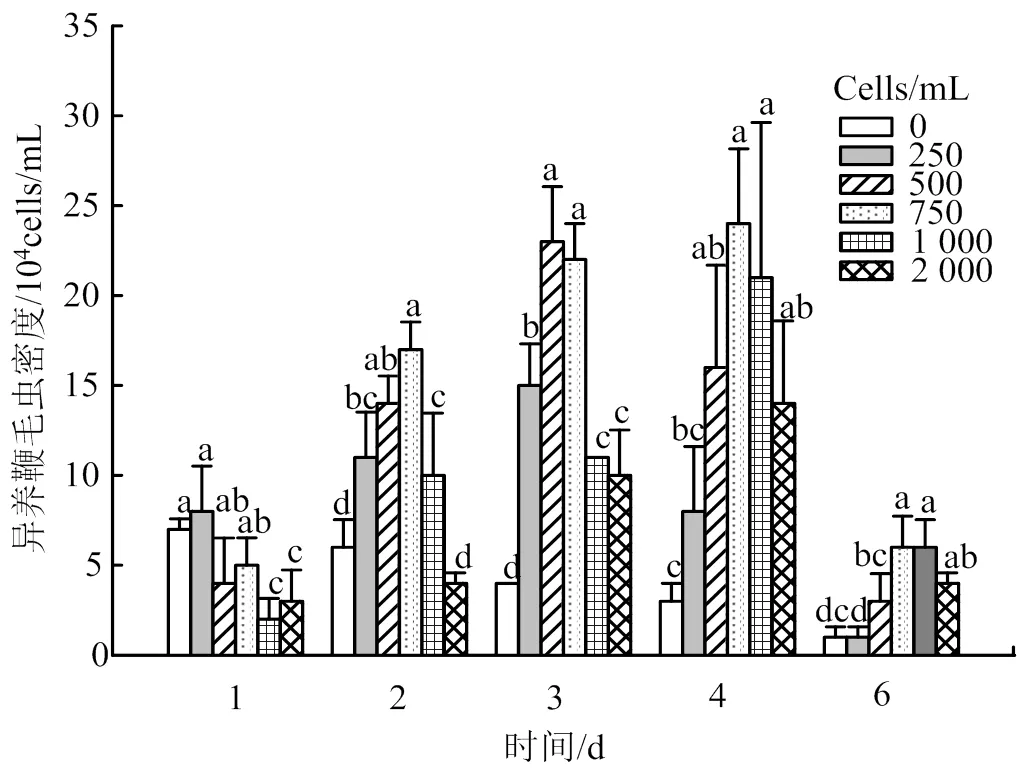
上標字母不同代表差異顯著(Duncan’s multiple range tests, P <0.05)

上標字母不同代表差異顯著(Duncan’s multiple range tests, P <0.05)
2.3 異養(yǎng)鞭毛蟲對不同密度銅綠微囊藻的抑制作用
異養(yǎng)鞭毛蟲對250×104~ 2 000×104cells/ mL密度的銅綠微囊藻均具有抑制作用,批量培養(yǎng)6 d后,其抑制率高達98.00% ~ 99.69%(圖4)。密度較低的微囊藻(250×104~ 500×104cells/ mL )的抑制率在實驗2 d即達到90%以上。當微囊藻密度較高(750×104~2 000×104cells/ mL)時,鞭毛蟲對微囊藻抑制率增長較慢,分別在實驗的4~6 d達到最高值。
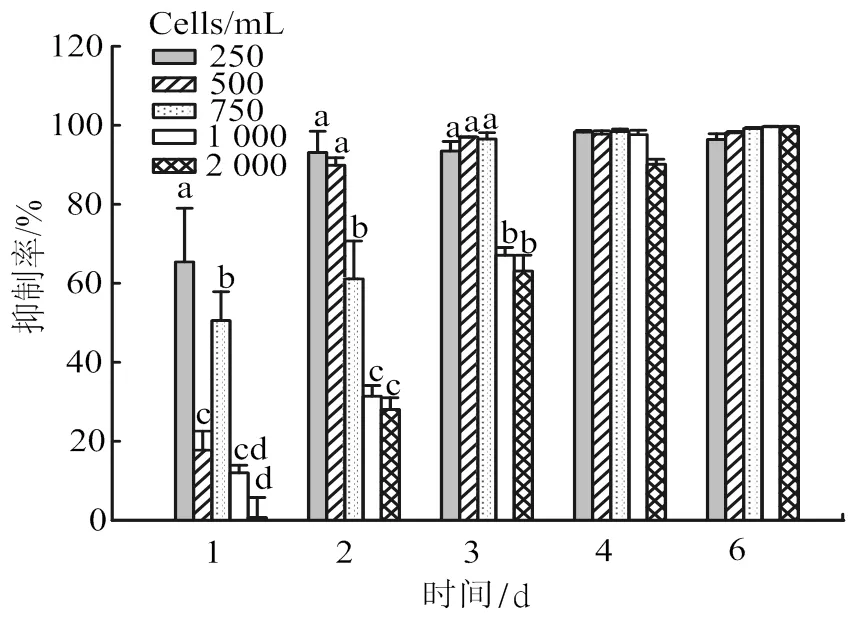
上標字母不同代表差異顯著(Duncan’s multiple range tests, P <0.05)
2.4 異養(yǎng)鞭毛蟲在不同銅綠微囊藻密度下攝食率變化
在不同微囊藻密度下,單個異養(yǎng)鞭毛蟲對微囊藻的攝食率高達38 ~ 49 cells/d(圖5)。在微囊藻密度為250×104~750×104cells/mL時,鞭毛蟲對微囊藻的攝食率呈現先增加后減少的趨勢;當微囊藻密度較高時(1 000×104、2 000×104cells/mL),鞭毛蟲在實驗1 d對微囊藻的攝食率為負值,隨后逐日增加。
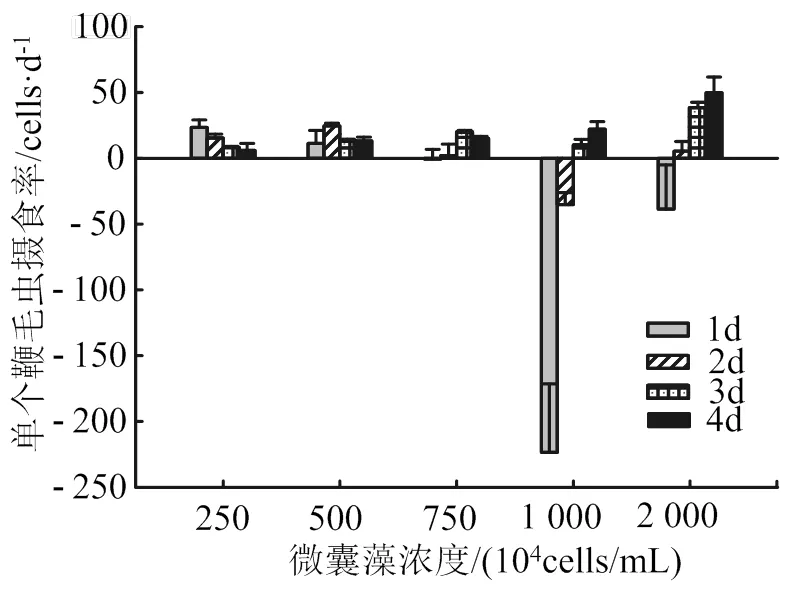
圖5 不同銅綠微囊藻密度下單個異養(yǎng)鞭毛蟲對銅綠微囊藻攝食率
3 討論
3.1 不同起始銅綠微囊藻密度對異養(yǎng)鞭毛蟲生長的影響
有些鞭毛蟲是混合營養(yǎng)型浮游生物,既能攝食有機顆粒物行異養(yǎng)生活,也可以進行光合作用合成有機物,如鞭毛蟲及sp.[15, 24]。本實驗中,當銅綠微囊藻密度為0時,異養(yǎng)鞭毛蟲的數量幾乎沒有增長,隨后急劇下降,表明無食物提供時,鞭毛蟲不能通過光合作用進行種群增長,證明本實驗所篩選的異養(yǎng)鞭毛蟲主要行異養(yǎng)生活,以吞噬有機物來進行種群增長。
Mohamed等[16]發(fā)現鞭毛蟲通過吞噬微囊藻,進而促進自身種群數量的增長,鞭毛蟲的生長率達0.71 ~ 0.99 d-1。本實驗中,異養(yǎng)鞭毛蟲在不同密度的微囊藻中均能有效攝食微囊藻,鞭毛蟲數量增長2 ~ 5倍,鞭毛蟲生長率高達0.65 ~ 0.74 d-1,與Mohamed的研究結果相似。在較低密度銅綠微囊藻時,異養(yǎng)鞭毛蟲種群增長最快,但是當銅綠微囊藻數量過低不足以支持異養(yǎng)鞭毛蟲生長時,異養(yǎng)鞭毛蟲的數量也隨之快速下降。然而,較高密度銅綠微囊藻在實驗1、2 d時抑制異養(yǎng)鞭毛蟲種群的增長(圖3)。Mohamed等[16]和Kim等[25]認為高濃度微囊藻所含毒素更多,鞭毛蟲消耗更多能量用于降解微囊藻毒素,從而使種群初始增長速度較慢。密度最高的2 000×104cells/mL濃度組中,異養(yǎng)鞭毛蟲生長率在實驗2、3 d顯著低于250×104~ 750×104cells/mL密度組,表明密度過高會抑制異養(yǎng)鞭毛蟲的種群增長。本實驗中,異養(yǎng)鞭毛蟲在銅綠微囊藻密度為250×104~ 2 000×104cells/mL中均能正常生長,其中,在微囊藻密度為500×104~1 000× 104cells/mL時維持了較高生長率。
3.2 異養(yǎng)鞭毛蟲對不同密度銅綠微囊藻攝食率的影響
鞭毛蟲可以攝食比其體積大幾倍顆粒[26]。已有研究報道鞭毛蟲可吞噬有機碎屑、細菌、綠藻、硅藻及藍藻等[13-14, 27]。鞭毛蟲主要通過胞吞作用攝取食物,胞吞作用可發(fā)生在蟲體多數部位,通過細胞膜內陷,把食物吞噬入體內形成食物泡[16, 28]。筆者在實驗過程中,觀察到單個鞭毛蟲體內包裹有十幾個微囊藻細胞。有研究表明,鞭毛蟲sp.能顯著減少微囊藻的密度,單個鞭毛蟲對微囊藻攝食率達18.3 ~ 30.8 cells/d[16, 25]。本實驗中,單個異養(yǎng)鞭毛蟲對銅綠微囊藻攝食率達13 ~ 49 cells/d。在微囊藻密度為250×104~ 750×104cells/mL時,隨實驗時間增加,鞭毛蟲對微囊藻攝食率呈現先增加后減少趨勢,表明當銅綠微囊藻的數量減少,鞭毛蟲對其攝食率也隨之下降。然而,微囊藻密度過高也會抑制鞭毛蟲攝食,當微囊藻密度較高時(1 000×104~ 2 000×104cells/mL),鞭毛蟲在實驗1 d對微囊藻的攝食率為負值,隨后才逐日增加。
3.3 異養(yǎng)鞭毛蟲對不同密度銅綠微囊藻的抑制作用
Zhang等[23]發(fā)現鞭毛蟲sp.在微囊藻密度低于1 000×104cells/mL時,均能有效降低微囊藻種群數量。本實驗中,異養(yǎng)鞭毛蟲對不同密度銅綠微囊藻均有抑制作用,其抑制率達98% ~ 99.69%,證明該鞭毛蟲具有較強抑制微囊藻的能力。然而,微囊藻濃度過高,鞭毛蟲對微囊藻數量的抑制能力下降。Zhang等[23]發(fā)現當微囊藻濃度高達3 200×104cells/mL時,鞭毛蟲對微囊藻無明顯抑制作用。本實驗中,異養(yǎng)鞭毛蟲對銅綠微囊藻抑制率隨時間而增大,低密度微囊藻組的抑制率較快達到高峰值99%,而高密度組的抑制率則緩慢增加。Nishibe等[29]認為銅綠微囊藻濃度過高,而異養(yǎng)鞭毛蟲密度過低,使異養(yǎng)鞭毛蟲對銅綠微囊藻的抑制能力上升較慢。Zhang等[17]也發(fā)現當微囊藻濃度不變,隨鞭毛蟲sp.的起始數量增加,鞭毛蟲對微囊藻抑制率從84%增加至99%。因此,異養(yǎng)鞭毛蟲對微囊藻的抑制效率也與兩者之間的相對數量比例有關。本實驗中,在銅綠微囊藻密度為250×104~ 2 000×104cells/mL,鞭毛蟲與微囊藻的數量占比為1∶50 ~ 1∶400的范圍內,異養(yǎng)鞭毛蟲對銅綠微囊藻均有抑制作用,其抑制率達98.00% ~ 99.69%。
Burkert等[30]發(fā)現棕鞭毛蟲sp.的牧食能激發(fā)銅綠微囊藻細胞活性,形成小型群體。本實驗中,鞭毛蟲對微囊藻的攝食并未誘發(fā)銅綠微囊藻形成防御群體,微囊藻因鞭毛蟲的攝食而死亡,形成部分無色絮狀團塊,表明本研究篩選的鞭毛蟲具有較高抑制微囊藻種群形成的能力。
3.4 異養(yǎng)鞭毛蟲在藍藻水華生物調控中的應用前景
De Kluijver等[8-11]利用穩(wěn)定同位素技術發(fā)現,藍藻水華發(fā)生時,藍藻是浮游動物的重要碳源,提出藍藻碳主要是通過微食物網途徑進入浮游動物,而不是通過直接攝食方式。Takamura等[31]發(fā)現霞浦湖里大部分微囊藻水華都在水里腐爛了。夏天高溫促進微囊藻水華的腐爛,從而形成微小碎屑,因此碎屑食物網(微食物網)是初級生產者微囊藻水華到浮游動物的主要途徑[32-34]。在水生態(tài)系統(tǒng)中,異養(yǎng)鞭毛蟲是微食物網的重要組成部分,參與了物質能量傳遞,是聯(lián)系細菌、初級生產者和浮游動物的中間環(huán)節(jié)[25]。本研究所篩選異養(yǎng)鞭毛蟲在微囊藻腐爛過程中大量增殖,是微囊藻降解過程中的先驅者[9]。實驗結果表明該異養(yǎng)鞭毛蟲能有效抑制銅綠微囊藻種群的增長。異養(yǎng)鞭毛蟲對銅綠微囊藻的攝食率高達每個鞭毛蟲13 ~ 49 cells/d,抑制率達98.3% ~ 99.69%,異養(yǎng)鞭毛蟲的生長率達 0.65 ~ 0.74 d-1。目前,利用生物抑藻方法控制藍藻水華已有一些研究,如利用細菌[35]、大型浮游動物、魚類[36]、病毒[37]、纖毛蟲[38]等方法,而培養(yǎng)利用鞭毛蟲清除藍藻水華是一種新探索。目前,異養(yǎng)鞭毛蟲種群數量在天然水體中的動態(tài)變化以及它對藍藻水華的數量控制動態(tài)及調控機理尚不清楚。因此,對于異養(yǎng)鞭毛蟲與藍藻水華的相互作用關系還有待進一步研究,以更好地利用異養(yǎng)鞭毛蟲實現藍藻水華的生物調控。
[1] 劉正文. 湖泊生態(tài)系統(tǒng)恢復與水質改善[J]. 中國水利, 2006(17): 30-33.
[2] 王朝暉, 林秋奇, 胡韌, 等. 廣東省水庫的藍藻污染狀況與水質評價[J]. 熱帶亞熱帶植物學報, 2004, 12(2): 117-123.
[3] HOLM, N P, SHAPIRO J. An examination of lipid reserves and the nutritional status offed[J]. Limnology and Oceanography, 1984, 29(5):1137-1140.
[4] WIEGAND C, PFLUGMACHER S. Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review[J]. Toxicol Appl Pharm, 2005, 203(3): 201-218.
[5] JULIAN HARTMANN H. Feeding ofandon mixtures of unicellular and filamentous algae [J]Internationale Vereinigung für theoretische und angewandte Limnologie: Verhandlungen, 1985, 22(5): 3178-3183.
[6] DE BERNARDI R, GIUSSANI G. Are blue-green algae a suitable food for zooplankton? An overview[J]. Hydrobiologia, 1990, 200(1):29-41.
[7] WIEGAND C, PFLUGMACHER S. Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review[J]. Toxicology and Applied Pharmacology, 2005, 203(3): 201-218.
[8] DE KLUIJVER A, YU J, HOUTEKAMER M, et al. Cyanobacteria as a carbon source for zooplankton in eutrophic Lake Taihu, China, measured by13C labeling and fatty acid biomarkers[J]. Limnology and Oceanography, 2012, 57(4):1245-1254.
[9] LUO X, LIU Z, GULATI R D. Cyanobacterial carbon supports the growth and reproduction of: an experimental study[J]. Hydrobiologia, 2015, 743(1):211-220.
[10] YU J, LI Y, LIU X, et al. The fate of cyanobacterial detritus in the food web of Lake Taihu: a mesocosm study using13C and15N labeling[J]. Hydrobiologia, 2013, 710(1):39-46.
[11] GER K A, URRUTIA-CORDERO P, FROST P C, et al. The interaction between cyanobacteria and zooplankton in a more eutrophic world[J]. Harmful Algae, 2016, 54:128-144.
[12] AZAM F, FENCHEL T, FIELD J, et al. The ecological role of water-column microbes in the sea. Marine ecology progress series1983.10(3): 257-263.
[13] DALEY R J, MORRIS G, BROWN S. Phagotrophic ingestion of a blue‐green alga by ochromonas[J]. The Journal of Protozoology, 1973, 20(1):58-61.
[14] COLE G T, WYNNE M J. Endocytosis ofby[J]. Journal of Phycology, 1974, 10(4):397-410.
[15] ZHANG X, WATANABE M M. Grazing and growth of the mixotrophic chrysomonad(Chrysophyceae) feeding on algae[J]. Journal of Phycology, 2001, 37(5):738-743.
[16] MOHAMED Z A, AL-SHEHRI A M. Grazing onand degradation of microcystins by the heterotrophic flagellate[J]. Ecotoxicology and environmental safety, 2013, 96:48-52.
[17] ZHANG X, HU H, MEN Y, et al. The effect ofabundance on production of intra-and extracellular microcystin-LR concentration[J]. Hydrobiologia, 2010, 652(1):237-246.
[18] KLEIN BRETELER W, SCHOGT N, BAAS M, et al. Trophic upgrading of food quality by protozoans enhancing copepod growth: role of essential lipids[J]. Marine Biology, 1999, 135(1):191-198.
[19] BEC A, MARTIN-CREUZBURG D, VON ELERT E. Fatty acid composition of the heterotrophic nanoflagellatesp.: influence of diet and de novo biosynthesis[J]. Aquatic Biology, 2010, 9(2):107-112.
[20] YAN C, LI J H, LI J J, et al. A heterotrophic nanoflagellate grazing on the toxic Cyanobacterium[J]. Annales De Limnologie-International Journal of Limnology: EDP Sciences. 2009, 45(1):23-28.
[21] 黃凌風, 潘科, 郭豐, 等. 我國海洋微型異養(yǎng)鞭毛蟲研究: 現狀與展望[J]. 廈門大學學報(自然科學版), 2007, 45(2): 62-67.
[22] BASU B, PICK F. Factors related to heterotrophic bacterial and flagellate abundance in temperate rivers[J]. Aquatic microbial ecology, 1997, 12(2):123-129.
[23] ZHANG X, HU H-Y, MEN Y-J, et al. Feeding characteristics of a golden alga (sp.) grazing on toxic cyanobacterium[J]. Water research, 2009, 43(12):2953-2960.
[24] VAN DONK E, CERBIN S, WILKEN S, et al. The effect of a mixotrophic chrysophyte on toxic and colony‐forming cyanobacteria[J]. Freshwater Biology, 2009, 54(9):1843-1855.
[25] KIM B-R, NAKANO S-I, KIM B-H, et al. Grazing and growth of the heterotrophic flagellateon the cyanobacterium[J]. Aquatic microbial ecology, 2006, 45(2):163-170.
[26] ARNDT H, DIETRICH D, AUER B, et al. Functional diversity of heterotrophic flagellates in aquatic ecosystems[J]. The Flagellates: Unity, Diversity and Evolution, 2000:240-268.
[27] ZHANG X, WATANABE M M, INOUYE I. Light and electron microscopy of grazing by(chrysophyceae) on a range of phytoplankton taxa[J]. Journal of Phycology, 1996, 32(1):37-46.
[28] 王進, 李建宏, 華秀紅, 等. 吞噬微囊藻的鞭毛蟲的培養(yǎng)[J]. 湖泊科學, 2005, 17(2):183-187.
[29] NISHIBE Y, KAWABATA Z I, NAKANO S-I. Grazing onby the heterotrophic flagellatein a hypertrophic pond[J]. Aquatic microbial ecology, 2002, 29(2):173-179.
[30] BURKERT U, HYENSTRAND P, DRAKARE S, et al. Effects of the mixotrophic flagellatesp. on colony formation in[J]. Aquatic ecology, 2001, 35(1):11-17.
[31] TAKAMURA N, IWAKUMA T, YASUNO M. Photosynthesis and primary production ofKütz. in Lake Kasumigaura[J]. Journal of plankton research, 1985, 7(3):303-312.
[32] HANAZATO T. Interrelations betweenand Cladocera in the highly eutrophic Lake Kasumigaura, Japan[J]. Internationale Revue der gesamten Hydrobiologie und Hydrographie, 2007, 76(1):21-36.
[33] HANAZATO T, ARAKAWA T, SAKUMA M, et al. Zooplankton community in Lake Suwa: community structure and its role in the ecosystem[J]. Japanese Journal of Limnology, 2001, 62(2):151-167.
[34] NANAZATO T, YASUNO M. Population dynamics and production of cladoceran zooplankton in the highly eutrophic Lake Kasumigaura[J]. Hydrobiologia, 1985, 124(1):13-22.
[35] JIA W, HUANG X, LI C A. Preliminary Study of the Algicidal Mechanism of Bioactive Metabolites of Brevibacillus laterosporus onin Prawn Ponds[J]. The Scientific World Journal, 2014, 2014:1-11.
[36] 陸開宏, 金春華, 王揚才. 羅非魚對藍藻的攝食消化及對富營養(yǎng)化水體水華的控制[J]. 水產學報, 2006, 29(6):811-818.
[37] 韓繼剛, 孟頌東, 葉寅, 等, 藻類污染生物防治新策略[J]. 微生物學報, 2001, 41(3):381-385.
[38] 閔智, 吳睿若, 張琨, 等. 草履蟲對銅綠微囊藻的吞噬能力研究 [J]. 中國給水排水, 2011, 27(13):63-66.
Effects of Heterotrophic Flagellates (sp.) on Growth Performance of
LUO Xiao-xia1,2,3,LI Chang-ling1,2,3,HE Xue-yi1, TAO De-qin1, DU Zhen-xiong1
(1.//2.,524088,;3.,518000,)
【】The growth status of heterotrophic flagellatessp. in different concentrations ofand its inhibitory effect onwere studied.【】was designated with six different concentrations, which were 0, 250′104, 500′104, 750′104, 1 000′104and 2 000′104cells?mL-1, respectively.【】sp. is mainly heterotrophic life and it could inhibit the growth ofwith concentration ranged from 250′104to 2 000′104cells·mL-1. The inhibition rate was 98 to 99.7%, and the ingestion rate of single flagellate was 13 - 49 cells·day-1. The number and growth rate of heterotrophic flagellates increased with the increased concentration ofand reached the highest peak at the concentration of 500′104- 1 000′104cells·mL-1. The number and growth rate of heterotrophic flagellates reached 21′104-24′104cells/mL, 0.6-0.74 d-1respectively. However, the excessive concentration ofdid not favor the population growth of heterotrophic flagellates, and the growth rate of heterotrophic flagellates in the 2 000×104cells·ml-1treatment was significantly lower than that of other concentration treatments.【】The flagellates can effectively inhibit the growth ofand the optimal M. aeruginosa concentration for the growth of heterotrophic flagellates is 500×104to 1 000×104cells·mL-1.
;heterotrophic flagellates; growth rate; inhibition rate; ingestion rate
Q954.4
A
1673-9159(2019)03-0038-08
10.3969/j.issn.1673-9159.2019.03.006
2019-01-02
廣東省自然科學基金項目(2018A030313212);廣東海洋大學自然科學研究項目(C17378);廣東海洋大學博士啟動項目(R17039) 大學生創(chuàng)新創(chuàng)業(yè)訓練計劃項目(CXXL2019043)
羅曉霞(1977-),女,博士,實驗師,水體生態(tài)修復。E-mail: xialemon@126.com
李長玲(1964-),女,教授,水產經濟動物繁殖生物學。E-mail:1094528615@qq.com
羅曉霞,李長玲,何雪怡,等. 異養(yǎng)鞭毛蟲對銅綠微囊藻的生長影響[J]. 廣東海洋大學學報,2019,39(3):38-45.
(責任編輯:劉嶺)
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
