2016年萊州灣扇貝養殖區浮游植物群落生態特征?
2019-06-14 12:06:38楊俊麗李希磊崔龍波
中國海洋大學學報(自然科學版) 2019年8期
楊俊麗, 李希磊, 于 瀟, 崔龍波
(煙臺大學生命科學學院, 山東 煙臺264005)
浮游植物是海洋生態系統中最重要的初級生產者,一方面浮游植物的光合作用為海洋提供了95%左右的生產力[1];另一方面為魚、蝦和扇貝等海洋生物提供餌料,其生長和代謝過程中釋放的溶解有機物也為細菌等提供了營養資源[2],因此,浮游植物群落結構間接或直接地影響著海洋生態系統的物質循環和能量流動。此外,浮游植物群落結構和種類的改變也受到海洋生態環境的影響,能夠在一定程度上反應海洋生態環境的狀況[3]。因此,對浮游植物群落結構的研究也是海洋生態研究的重要內容。
萊州灣是渤海三大海灣之一,位于山東半島北部,是魚、蝦、蟹以及貝類等經濟生物的主要繁育場,也是山東省重要的養殖漁業基地。近年來,對萊州灣浮游植物的研究已經有諸多報道[4-9],但對萊州灣養殖區浮游植物群落變化及月份變化卻鮮有報道,且多采用網采的方法,難免會漏掉部分個體小于76 μm的物種。本實驗采用有機玻璃采水器采集扇貝養殖區表層和底層的水樣,采取逐步濃縮的方法,避免了以往網采樣品的弊端,更加系統地對養殖區內浮游植物的群落組成、數量變化以及多樣性做出了分析,希望為萊州灣海域浮游植物群落和生態環境的變化提供背景資料,并為萊州灣漁業資源評估和管理以及合理的養殖開發提供科學依據。
1 材料與方法
1.1 采樣時間與站位設置
于2016年4—11月每月中旬進行采樣,根據萊州灣扇貝養殖區的分布狀況,選取了8個海灣扇貝養殖區,分別命名為L1~L8,具體位置見圖1。每個養殖區設置4個平行采樣點,最后取其平均值記錄實驗結果。
1.2 浮游植物的采集與處理
于2016年4—11月每月中旬在上述站位水域用有機玻璃采水器進行采樣,每個采樣點采集表層和底層混合等量水樣1 L,加入15 mL 10%的魯哥氏液,帶回實驗室,靜置24~48 h后濃縮至20~40 mL,暫儲存于50 mL磨口玻璃瓶中,觀察時將樣品搖勻,取0.1 mL滴于浮游植物計數板上,然后在顯微鏡下觀察鑒定,并記錄樣品中浮游植物的種類與數量[10]。
圖1 萊州灣海域扇貝養殖區調查站位
1.3 數據處理
采用物種優勢度Y(McNaughton)[11]判斷群落的優勢種,采用物種多樣性指數H′(Shannon-Wiener index)[12]、均勻度指數J(Pielou index)[13]和物種豐富度指數D(Margalef index)[14]來研究浮游植物群落的結構特征。計算公式分別為:
Y=(ni/N)fi;
J=H′/log2S;
D=(S-1)/InN。
式中:ni為第i種的總個數;N為所有種類的總個數;fi為該種在各樣品中出現的頻率;Pi為第i種的個體數與樣品中總個體數的比值;S為樣品中種類總數。
2 結果與分析
2.1 浮游植物種類組成
萊州灣8個扇貝養殖區在4—11月共鑒定出浮游植物5門96種,其中硅藻門最多,有63種,占所有物種的65.5%;其次為甲藻門和綠藻門,分別有14和9種,分別占所有物種的14.6%和9.4%;金藻門和隱藻門最少,各有5種,分別占所有物種的5.2%(見表1)。

表1 萊州灣扇貝養殖區浮游植物種類組成

續表1
所有扇貝養殖區在4—11月浮游植物各門類所含物種數見圖2。硅藻門10月份檢出物種數最多,有40種,4月份檢出物種數最少,有11種;甲藻門9月份檢出物種數最多,有11種,4月份檢出種類數最少,有2種;綠藻門5月份檢出物種數最多,有8種,9—11月份檢出種類數最少,各有1種;金藻門5、7和8月份各檢出4種,6和9月份未檢出;隱藻門4—8月份各檢出2~3種,其余月份均未檢出。
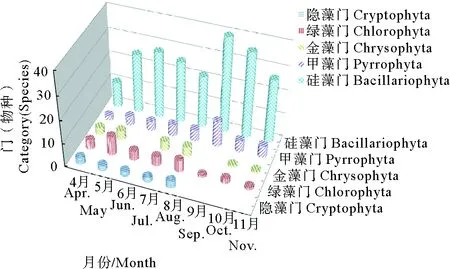
圖2 萊州灣扇貝養殖區浮游植物各門類所含物種數分布圖
2.2 浮游植物優勢種變化
本研究取優勢度(Y)≥0.02為優勢種,8個扇貝養殖區在4—11月浮游植物的優勢種及其優勢度見表2。由表2可知浮游植物在不同月份所形成的優勢種雖有所相同,但總體呈現演替狀態,有的浮游植物只在單一季節形成優勢種,如單鞭金藻、等鞭金藻、新月筒柱藻、針桿藻以及針狀菱形藻等只在春季形成優勢種;群生舟形藻、小球藻、舟形藻、尖布紋藻以及膝溝藻只在夏季形成優勢種;布紋藻、浮動彎角藻以及尖刺擬菱形藻等只在秋季形成優勢種。有的浮游植物在兩個季節出現,如扁藻、藍隱藻、柔弱幾內亞藻、隱藻、長菱形藻、圓篩藻、角毛藻以及斯氏幾內亞藻等。
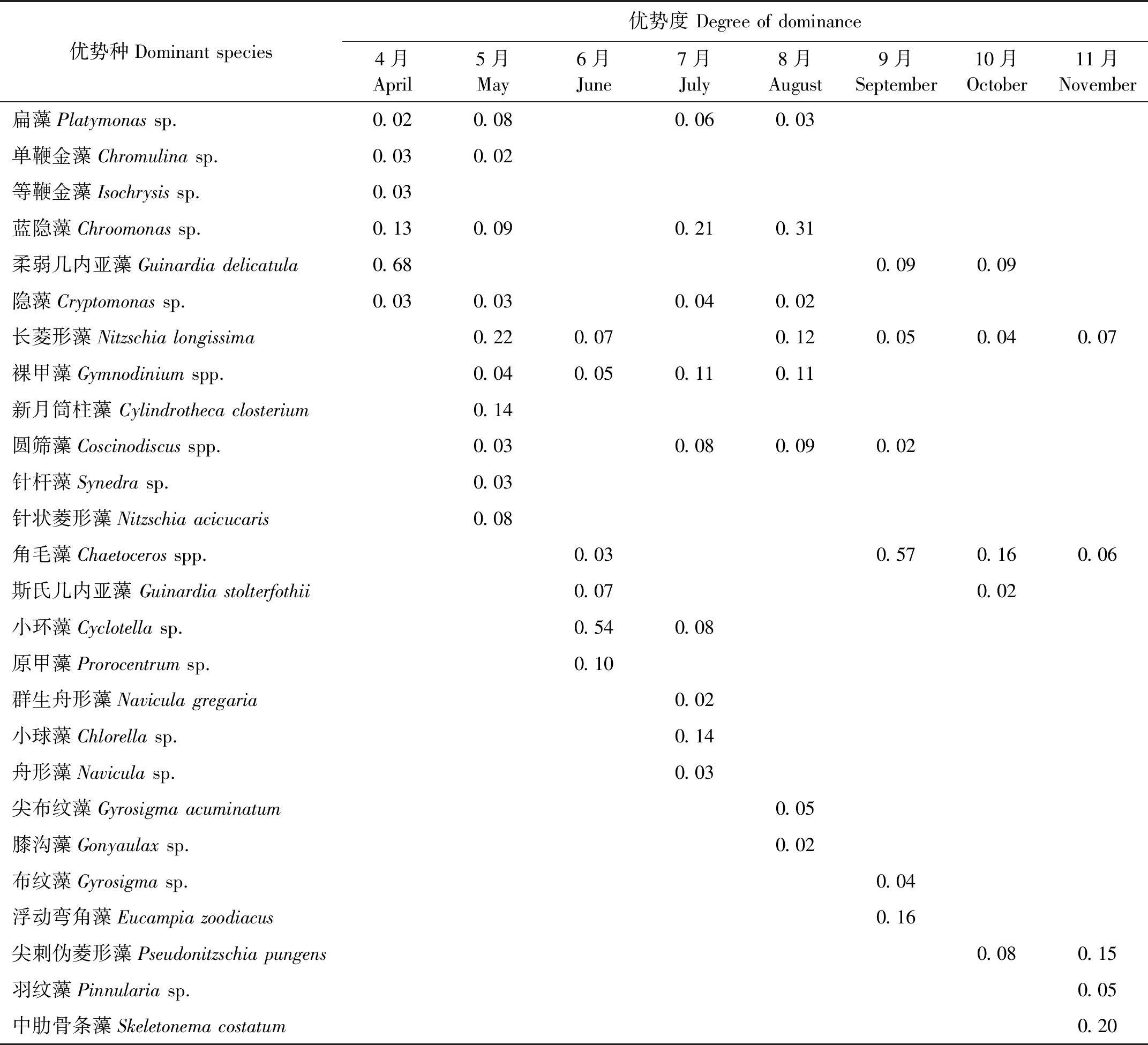
表2 萊州灣扇貝養殖區浮游植物優勢種及其優勢度
注:空白表示該物種在該月份未形成優勢。
Note: The gap indicates that the species did not have an advantage in this month.
2.3 浮游植物密度變化
萊州灣8個扇貝養殖區在4—11月浮游植物密度變化如圖3所示,春季(3和4月)密度變化范圍為3.72×104~19.90×104cell/L,平均值為8.86×104cell/L,不同養殖區之間密度差異較小;夏季(6—8月)密度變化范圍為1.62×104~112.04×104cell/L,平均值為14.11×104cell/L,其中6月份養殖區之間浮游植物密度分布差異較大,L1養殖區密度最大,L2養殖區密度最小,約相差9倍;秋季(9—11月)密度變化范圍為0.43×104~36.80×104cell/L,平均值為7.97×104cell/L,其中9月份L4和L5養殖區浮游植物密度明顯大于其他養殖區,10和11月份養殖區之間差異較小,整體明顯為6月浮游植物平均密度最大,11月份平均密度最小。
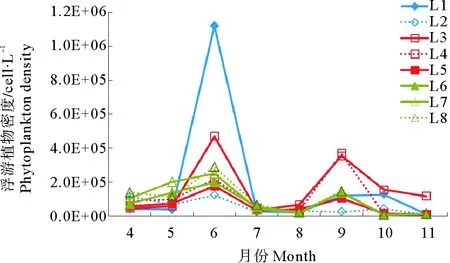
圖3 萊州灣扇貝養殖區浮游植物密度月份變化規律
2.4 浮游植物群落多樣性分析
萊州灣8個扇貝養殖區在4—11月浮游植物多樣性指數(H′)、均勻度(Y)以及物種豐富度指數(D)如表3所示。由表3可知,(1)8個扇貝養殖區在4—11月浮游植物多樣性指數的變化范圍為1.06~3.92,平均為2.86。L5和L6養殖區在4月、L1和L2養殖區在6月以及L3和L4養殖區在9月游植物多樣性指數(H′)較低,其余月份與其他養殖區一樣均處于動態變化之中。(2)8個扇貝養殖區在4—11月份浮游植物均勻度的變化范圍為0.26~0.96,平均為0.72。L1養殖區在6月以及L3養殖區在9月份浮游植物均勻度略小于0.3,物種分布較不均勻,但整體來看,8個扇貝養殖區浮游植物物種分布較均勻。(3)8個扇貝養殖區在4—11月份浮游植物豐富度指數的變化范圍為2.09~4.89,平均為3.27,同一養殖區在不同月份以及不同養殖區在同一月份差異均不大,整體處于動態波動之中,說明8個養殖區在8個月份浮游植物物種數分布較均勻。
3 討論
3.1 浮游植物種類變化
本研究通過對萊州灣扇貝養殖區浮游植物的調查,檢出硅藻門、甲藻門、隱藻門、金藻門和綠藻門共5門96種,除隱藻門在秋季未檢出外,其它4門在春、夏和秋均有檢出。程玲[7]、孫慧慧[9]以及吳雪[10]的研究結果表明萊州灣海域(包括養殖區海域)域浮游植物組成中,硅藻門在豐度和種類數上均占絕對優勢,其次是甲藻門。郭術津等[15]、孫雪梅等[16]以及苑明莉等[17]對渤海海域浮游植物的調查也得出相同結論,均與本研究結果較為一致。
3.2 浮游植物密度變化
萊州灣8個扇貝養殖區浮游植物密度呈較明顯的季節變化特征,最高值出現在夏季(6月),平均值為30.48×104cell/L,最低值出現在秋季(11月),平均值為1.98×104cell/L,歷史資料顯示,2003年[4]和2009[5]年萊州灣浮游植物數量均值最高值均出現在夏季(8月),最低值出現在春季(5月);2012—2014年[7]萊州灣浮游植物豐度最高值均在夏季(8月),最低值在秋季(11月);2015年[10]萊州灣扇貝養殖區浮游植物密度最高值在夏季(6月),最低值在秋季(11月),說明14年間萊州灣浮游植物群落結構的季節演替發生了變化,雖然浮游植物密度最高值都出現在夏季,但養殖區和非養殖區最高值出現的月份不相同,可能原因為7和8月份是扇貝的生長速度最快,對浮游植物的需求最大,期間絕大多數的浮游植物被扇貝攝取,所以養殖區內浮游植物密度在6月份最高而不是8月份。
本研究結果顯示,萊州灣8個養殖區浮游植物密度變化范圍為0.43×104~112.04×104cell/L,年平均值為10.60×104cell/L,其密度明顯大于萊州灣海域歷年研究結果,主要原因為歷年采樣均采用網采方法,相比本研究的水采方法,采樣過程中難免會漏掉一部分直徑小于淺水Ⅲ型浮游生物網孔徑的微藻,另一原因可能為浮游植物在物種粒徑上有趨于“小型化”的趨勢,個體數增加[18],比如,圓篩藻屬物種的地位有所下降,并逐漸被更小的小環藻(Cyclotellasp.)、藍隱藻(Chroomonassp.)、裸甲藻(Gymnodiniumspp. )以及偽菱形藻(Pseudonitzschiasp.)等取代。張建樂等[19]研究結果表明2007年北戴河貝類養殖區的浮游植物豐度年均值為1.14×104cell/L;王大鵬等[20]研究結果表明2010—2011年北海營盤新珍珠貝養殖區浮游植物豐度為2.80×104~9.60×104cell/L;陳麗梅等[21]研究結果表明2012年大神堂貝類養殖區浮游植物密度年均值為6.22×104cell/L;與其它養殖區相比,該養殖區浮游植物密度處于較高水平,可見浮游植物作為貝類的餌料,其細胞數量足以滿足扇貝生長的需求。

表3 萊州灣扇貝養殖區浮游植物多樣性分析表
3.3 浮游植物優勢種變化
優勢種對浮游植物群落結構的變化產生著重要影響,其優勢度越大,優勢種的地位就越突出。本研究選取優勢度(Y)≥0.02的種類為優勢種。8個養殖區浮游植物優勢種表現為既交叉又演替的狀態,有的優勢種在幾個月份交替出現,有的優勢種只在單個月份中出現。春季共有優勢種12種,夏季共有優勢種15種,秋季共有優勢種11種,其中春、秋季節絕對優勢種較為單一,除4月份藍隱藻(Chroomonassp.)形成絕對優勢種外,其余均屬于硅藻門,如柔弱幾內亞藻(Guinardiadelicatula)、長菱形藻(Nitzschialongissima)、新月筒柱藻(Cylindrothecaclosterium)、浮動彎角藻(Eucampiazoodiacus)、角毛藻(Chaetocerosspp.)、尖刺偽菱形藻(Pseudonitzschiapungens)和中肋骨條藻(Skeletonemacostatum),而夏季,絕對優勢種種類較多,有硅藻門的小環藻(Cyclotellasp.)和長菱形藻(Nitzschialongissima),隱藻門的藍隱藻(Chroomonassp.),甲藻門的原甲藻(Prorocentrumsp.)和裸甲藻(Gymnodiniumspp.)以及綠藻門的小球藻(Chlorella sp.),硅藻門相比春、秋季密度有所下降,其他門類的比例有所上升,特別是甲藻門。硅藻喜低溫,最適合的溫度通常低于18 ℃[22-23],而甲藻群落則與溫度呈顯著正相關[9],萊州灣海域春、秋季(除9月份)的水溫[9,24]均是硅藻生長的理想溫度,從而能促進其大量繁殖,形成明顯的優勢種,而夏季水溫較高.甲藻數量開始增加,且7和8月份又是扇貝生長最快的時期,對硅藻的需求量大大增加,從而使硅藻門的優勢地位下降,甲藻門優勢地位增強。
近10年的研究發現(見表4),優勢種絕大多數屬于硅藻門,偶爾出現甲藻門,且體積較大,而近2年隱藻門、綠藻門和金藻門的藻類也有優勢種出現,且大型浮游植物如圓篩藻(Coscinodiscusspp.)、三角角藻(Ceratiumtripos)和布氏雙尾藻(Ditylumbrightwellii)等雖有檢出,但其密度所占比例大幅度降低,優勢種有向“小型化”發展的趨勢。

表4 萊州灣鄰近海域浮游植物優勢種歷年資料的比較
注:*為本研究結果。Note:*is the content of this study.
3.4 浮游植物多樣性變化
多樣性指數(H′)可以反映浮游植物群落結構的復雜程度;均勻度(Y)和物種豐富度指數(D)可以反映群落個體和物種數的分布情況,三者均能在一定程度上反應浮游植物群落結構的特征。一般來說,多樣性指數(H′)、均勻度(Y)和物種豐富度指數(D)越高,該群落所屬的生態系統就越穩定和成熟,在浮游植物生物多樣性評價中常以多樣性指數小于1.0、均勻度指數小于0.30作為群落多樣性較差的標準[4]。本研究結果顯示,8個養殖區的多樣性指數(H′)、均勻度指數(Y)和物種豐富度指數(D)變化范圍分別1.06~3.92、0.26~0.96和2.09~4.89,多樣性指數(H′)均大于1,均勻度指數(Y)除L1養殖區(6月)和L3養殖區(9月)小于0.3之外,其它養殖區浮游植物均勻度均大于0.3,整體生物多樣性較好,物種分布較均勻,群落結構較復雜。此外,多樣性指數(H′)又可作為環境質量評估的依據,根據生物多樣性指數評價標準[26]:H′ > 3.0,生態環境質量優良;2.0≤H′ < 3.0,生態環境質量一般;1.0≤H′< 2.0,生態環境質量較差;H′<1.0,生態環境質量極差,可知4月L5和L6養殖區、6月L1和L2養殖區以及9月L3和L4養殖區生態環境質量較差,其余月份所有養殖區生態環境質量一般或者優良。
4 結論
(1)2016年萊州灣8個扇貝養殖區共檢出浮游植物5門96種,包括硅藻門、甲藻門、隱藻門、金藻門和綠藻門,其中以硅藻門種類數最多。
(2)浮游植物密度變化范圍為0.51×104~11 204×104cells/L,6和9月密度較高,呈現季節差異性。浮游植物優勢種主要有柔弱幾內亞藻、小環藻、長菱形藻、角毛藻、藍隱藻、尖刺菱形藻、尖刺偽菱形藻和中肋骨條藻。
(3)萊州灣8個扇貝養殖區浮游植物多樣性指數較高,其中生物多樣性指數在1.06~3.92之間,均勻度指數在0.26~0.96之間,物種豐富度指數在2.09~4.89之間,總體群落結構較復雜,生態環境較穩定。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
