禽流感病毒HA蛋白的原核表達與鑒定
2019-06-14 05:47:54李令臣侯力嘉吳勝昔李俊萱魯友銘梁望旺
重慶理工大學學報(自然科學) 2019年5期
李令臣,侯力嘉,吳勝昔,李俊萱,魯友銘,徐 緣,李 志,曾 政,梁望旺
(1.重慶理工大學 藥學與生物工程學院, 重慶 400054; 2.重慶市動物疫病預防控制中心, 重慶 401120)
禽流感,是由A型流感病毒引起的從呼吸系統病變到全身性敗血癥的一種高度接觸性急性傳染病[1-3]。據世衛組織稱,自2003年以來,全球已報告860例人感染H5N1型流感病毒病例,意味著該病毒具有感染人類的潛力。在不斷研究中,發現此類病毒可能對家禽工業和人類公共衛生構成潛在威脅,因此對病毒進行持續監測和鑒定對于預防任何可能的疾病暴發至關重要。
高致病性禽流感(HPAI)是由正黏病毒科A型流感病毒屬的H5亞型或H7亞型禽流感病毒(AIV)引起的[6],其基因組由8個分節段的單股負鏈RNA組成,共編碼11種蛋白[7]。血凝素蛋白(hemagglutinin,HA)是作為目前存在于甲型流感病毒(IAV)包膜上的最主要表面糖蛋白,許多研究都致力于了解HA的結構及其抗原性[8-9]。本研究擬在不改變HA蛋白氨基酸序列的情況下,根據大腸桿菌密碼子偏好性優化基因序列,化學合成HA全基因,將其克隆至原核表達載體pET28a(+)中,構建pET28a(+)-HA重組質粒,并轉化BL21大腸桿菌進行誘導表達,以期建立低廉、快速、高效的HA重組蛋白原核表達系統,為后續疫苗及診斷方法的研究奠定基礎。
1 材料與方法
1.1 菌株和質粒
實驗所用大腸桿菌BL21(DE3)、感受態DH5α菌株購自北京鼎國昌盛生物有限公司;所用pET28a(+)質粒由重慶理工大學基因工程實驗室惠贈。
1.2 主要試劑
DNA Marker、QuickCut BamHI、QuickCut EcoRI限制性內切酶,購自TaKaRa;小型質粒提取試劑盒、小量膠回收試劑盒,購自重慶威斯騰生物科技有限公司;SDS-PAGE凝膠配制試劑盒、蛋白質分子質量標準,購自北京鼎國昌盛生物有限公司。
1.3 序列分析與合成
根據從NCBI的基因數據庫GenBank中査取的禽流感 HA(ID=DQ023145.1)的基因組序列,利用DNAWorks2.4軟件進行密碼子優化設計。Primer Premier 5.0軟件設計1對特異性引物,通過設計在序列上游引物P1中引入EcoRI (堿基序列為GAATTC)限制性酶切位點,在序列下游引物P2中引入BamHI (堿基序列為GGATTC)限制性酶切位點,此引物交由上海生工生物工程有限公司合成。引物P1、P2序列如下:
P1:
5′-GACGAATTCACCAATGTGCCGGAATGG-3′
(含EcoRI 酶切位點);
P2:
5′-GGGGGATTCACCATCCTAATGATGCGG-3′
(含BamHI酶切位點)。
1.4 pET28a(+)-HA重組載體的構建
將含有禽流感HA 基因的TOP10甘油菌在無菌條件下劃線培養后,挑菌入小試管再培養。得到大量菌體后,離心并用DNA小量提取試劑盒提取含有HA基因的質粒。將所得質粒用BamHI和EcoRI進行雙酶切鑒定。利用瓊脂糖凝膠電泳得到有效切割的HA基因片段。鑒定正確的HA質粒轉化入表達宿主[BL21(DE3)]中,將其大量擴增,留取部分做菌株保存。
1.5 重組蛋白的誘導表達及純化
將鑒定成功的菌株大量擴增,使菌濃度OD600 nm值在0.6~0.8之間時,加入誘導劑IPTG。進行誘導溫度(20、25、30、37 ℃)、IPTG濃度(0.1 mmol/L、0.5 mmol/L、1 mmol/L)、誘導時間(8、10、12、16 h)的篩選。取出0.5 mL 菌液,10 000 r/min 離心 5 min,棄上清,沉淀重懸于 40 μL 1×PBS,并加入10 μL 5×SDS-PAGE 凝膠電泳上樣緩沖液,100 ℃變性 10 min進行 SDS-PAGE電泳。按優化后的最佳條件重新誘導表達,采用蛋白純化儀的Ni柱蛋白純化得到重組HA蛋白,利用電泳鑒定。
1.6 蛋白質印記(Western blot)分析
經過鎳柱純化后得到的HA重組蛋白要經過Western免疫印跡鑒定分析,具體步驟參見文獻[8]。
2 結果與分析
2.1 重組質粒pET28a(+)-HA酶切鑒定
重組質粒pET28a(+)-HA經BamHI和EcoRI雙酶切后電泳,分別切出與對應目的基因大小相符的條帶(1 740 bp)以及與空載體pET28a(+)大小相符約5 369 bp的另一條帶(見圖1)。這表明已成功構建重組表達載體。
2.2 重組蛋白的誘導表達
將pET28a(+)-HA經IPTG誘導破菌,分別取適量的沉淀(滅菌水溶解)和上清進行SDS-PAGE檢測。結果表明:當誘導條件為30 ℃、IPTG 1 mmol/L誘導8 h時,蛋白表達最高,在70 kD處有一條明顯的蛋白條帶(見圖2),與預期大小相一致,說明pET28a(+)-HA成功表達。且重組菌在上清和沉淀中顯示70 kD的條帶,上清表達明顯高于沉淀(見圖3、4)。
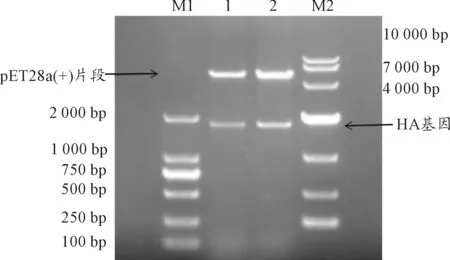
M1、M2:DNA分子質量標準;1、2:pET28a(+)-HA的雙酶切產物
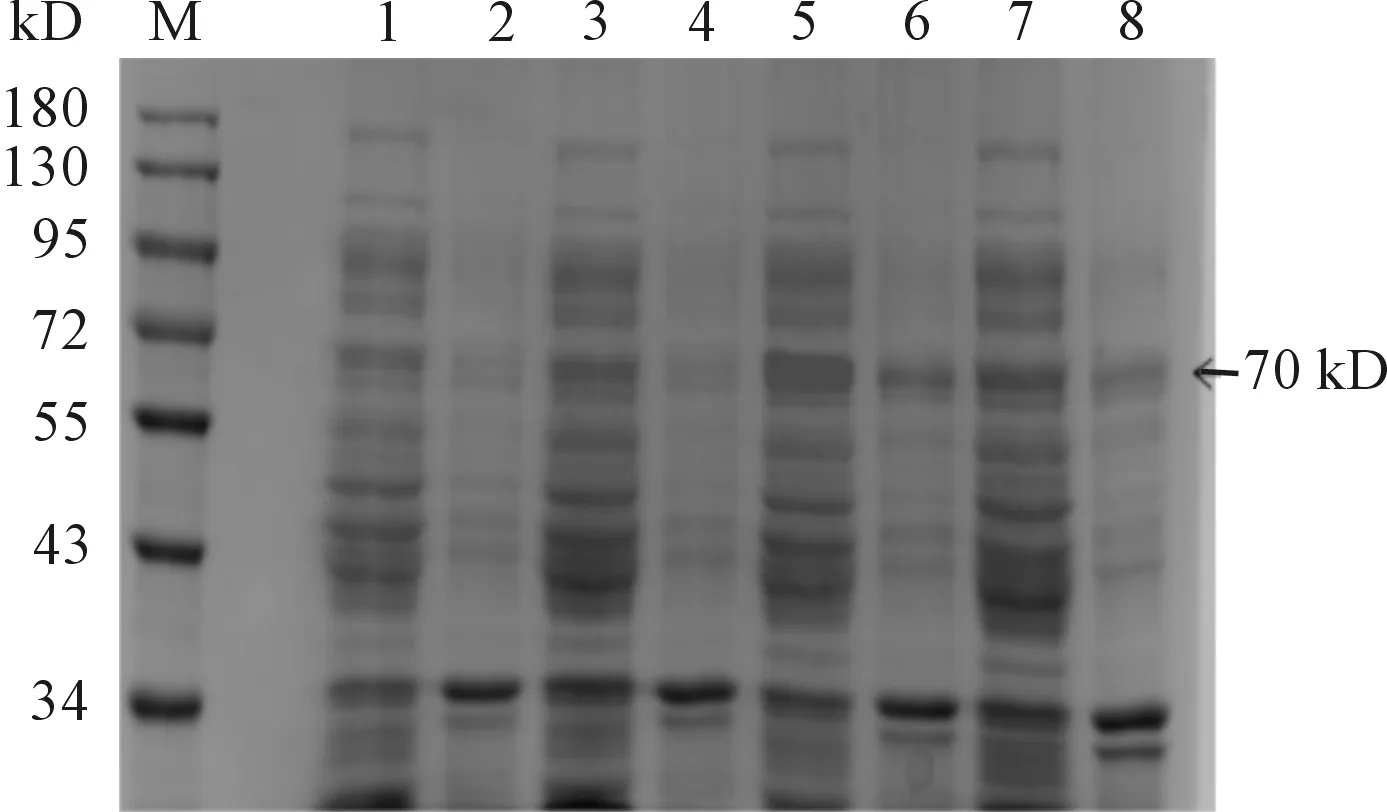
M:蛋白質分子質量標準;1、3、5、7:20、25、30、37 ℃(0.5 mmol·L-1IPTG,10h)條件下重組菌pET28a(+)-HA/BL21(DE3)經裂解液破碎后的上清; 2、 4、 6、 8: 20、25、30、37 ℃(0.5 mmol·L-1IPTG,10 h)條件下重組菌pET28a(+)-HA/BL21(DE3)經裂解液破碎后的沉淀
圖2 HA蛋白在不同誘導溫度條件下SDS-PAGE檢測結果
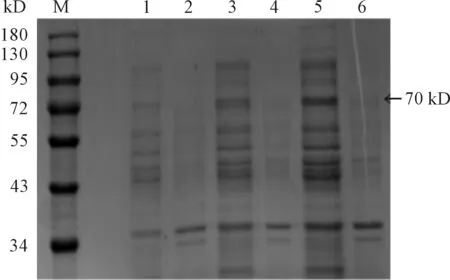
M:蛋白質分子質量標準;1、3、5:0.1、0.5 、1 mmol·L-1IPTG(30 ℃,10 h)條件下重組菌pET28a(+)-HA/BL21(DE3)經裂解液破碎后的上清; 2、 4、 6: 0.1、0.5、1 mmol·L-1IPTG(30 ℃,10 h)條件下重組菌pET28a(+)-HA/BL21(DE3)經裂解液破碎后的沉淀
圖3 HA蛋白在不同誘導劑量條件下SDS-PAGE檢測結果
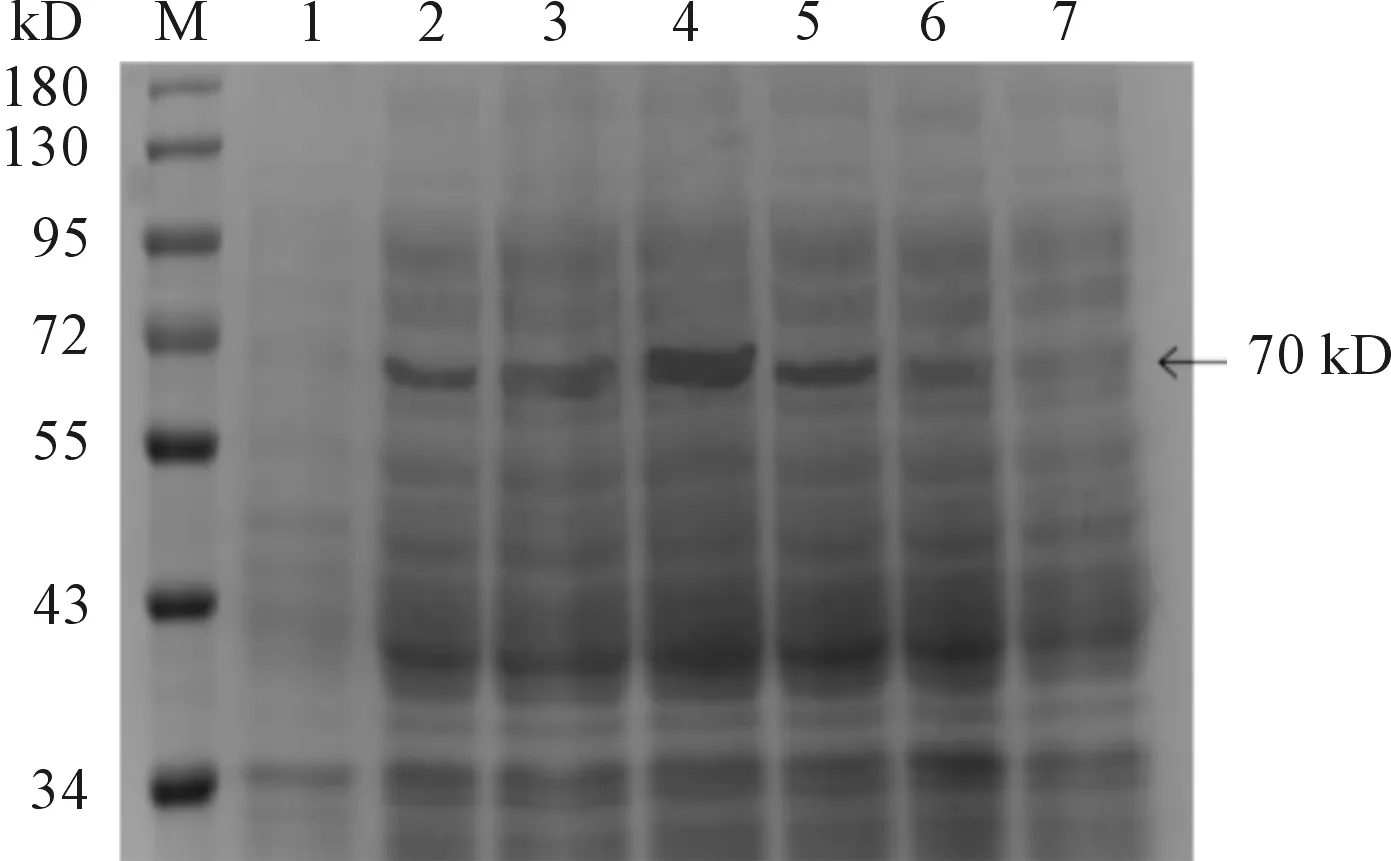
M:蛋白質分子質量標準;1、2、3、4、5、6、7:分別為2、4、6、8、10、12、14 h(1 mmol·L-1IPTG,30℃)條件下重組菌pET28a(+)-HA/BL21(DE3)經裂解液破碎后的上清
圖4 HA蛋白在不同誘導時間條件下SDS-PAGE檢測結果
2.3 重組蛋白PET28a(+)-HA純化及Western blot分析
利用最大濃度咪唑(imidazole)線性洗脫后確定最佳的洗脫濃度,并進行完全洗脫,收集洗脫液,經10% SDS-PAGE電泳鑒定(見圖5)。結果發現在150 mM的咪唑濃度可將大部分目的蛋白洗脫下來。純化后的重組蛋白經Western blot鑒定后發現:在70 kD處有特異性條帶,與預期的目的蛋白的大小相符(圖6)。
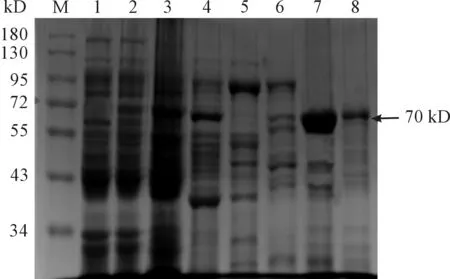
M:蛋白分子質量標準;1.空載體pET28a(+)感受態細菌超聲破碎后沉淀;2.重組菌pET28a(+)-HA/BL21(DE3) 經裂解液破碎后的上清;3.HA穿透液;4.20 mM咪唑洗脫蛋白;5.50 mM咪唑洗脫蛋白;6.100 mM咪唑洗脫蛋白;7.150 mM咪唑洗脫蛋白;8.250 mM咪唑洗脫蛋白。
圖5 HA純化蛋白的SDS-PAGE電泳結果

M:蛋白分子質量標準; 1.純化后的重組HA蛋白;2.空載體pET28a(+)感受態細菌對照
圖6 HA蛋白的Western blot檢測結果
3 討論
近年來隨著各類禽流感的多次大規模暴發,對感染的家禽只能大批量捕殺。隨著病毒加速變異,在國內也出現了多例關于人感染高致病性禽流感死亡的相關報道。禽流感 H5N1 亞型是能夠通過家禽傳染給人類的高致病型禽流感,每年疫情爆發時期均產生大規模的公眾恐慌。國內的研究人員在有關禽流感H5N1的病原學、致病機制、爆發規律以及預防措施方面都有較為深入的研究。本實驗采用的成本低廉、周期短的原核表達系統,被廣泛應用于診斷試劑、亞單位疫苗生產和蛋白功能的研究等方面[10-12]。
本研究根據 GeneBank 已發表的基因序列,設計并合成了HA的特異性引物,并選擇pET28a(+)作為表達載體導入大腸桿菌中,在載體上兩端都帶有多聚組氨酸標簽(His-tag),可以和鎳柱與咪唑發生相互作用,從而有利于蛋白的親和純化。在篩選最佳誘導條件中,經過對溫度、時間、IPTG濃度等因素的重復試驗,逐漸摸索出重組質粒PET28a(+)-HA在30 ℃、IPTG濃度在1 mmol/L誘導8 h時蛋白表達量最大。鑒定結果顯示:純化后的重組蛋白可達到較高的濃度,但純化結果略有雜蛋白,可能的原因在于蛋白洗脫濃度階梯需要進一步優化,后期將以更精準的咪唑濃度洗脫以獲得高純度的HA蛋白。在后期ELISA實驗中,所得重組蛋白與相關病毒血清有強烈反應(數值略),所以所得重組蛋白具有相似的免疫原性。吳艷菊等[13]將H5N1亞型的NS1 和NA蛋白進行包涵體原核表達,得到的蛋白免疫小鼠效果良好;鄒淑梅等[14]利用包涵體表達,利用8 mol/L的尿素洗脫蛋白,表達量占總蛋白的90%。對于可溶性表達,包涵體表達量高、蛋白折疊度好。但蛋白本身無活性,需要蛋白復性等操作,因此建議比較分析其他蛋白的原核誘導表達條件、異同及其原因。
本研究獲得的重組HA蛋白將為后續開展禽流感檢測方法及疫苗的研究提供參考。
