微小RNA-664a-3p在乳腺癌組織的表達及對乳腺癌細胞生物學行為的影響
2019-06-24 03:05:52岳青芬侯青霞韓聽鋒
醫學研究雜志 2019年5期
張 沛 岳青芬 侯青霞 韓聽鋒
乳腺癌是女性最常見的惡性腫瘤之一,中國乳腺癌發病人數僅次于美國,居全球第二位,且呈逐年上升趨勢[1, 2]。微小RNA(microRNA,miRNA)是一類內源性非編碼RNA,長度約為19~25個核苷酸[3]。miRNA通過與靶基因mRNA的3′-UTR配對結合,導致靶基因mRNA降解或者抑制其翻譯,負向調控靶基因的表達[4]。miR-664a-3p的研究尚處于起步階段,目前僅發現miR-664a-3p在骨肉瘤細胞中具有腫瘤抑制作用[5]。本研究通過qPCR檢測miR-664a-3p在乳腺癌組織和細胞株的表達,并將miR-664a-3p轉染到乳腺癌細胞中,觀察miR-664a-3p對乳腺癌細胞增殖和侵襲的影響,探討其影響乳腺癌細胞生物學行為的分子機制。
材料與方法
1.實驗材料:(1)一般資料:經筆者醫院醫學倫理學委員會同意,患者本人或家屬簽署知情同意書,收集筆者醫院婦產科2017年1月~8月經手術切除的新鮮乳腺癌組織標本16例,所有患者在手術前均未行放射治療、化學治療等。取其乳腺癌原發腫瘤組織和癌旁組織(距離癌灶邊緣>5cm)標本。經病理學診斷證實所得標本為乳腺癌組織,根據具體術中病理報告分為早期浸潤癌10例、浸潤癌6例。標本術中取出后立即放入液氮中保存。(2)細胞與試劑:RNA提取試劑盒和熒光實時定量試劑盒購自日本TaKaRa公司。DMEM/F12 培養基、RPMI 1640培養基和胎牛血清購自美國Gibco公司。工具細胞HEK293、人乳腺癌細胞株(T47D、MDA-MB-231、BT-549、MCF-7)和正常乳腺上皮細胞MCF-10A購自中國醫學科學院基礎醫學研究所。miR-664a-3p模擬物和miR-NC購自廣州市銳博生物科技有限公司。脂質體轉染試劑Lipofectamine 3000購自美國Invitrogen公司。pGL3熒光素酶檢測系統購自美國Promega公司。二喹啉甲酸(BCA)蛋白定量試劑盒購自武漢博士德公司。一抗β-actin、TEM8、CDK4、Cyclin D1、Vimentin、β-catenin和辣根過氧化物酶標記的二抗均購自美國Cell Signaling Technology公司。增強型化學發光(ECL)檢測試劑盒購自北京康為世紀生物科技有限公司。細胞周期檢測試劑盒購自中國凱基公司。Transwell小室購自美國Corning公司。
2.細胞培養及轉染:在37°C、5%CO2、飽和濕度的培養箱中,使用含10%胎牛血清的DMEM/F12培養基常規培養MCF-10A、T47D、MDA-MB-231和HEK293細胞,使用含10%胎牛血清的RPMI 1640培養基常規培養BT-549和MCF-7細胞。將對數生長期的T47D細胞以每孔2×105個接種于6孔板。依據Lipofectamine 3000轉染試劑盒說明書進行轉染,轉染濃度為50nmol/L。實驗細胞分兩組,即實驗組(轉染miR-664a-3p模擬物)和對照組(轉染miR-NC)。
3.熒光實時定量聚合酶鏈反應(qPCR)檢測miR-664a-3p和靶基因mRNA表達:Trizol試劑提取總RNA,依據RNA反轉錄試劑盒說明書反轉錄RNA為cDNA,取反轉錄產物qPCR反應,擴增條件為94℃ 5min,94℃ 30s,60℃ 20s,72℃ 20s,共40個循環。使用2-△△Ct法計算miR-664a-3p相對于內參U6的表達量及TEM8相對于內參GAPDH的表達量。引物序列詳見表1。
4.生物信息學軟件預測miR-664a-3p靶基因及雙熒光素酶活性檢測:生物信息學軟件miRWalk、TarBase和TargetScan預測miR-664a-3p的靶基因并取交集,miR-664a-3p的靶基因可能為TEM8,互補結合序列詳見圖1。構建野生型及突變型 TEM8-3′ UTR的報告基因質粒,突變區域詳見圖1,并將野生型、突變型分別與miR-664a-3p和miR-NC共轉染入HEK293細胞中,48h后按照雙熒光素酶檢測試劑盒說明書,檢測各組螢火蟲熒光素酶和海腎熒光素酶的活性。以螢火蟲熒光強度/海腎熒光強度比值反映各組的相對熒光強度。
5.Western blot法檢測目的蛋白表達:分別使用細胞裂解液提取兩組細胞總蛋白。取25μg蛋白,行10%聚丙烯酰胺凝膠電泳 (SDS-PAGE),電轉至聚偏氟乙烯(PVDF)膜,5%脫脂牛奶26℃封閉3h。一抗4℃孵育12h。Tris-HCl緩沖液漂洗2次,辣根過氧化物酶標記的二抗26℃孵育2h,Tris-HCl緩沖液漂洗2次,滴加增強型化學發光試劑(ECL)發光顯影分析比較條帶變化。
6.流式細胞術(FCM)檢測細胞周期:將兩組細胞消化收集,使用磷酸鹽緩沖液(PBS)漂洗2次,加入1ml 70%乙醇溶液4℃固定1h,離心去上清,PBS溶液漂洗2次,加入1ml碘化丙啶(PI)試劑,26℃避光孵育30min,流式細胞儀檢測并分析各組細胞周期,以細胞所處細胞周期百分率表示。
7.CCK-8法檢測細胞活力:將兩組T47D細胞重懸后計數,每孔4000個接種于96孔板,每間隔24h每孔加入CCK-8試劑10μl,培養箱內孵育4h,在酶標儀450nm波長處測定吸光度(A)值,連續檢測5天,以時間為橫軸,A值為縱軸,繪制細胞生長曲線。每組實驗重復4次。
8.細胞侵襲實驗:Transwell侵襲小室上室面鋪以細胞外基質(ECM)膠,培養箱內放置4h使ECM膠凝固。將轉染后48h的兩組T47D細胞消化,使用無血清培養基重懸并計數,以2×104個/孔接種于上室,下室中加入500μl完全培養基,放入培養箱內孵育24h,吸取培養基,小室內加入1ml甲醇固定20min,加入結晶紫溶液500μl染色20min,自來水清洗,棉簽擦去未穿過小室底膜的細胞,倒置顯微鏡下計數穿過小室底膜的細胞數目。

表1 qPCR引物序列
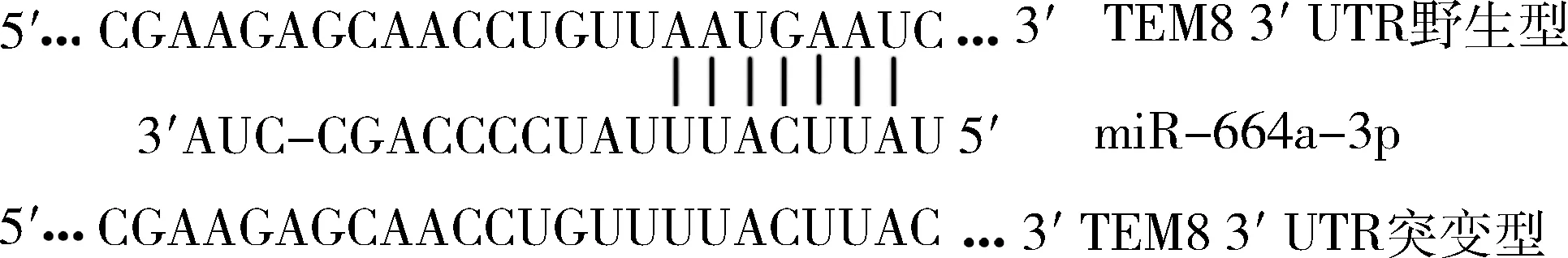
圖1 野生型TEM8 mRNA的3′-UTR區域含有miR-664a-3p靶向結合片段

結 果
1.miR-664a-3p在乳腺癌組織的表達:qPCR結果顯示(圖2),miR-664a-3p在乳腺癌組織和癌旁組織的表達量分別為1.03±0.09、2.10±0.20,miR-664a-3p在乳腺癌組織中表達量顯著低于癌旁組織(t=4.83,P<0.01)。
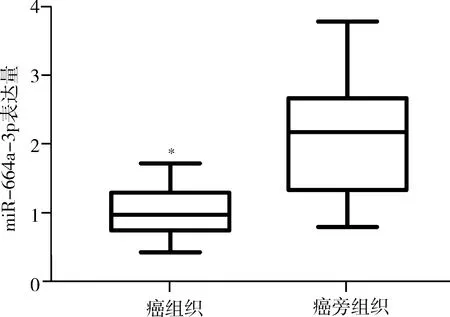
圖2 qPCR檢測miR-664a-3p在乳腺癌及癌旁組織的表達量與癌旁組織比較,*P<0.01
2.miR-664a-3p在乳腺癌細胞株的表達:qPCR結果顯示,miR-664a-3p在乳腺癌細胞T47D、MDA-MB-231、BT-549、MCF-7的表達量明顯下調,其中在T47D細胞中的表達量最低,與正常乳腺上皮細胞MCF-10A及其他乳腺癌細胞株比較,差異有統計學意義(P<0.05,圖3)。以T47D細胞為對象進行后續實驗。
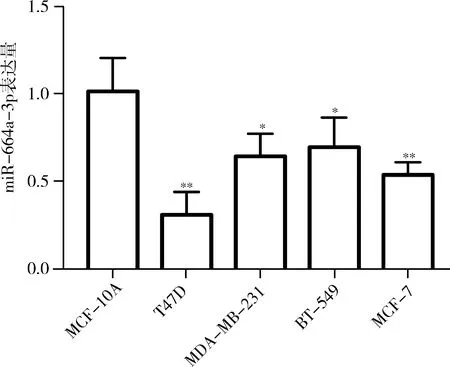
圖3 qPCR檢測miR-664a-3p在乳腺癌細胞株及正常乳腺上皮細胞的表達量與正常乳腺上皮細胞,*P<0.05,**P<0.01
3.轉染miR-664a-3p對T47D細胞miR-664a-3p表達的影響:qPCR結果顯示,T47D細胞轉染miR-664a-3p后miR-664a-3p表達較對照組明顯增加(24.87±2.17 vs 1.02±0.10,t=10.96,P<0.01)。
4.熒光素酶報告基因實驗:熒光素酶活性檢測結果顯示,miR-664a-3p可明顯抑制野生型 TEM8-3′ UTR質粒轉染細胞的熒光素酶活性(P<0.01),而對突變型TEM8-3′ UTR 質粒轉染細胞的熒光素酶活性并無影響(圖4)。

圖4 熒光素酶報告基因驗證miR-664a-3p與TEM8-3′ UTR的靶向結合與“miR-NC+野生型質粒”共轉染比較,*P<0.01
5.轉染miR-664a-3p對T47D細胞TEM8 mRNA表達的影響:qPCR結果顯示,T47D細胞轉染miR-664a-3p后TEM8 mRNA表達較對照組明顯下調(0.27±0.07 vs 1.00±0.01,t=9.97,P<0.01)。
6.轉染miR-664a-3p對T47D細胞TEM8相關蛋白表達的影響:Western blot法檢測結果顯示, T47D細胞轉染miR-664a-3p后TEM8、CDK4、Cyclin D1、Vimentin蛋白表達較對照組顯著下調,β-catenin蛋白表達較對照組顯著上調(圖5)。
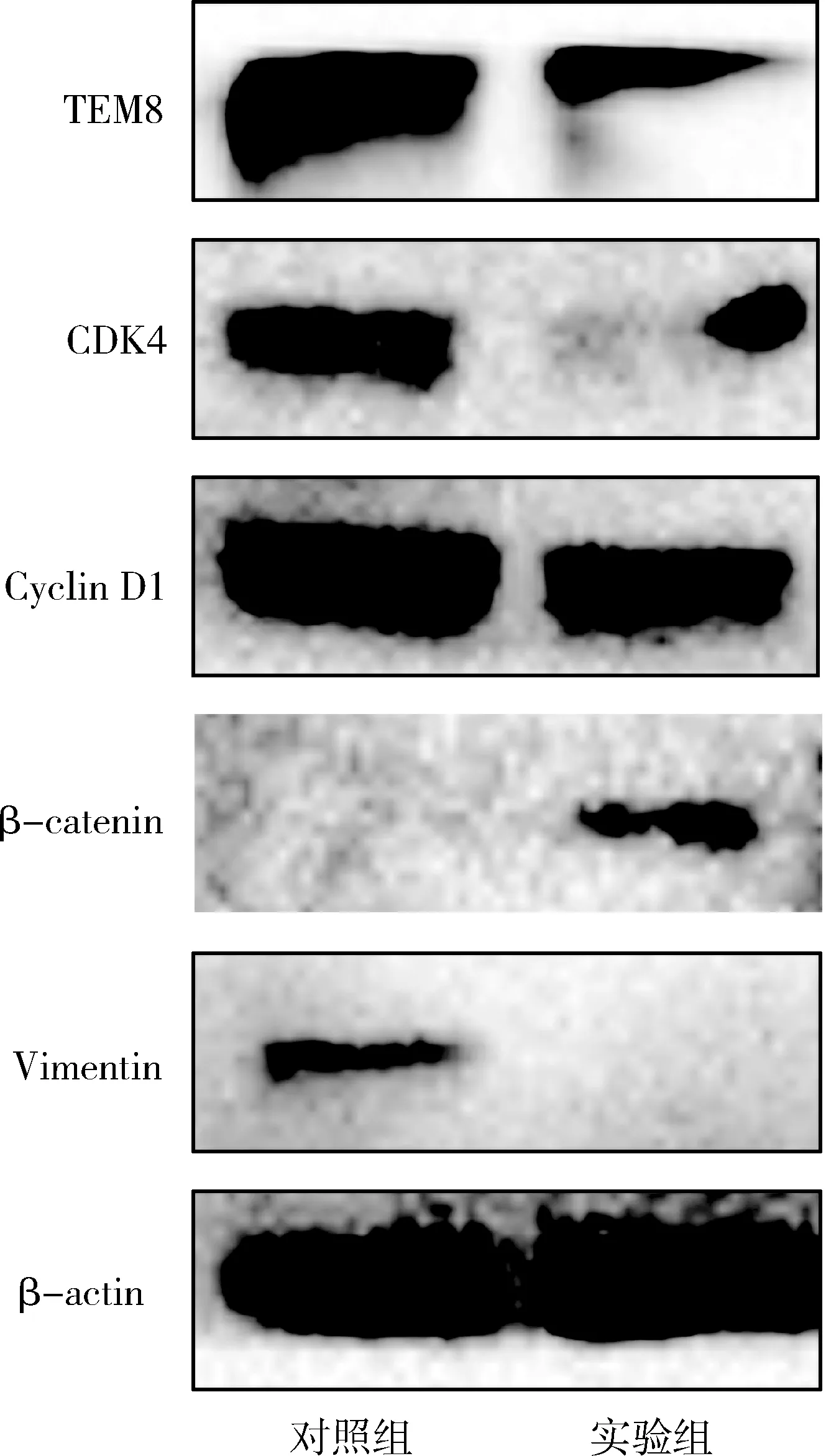
圖5 Western blot法檢測miR-664a-3p轉染T47D細胞后目的蛋白的表達水平
7.轉染miR-664a-3p對T47D細胞周期的影響:流式細胞術結果顯示,T47D細胞轉染miR-664a-3p后在G0/G1期的細胞比例較對照組明顯上升,高表達miR-664a-3p可抑制T47D 細胞周期的進展(P<0.01,表2)。
8.轉染miR-664a-3p對T47D細胞增殖能力的影響:使用CCK-8連續5天檢測T47D細胞的生長,當細胞生長至第4天和第5天,對照組細胞A值顯著高于實驗組,差異有統計學意義(P<0.01,圖6)。
9.轉染miR-664a-3p對T47D細胞侵襲能力影響:Transwell實驗結果顯示,在乳腺癌T47D細胞中高表達miR-664a-3p的細胞穿過小室底膜的細胞數明顯低于對照組(96.76±14.54 vs 217.50±17.96,t=5.22,P<0.01)。
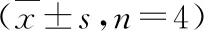
表2 miR-664a-3p對乳腺癌細胞周期的影響
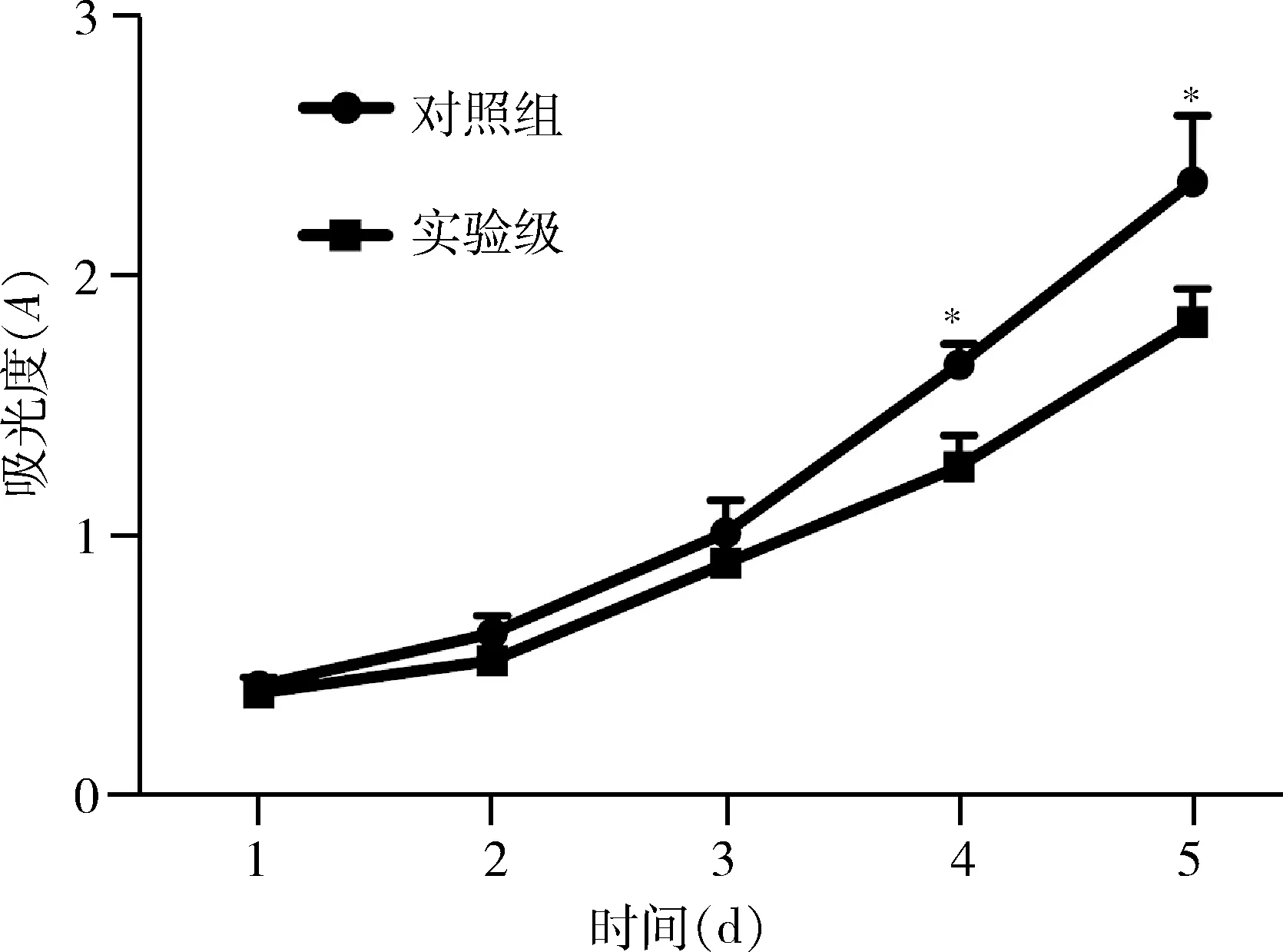
圖6 CCK-8 法檢測轉染miR-664a-3p后不同時點T47D細胞增殖能力的變化與對照組比較,*P<0.01
討 論
miRNA在轉錄水平對腫瘤細胞中的抑癌基因或癌基因的表達發揮重要的調節作用[6]。幾乎所有的腫瘤中均出現miRNA的異常表達,與腫瘤細胞的惡性生物學行為如增殖、侵襲轉移等密切相關[7]。miRNA的異常表達參與乳腺癌的發生和發展。miR-320、miR-770、miR-361-5p等miRNA通過抑制癌基因的表達負向調控乳腺癌細胞的增殖或遷移、侵襲[8~10]。miR-421、miR-1246、miR-27a等miRNA可抑制抑癌基因的表達,促進乳腺癌細胞的增殖、遷移和侵襲[11~13]。miR-664a-3p是miRNA家族的重要一員,可抑制骨肉瘤細胞的遷移和侵襲,然而其在乳腺癌中的作用目前尚不清楚[5]。
本研究結果顯示,miR-664a-3p在乳腺癌組織及乳腺癌細胞中低表達,提示miR-664a-3p可能與乳腺癌的發生、進展有一定的關系。生物信息學軟件分析顯示腫瘤內皮標志物8(TEM8)可能是miR-664a-3p的靶基因。以往的研究表明TEM8在腫瘤血管內皮細胞中高表達,通過促進內皮細胞的遷移進而利于腫瘤血管的生成,在腫瘤血管生成中發揮關鍵性作用[14]。近年來研究顯示,TEM8可促進腫瘤細胞的增殖和侵襲,在越來越多的腫瘤發生、發展中發揮重要作用[15]。有研究表明,TEM8在乳腺癌組織中高表達,其表達水平與患者的預后呈負相關[16]。本研究通過雙熒光素酶檢測實驗進一步驗證TEM8是miR-664a-3p的靶基因。本研究結果通過qPCR和Western blot法檢測結果發現,過表達miR-664a-3p后TEM8 mRNA和蛋白的表達均明顯降低。細胞周期蛋白依賴性激酶4(CDK4)和細胞周期蛋白D1(Cyclin D1)是正向調控細胞周期的重要蛋白。本研究顯示TEM8蛋白的下調引起CDK4和Cyclin D1蛋白表達降低。β-鏈蛋白(β-catenin)和波形蛋白(Vimentin)與腫瘤細胞遷移侵襲能力的變化密切相關,β-catenin蛋白的減少和Vimentin蛋白的增加均是腫瘤細胞遷移侵襲能力增強的表現。本研究顯示TEM8蛋白的下調引起β-catenin蛋白表達降低,Vimentin蛋白表達升高。本研究通過轉染miR-664a-3p至乳腺癌細胞T47D,發現T47D細胞周期進展明顯被抑制,細胞增殖能力和侵襲能力顯著降低,提示miR-664a-3p具有顯著的抑癌作用。
綜上所述,本研究表明miR-664a-3p在乳腺癌組織中處于高表達的狀態,通過抑制TEM8基因的表達,抑制細胞增殖能力和侵襲能力,在未來可能具有較好的臨床應用價值,并可能成為乳腺癌潛在的腫瘤標志物,用于乳腺癌的臨床診斷。
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
祝您健康(2018年5期)2018-05-16 17:10:16
