高原鼢鼠干擾下高寒草甸無脊椎動物分布與環境因子的關系
2019-07-19 07:28:12葉國輝張飛宇華銑澤花立民
草原與草坪 2019年3期
葉國輝,楚 彬,周 睿,張飛宇,華銑澤,花立民
無脊椎動物是陸地生物系統的重要組成部分,通過食物網參與物質循環與能量流動等生態系統過程,在維持陸地生態系統結構和功能方面具有重要調節作用[1-2]。在陸地生態系統中,植被、土壤、氣候和地形構成了無脊椎動物群落棲息環境。其中,植被和土壤是兩個主要因素[3],植物通過調節輸入土壤中有機體的品質和數量影響無脊椎動物食物資源。而土壤作為無脊椎動物生命的承載體,其屬性(結構和肥力)變化對土壤無脊椎動物生存和繁殖產生影響。近年來對土壤無脊椎動物研究主要集中在森林生態系統和農田生態系統中,對草地生態系統的研究相對較少[3]。而青藏高原高寒草地生態系統復雜多樣,對全球氣候變暖響應明顯,受氣候變化和人為因素等干擾呈退化趨勢[4],因此,針對這一敏感生態系統開展無脊椎動物對棲息地環境因子變化響應研究極其必要。
研究表明,植物群落和土壤理化性質等環境因子對土壤無脊椎動物群落和功能產生影響[5-6]。而土壤無脊椎動物受其體型大小影響,在土壤中活動能力相差懸殊,其中土壤無脊椎動物位于土壤食物網的頂端,其類群數和體內礦質元素與植物凋落物質量和礦質元素緊密相關[7],且植物凋落物的前期分解主要由土壤無脊椎動物完成[8]。因此,土壤無脊椎動物對植物和土壤環境因子變化響應比較敏感。在草地生態系中嚙齒動物是除家畜外重要的生物干擾源,它對草地物種多樣性,生產力以及水土保持等產生復雜而深遠的影響[9-10]。其中,高原鼢鼠(Easpalaxbaileyi)作為優勢鼠種之一,被譽為草地“生態系統工程師”[11],其推土造丘活動形成草地生境異質性斑塊,改變了草地原有植被物種組成以及土壤結構和營養。以往研究主要集中在高原鼢鼠干擾對草地植物群落組成、多樣性、生產力以及土壤肥力變化等方面[12-14]。高原鼢鼠干擾造成植物群落特征以及土壤理化性質變化,與土壤無脊椎動物的食物資源和棲息地環境條件緊密相關。因此,研究高原鼢鼠對地下生物影響可以進一步加強地下生物對環境條件變化響應的反饋研究。基于以上認識,在青藏高原祁連山東段典型高原鼢鼠棲息區,選擇同一放牧強度下3個不同高原鼢鼠相對種群密度棲息地,以鼠丘數量代表干擾強度,調查土壤無脊椎動物種類,數量以及植被和土壤狀況,并分析無脊椎動物類群對高原鼠兔干擾下土壤理化性質和植被性狀變化的響應。對于科學解釋高寒草甸無脊椎動物類群分布與環境因子關系,進而評價高原鼢鼠在草地生態系統中的功能和位置具有重要意義。
1 材料和方法
1.1 研究區概況
試驗地設于甘肅省武威市天祝藏族自治縣抓喜秀龍鄉,該區位于東祁連山金強河谷地,地理坐標為N 37°12′,E 102°46′,海拔2 937 m,地形受馬牙雪山和雷公山影響。年均溫-0.1℃,1月平均溫度-18.3℃,7月平均溫度12.7℃,>0℃年積溫1 380℃;年均蒸發量1 592 mm,年均降水量416 mm,多為地形雨,集中于7,8和9月,無絕對無霜期。植物生長期120~140 d。土壤以亞高山草甸土、亞高山黑鈣土為主[15]。植被類型為高寒草甸,以莎草科、禾本科植物為主,主要植物有矮嵩草(Kobresiahumilis)、線葉嵩草(Kobresiacapillifolia)、垂穗披堿草(Elymusdahuricus)、鵝絨委陵菜(Potentillaanserina)、蒲公英(Taraxacummaurocarpam)、平車前(Plantagodepressa)、二裂委陵菜(Potentillabifurca)、秦艽(Gentianamacrophylla)、扁蓿豆(Ruthenianmedic)。高原鼢鼠為該地區絕對優勢鼠種。
1.2 試驗設計與取樣
試驗選取7 hm2放牧強度一致的冬季牧場。于2017年6月下旬,依據高原鼢鼠造丘數量,將研究區劃分為高、中、低3個高原鼢鼠干擾區。各干擾區面積為100 m×100 m,每個區間隔50 m。在每個干擾區隨機設置5個土壤無脊椎動物采樣點,采樣點避開鼠丘。每個樣方面積0.5 m×0.5 m,取樣深度15 cm。
采用手撿法挑出土壤無脊椎動物,用75%乙醇溶液保存,帶回室內鑒定。由于部分土壤無脊椎動物幼蟲的科、屬鑒定比較困難,其次采集到的土壤無脊椎動物數量相對較少,可鑒定的科、屬難以用于數據處理分析。因此,此次研究無脊椎動物參考《中國土壤動物檢索圖鑒》[16],鑒定統計到目,并統計個體數量。根據土壤無脊椎動物調查樣點記錄樣方內植物物種數目,并剪掉樣方內植物,依據莎草科,禾本科和雜類草功能群分開,用烘箱75℃烘干稱取地上植物干重。樣地總蓋度采用步測法[17]。取樣調查土壤無脊椎動物采集點附近0~15 cm 土壤的理化性質。土壤含水量用環刀法測定,土壤緊實度使用土壤緊實度儀(Field scout SC900)測定。土壤有機質、全氮、全磷和全鉀含量測定方法參考《土壤理化分析》[18],土壤pH測定采用pHSJ-4A型pH計。
1.3 數據分析
1.3.1 植物物種豐富度 豐富度指數R=S(S為樣方中出現的物種數)。
1.3.2 無脊椎動物功能群劃分 依據無脊椎動物食性將其分為雜食性、腐食性、植食性和捕食性功能群[19]。植食性包括鞘翅目幼蟲(Coleoptera)、鱗翅目幼蟲(Lepidoptera)、同翅目成蟲(Homoptera)、半翅目成蟲(Hemiptera )、雙翅目成蟲(Dipterahrt)和柄眼目(Stylommatophora);捕食性包括蜘蛛目(Araneae)和鞘翅目成蟲(Coleoptera );腐食性包括雙翅目幼蟲(Dipterahr );雜食性有膜翅目蟻科(Hymenoptera:Formicidae)和雙尾目(Diplura)[6]。根據棲息生境條件劃分為土壤無脊椎動物(鞘翅目幼蟲、鱗翅目幼蟲、柄眼目、蜘蛛目、鞘翅目成蟲、雙翅目幼蟲膜翅目和雙尾目)和地上無脊椎動物(同翅目成蟲,半翅目成蟲和雙翅目成蟲),其中土壤無脊椎動物占據優勢。
1.3.3 多元排序 采用Canoco 4.5軟件分析無脊椎動物類群和數量與環境變量的關系。排序之前做除趨勢對應分析[20]。為評估環境因子對無脊椎動物數量的影響,分別計算每個環境因子的總效應和凈效應[21],采用CanocoDraw繪制RDA排序圖。
試驗數據用Excel 2013進行初步處理,用Spass 19.0中的LSD法進行數據的多重比較(a=0.05)。
2 結果與分析
2.1 不同高原鼢鼠干擾強度下植被群落特征和地上生物量變化
高原鼢鼠干擾下植物群落結構和地上生物量明顯不同(表1)。隨高原鼢鼠干擾強度增加,草地植被總蓋度和物種豐富度呈明顯下降趨勢,且高強度干擾區植被蓋度顯著低于中低強度干擾區(P<0.05)。植物物種豐富度未發生顯著變化(P>0.05)。莎草科和禾本科植物地上生物量伴隨高原鼢鼠干擾強度增加呈上升趨勢,高強度干擾度區莎草科和禾本科植物地上生物量顯著高于低干擾區(P<0.05);而高強度干擾區雜類草植物地上生物量顯著高于中、低干擾區(P<0.05)。
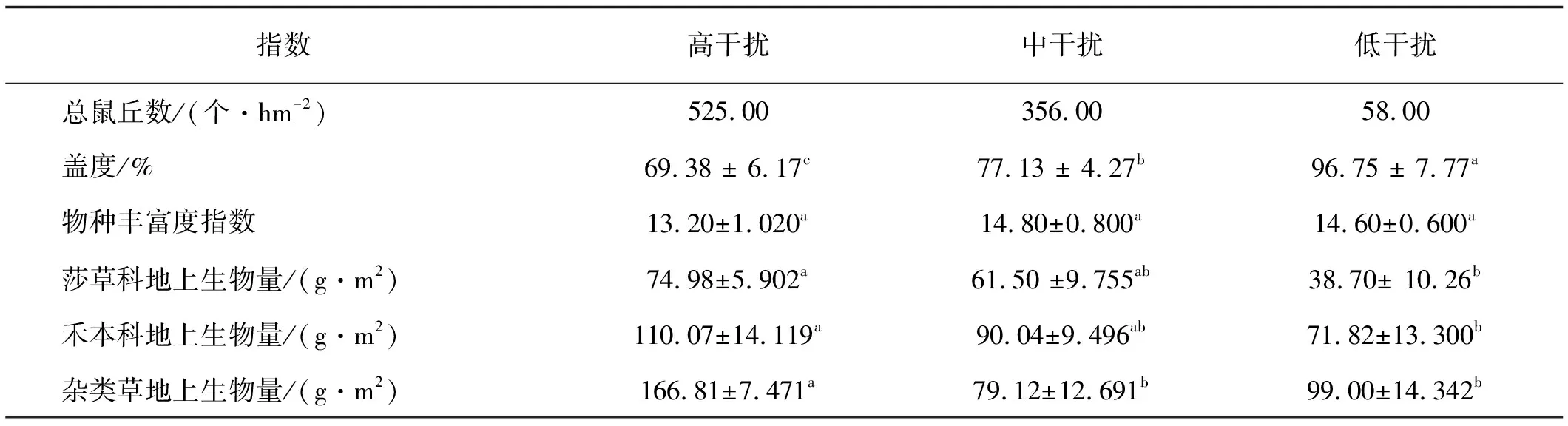
表1 高原鼢鼠干擾下的樣地變化
注:同行不同小寫字母表示差異顯著(P<0.05)
2.2 不同高原鼢鼠干擾強度下土壤理化性狀變化
土壤化學性質隨高原鼢鼠干擾差異不大(表2),其中土壤有機質,全鉀和pH在高原鼢鼠干擾下差異不顯著(P>0.05)。土壤全磷和全氮含量伴隨高原鼢鼠干擾強度增加呈降低趨勢,高強度干擾區土壤全磷含量顯著低于低干擾區(P<0.05),而中強度干擾區土壤全氮含量顯著低于低強度干擾區(P<0.05)。
土壤物理性質隨干擾強度變化不同,0~15 cm土層土壤緊實度伴隨高原鼢鼠干擾強度增加明顯下降,高干擾區土壤緊實度顯著低于低干擾區(P<0.05);土壤含水量呈相反變化趨勢,且高強度干擾區土壤含水量顯著高于低干擾區(P<0.05)。
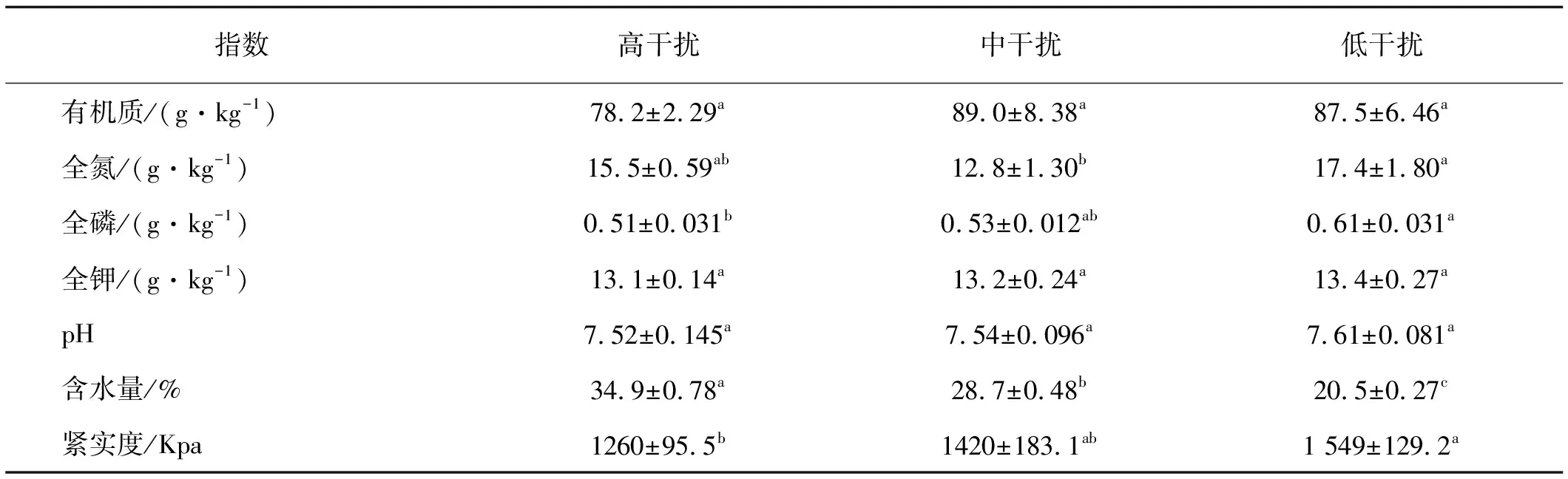
表2 不同高原鼢鼠干擾強度下表層土壤理化性狀
注:同行不同小寫字母表示差異顯著(P<0.05).
2.3 不同高原鼠兔干擾強度下無脊椎動物類群及數量變化
各干擾區共采集到無脊椎動物標本317個,分屬于3綱9目(鞘翅目、同翅目、雙尾目、蜘蛛目、柄眼目、雙翅目、膜翅目、半翅目和鱗翅目)。高,中和低強度干擾樣地無脊椎動物類群分別占9目,8目和8目。其中鞘翅目成蟲,柄眼目和鱗翅目幼蟲為各干擾區共有優勢類群,占各調查區域無脊椎動物總數量百分比大于10%;鞘翅目幼蟲為高干擾區和中干擾區優勢類群,分別占所調查區無脊椎動物總數量的10.27%和10.34%,而雙翅目幼蟲為中低干擾區優勢類群,占所調查區無脊椎動物總數量的32.18%和12.09%。雙尾目、膜翅目成蟲、半翅目成蟲和雙翅目幼蟲為高干擾樣地的常見類群,同翅目成蟲、雙翅目成蟲、膜翅目成蟲、半翅目成蟲和雙尾目為中度干擾區常見類群,蜘蛛目、同翅目成蟲、雙尾目、膜翅目成蟲、雙翅目成蟲和鞘翅目幼蟲為低強度干擾樣地常見類群。高,中和低干擾區無脊椎動物密度分別為112,73和68只/m2,且高強度干擾區無脊椎動物密度顯著高于中低強度干擾區(P<0.05)(表3)。
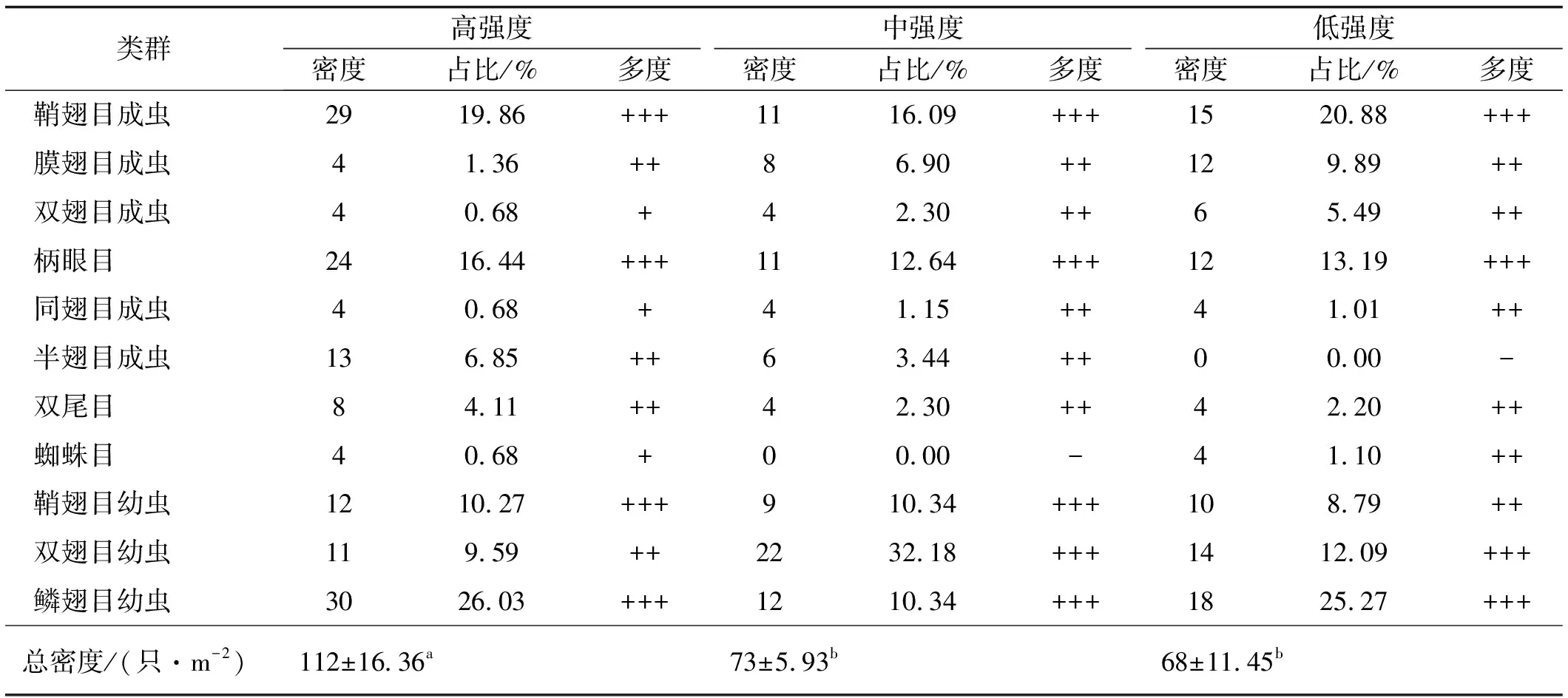
表3 高原鼢鼠不同干擾強度下無脊椎動物的類群和密度
注:“+++”為優勢類群,占總個體數10%以上;“++”為常見類群,占總個體數1%~10%;“+”為稀有類群,占總個體數1%以下;“-”表示沒有采集到標本。
單因素方差分析結果表明,不同高原鼢鼠干擾強度下無脊椎動物功能群密度在不同生境中存在差異(圖1)。植食性無脊椎動物密度在各干擾區占據優勢,且植食性無脊椎動物密度隨高原鼢鼠干擾強度增加呈明顯升高趨勢,高強度干擾區植食性無脊椎動物密度顯著高于低強度干擾區(P<0.05)。捕食性無脊椎動物密度呈現升高趨勢,但差異不顯著(P>0.05)。而腐食性和雜食性無脊椎動物類群密度伴隨高原鼢鼠干擾強度未呈現規律性變化,表明植食性無脊椎動物對植被生境條件的變化比較敏感。
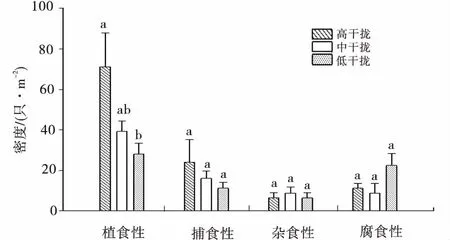
圖1 高原鼢鼠不同干擾強度下無脊椎動物功能類群密度Fig.1 The density of functional groups of invertebrate in the plots under different disturbance intensities of plateau zoker
2.4 高原鼢鼠干擾下影響無脊椎動物類群的環境因素
利用冗余分析(RDA)對高原鼢鼠干擾下無脊椎動物類群與環境因子排序(圖2),沿著軸1方向,不同干擾強度區無脊椎動物類群分布主要受土壤緊實度的影響,軸2方向表明土壤動態分布主要受土壤含水量和生物量等因子的影響,兩軸共解釋了51.8%的環境因子對無脊椎動物數量的影響。同翅目(S5)、半翅目(S7)、鞘翅目幼蟲(S9)、雙翅目幼蟲(S10)、鱗翅目幼蟲(S11)、柄眼目(S8)、雙尾目(S2)和蜘蛛目(S3)類群分布主要受土壤含水量和地上生物量影響,其次受土壤緊實度影響(負相關)。綜合分析表明不同干擾區土壤無脊椎動物優勢類群與其土壤物理性質相關性強。
不同干擾區無脊椎動物類群分布不同(圖3),高干擾區無脊椎動物調查樣方在排序軸1的上方分布,而中低干擾區無脊椎動物調查樣方基本在排序軸1下方分布,表明無脊椎動物群落結構在不同干擾區存在差異。同時鞘翅目成蟲(S1)、雙尾目(S2)、蜘蛛目(S3)、同翅目(S5)、半翅目(S7)、鞘翅目幼蟲(S9)、雙翅目幼蟲(S10)、鱗翅目幼蟲(S11)和柄眼目(S8)數量在高干擾區較高;而雙翅目(S4)、膜翅目(S6)和雙翅目幼蟲(S11)數量在低干擾區較高。
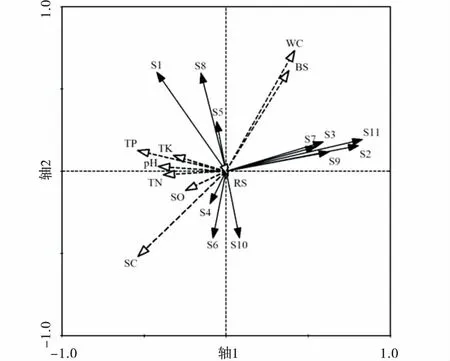
圖2 無脊椎動物類群與環境因子排序Fig.2 RDA biplot between environmental factors and soil animal communities注:圖2代碼S1~S11表示的無脊椎動物類群依次為鞘翅目、雙尾目、蜘蛛目、雙翅目、同翅目、膜翅目、半翅目、柄眼目、鞘翅目幼蟲、雙翅目幼蟲、鱗翅目幼蟲,下同
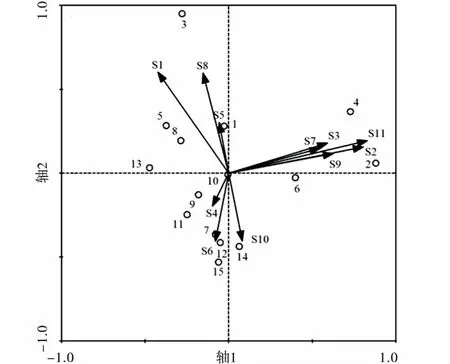
圖3 無脊椎動物類群與干擾區樣方排序Fig.3 RDA biplot between disturbance zone plots and soil animal communities注:圖3數字1~5表示高強度干擾區采樣點樣方,數字6~10表示中強度干擾區采樣點的樣方,數字10~15表示低強度干擾區采樣點的樣方
通過蒙特卡羅置換檢驗和偏蒙特卡羅置換檢驗評估每個環境因子對無脊椎動物類群分布影響的解釋貢獻,檢驗各環境因子的膨脹系數( Inflation factors,IFs) 均小于10,不受因子自相關性影響。經Monte Carlo 隨機置換檢驗999次(表4),結果表明9個獨立的環境解釋變量中,土壤緊實度和土壤含水量對土壤無脊椎動物類群分布解釋總效應達到顯著水平(P<0.05),而其余因子對土壤無脊椎動物類群分布解釋的總效應未達到顯著水平(P>0.05)。除去協同變量影響,各環境因子解釋變量均無顯著影響(P>0.05)。
注:* 表示在0.05 水平(雙側)上顯著相關,**表示在0.01 水平(雙側) 上顯著相關
3 討論
3.1 高原鼢鼠干擾對無脊椎動物群落影響
伴隨高原鼢鼠干擾強度增加,草地植被蓋度和地上生物量發生顯著變化。高強度干擾區植被總蓋度顯著低于低干擾區(P<0.05),而莎草科,禾本科和雜類草地上生物量在高干擾區顯著較高(P<0.05)。由于單位面積內高干擾區鼠丘數量明顯高于低干擾區,造成植被總蓋度顯著較低,衛萬榮[12]研究結果也表明草地植被蓋度伴隨新鼠丘數量增加顯著降低。高原鼢鼠通過采食和推土造丘活動改變了草地植被群落組成。于海玲等[14]研究結果顯示伴隨鼠丘數量增加,以禾草和莎草為主的植物優勢類群向雜類草群落轉變。劉錦上[11]結果表明相比無鼠洞道對照,鼠道上方植物生物量明顯較高;其次,Yurkewyca等[22]研究發現囊鼠活動強度增加,會導致鼠丘土壤營養礦化,促進鼠丘邊植物生長。試驗表明高強度干擾區植被受鼠丘微地形和鼢鼠地下復雜洞道影響植物生長趨勢明顯優于低干擾區,從而造成植物地上生物量顯著較高。伴隨高原鼢鼠干擾強度增加,高干擾區土壤無脊椎動物密度顯著高于低強度干擾區(P<0.05);植食性無脊椎動物密度變化同無脊椎動物總密度變化趨勢一致,高強度干擾區植食性無脊椎動物密度最大,而捕食性,腐食性和雜食性類群密度變化不大。表明高原鼢鼠干擾下植食性類群密度對地上植物生物量變化響應明顯。研究表明地上植物群落組成能夠影響土壤無脊椎動物群落組成和結構[5],其原因是生產力高的植物群落不僅為無脊椎動物提供充足食物資源滿足自身需求,同時為無脊椎動物提供合適的棲息環境。試驗表明高強度干擾區禾本科和雜類草地上生物量顯著高于低干擾區,是造成土壤無脊椎動物密度和植食性類群密度顯著高于低干擾區原因之一。RDA 排序圖顯示植食性類群密度(鞘翅目幼蟲、鱗翅目幼蟲和柄眼目等)與地上生物量具有相關性。同樣張洪芝等[6]研究表明植食性類群密度和地上地下生物量呈正相關。因此,高原鼢鼠干擾下無脊椎動物類群密度(植食性功能群)對生態系統自上至下控制響應明顯。
3.2 土壤緊實度和含水量對土壤無脊椎動物類群影響
已有研究表明土壤無脊椎動物類群受植物群落和土壤理化性質變化影響[5-6]。高原鼢鼠干擾下土壤物理性質發生明顯改變,高干擾區土壤緊實度顯著低于低干擾區(P<0.05)。此次研究高強度干擾區鼠丘數量達到525個/hm2,明顯高于低干擾區鼠丘數量。因此,受高原鼢鼠造丘活動影響,高干擾區地下復雜洞道系統,破壞了致密草氈層結構,從而顯著降低了表層土壤緊實度。周延山等[23]研究發現絕對鼠密度為28.33只/hm2,相比絕對鼠密度14.67 只/hm2,0~20 cm土壤緊實度顯著較低,而0~30 cm土壤含水量顯著較高。同樣,伴隨高原鼢鼠干擾強度增加,土壤含水量顯著升高(P<0.05),同土壤緊實度變化趨勢相反。高寒草甸植被群落主要以莎草科和雜類草占據優勢,其致密根系層嚴重阻礙降水下滲,由于研究區受降水影響,低緊實度高原鼢鼠干擾區表層土壤含水量明顯較高。而土壤無脊椎動物類群密度變化趨勢同土壤緊實度和含水量變化相同,表明土壤緊實度和含水量是造成土壤無脊椎動物類群密度變化的主要環境因子。
RDA分析結果表明土壤無脊椎動物類群密度與土壤因子(緊實度和含水量)顯著相關(P<0.05)。土壤無脊椎動物優勢類群密度(鞘翅目幼蟲和成蟲、鱗翅目幼蟲、柄眼目)與土壤含水量呈正相關,而與土壤緊實度呈負相關關系(圖2)。已有研究結果表明環境因子與土壤無脊椎動物類群具有相關性,土壤容重與土壤無脊椎動物類群數和密度呈負相關[5],如鞘翅目和蜘蛛目密度與土壤容重呈負相關[24];土壤無脊椎動物類群密度與含水量呈極顯著正相關[24],Set?l?等[25]也表明濕度是影響土壤無脊椎動物群落的主要氣候因子。此外,已有研究表明土壤化學因子和植被組成與無脊椎動物類群數和密度具有相關性。吳鵬飛等[5]研究發現土壤無脊椎動物密度與pH、有機質和全氮具有顯著相關性。不同土壤無脊椎動物類群有其主要的環境影響因子,如雙翅目幼蟲、鞘翅目和鱗翅目與土壤有機質和全氮呈顯著正相關[24],鞘翅目和半翅目與土壤全鉀呈正相關[6];其次,劉繼亮等[26]研究土壤功能群密度與植物物種豐富度呈顯著正相關。此次研究表明伴隨高原鼢鼠干擾強度變化,土壤化學因子(有機質、全鉀、全氮和pH)和植物物種豐富度未發生顯著變化(P>0.05),因此,土壤化學因子和植物物種豐富度對土壤無脊椎動物類群密度影響不明顯。
4 結論
試驗調查到祁連山東段高寒草甸無脊椎動物共3綱9目,其中鞘翅目成蟲、柄眼目、鱗翅目幼蟲為土壤無脊椎動物優勢類群。高原鼢鼠干擾對無脊椎動物密度和植食性功能群密度具有顯著影響,其通過推土造丘活動明顯改變了土壤物理性狀(緊實度和含水量),進而影響無脊椎動物類群和密度。RDA分析結果表明,土壤緊實度和含水量與無脊椎動物類群和密度具有顯著相關性,而植物物種豐富度和地上生物量與無脊椎動物類群和密度不具有顯著相關性。
