茶樹CsGPX基因全基因組鑒定和表達分析
2019-07-23 07:37:55王贊曹紅利岳川郭雅玲
熱帶作物學報 2019年6期
王贊 曹紅利 岳川 郭雅玲
摘 ?要 ?為明確茶樹谷胱甘肽過氧化物酶(CsGPX)基因與非生物脅迫的關系,本研究利用生物信息學手段在茶樹基因組中篩選得到3條CsGPX基因,對其編碼蛋白理化性質、基因結構、系統進化樹、順式作用元件進行了分析。結果表明,CsGPX包含完整GPX結構域和3段保守序列,都具6個外顯子。預測啟動子上有參與植物激素和非生物脅迫響應的順式作用元件。使用實時熒光定量PCR測定該基因的表達譜,發現它們在不同組織中的表達有顯著差異。進一步分析表明,CsGPX被低溫、ABA、鹽以及干旱處理上調表達,但CsGPX2和CsGPX3的表達受低溫抑制。本研究為茶樹CsGPX家族基因克隆和功能驗證提供基礎。
關鍵詞 ?茶樹;谷胱甘肽過氧化物酶(GPX);非生物脅迫
中圖分類號 ?S571.1 ?????文獻標識碼 ?A
Abstract ?To reveal the possible involvement of glutathione peroxidase genes (CsGPX) in the responses of tea plants to abiotic stresses, three CsGPX genes were identified by bioinformatics tools from the tea genome data, and the gene structures, the cis-elements on their promoters, the physiochemical characteristics of the encoded proteins and the phylogenetic trees were also studied. The results showed that all the CsGPX genes contained 6 exons and shared the gene family-specific GPX domain in addition to three other conserved domains. Cis-elements for the responses of plants to hormones and abiotic stresses were identified on the promoters. Quantitative real-time PCR analysis of the expression profiles revealed that they were significantly different in expression in different tissues. Further analysis showed that the three CsGPX genes were, in general, up-regulated by treatments of low temperature, ABA, salt and drought except that CsGPX2 and CsGPX3 were down-regulated by treatment of low-temperature. The results should have laid down a basis for further characterization of CsGPX family genes in tea plant.
Keywords ?tea plant (Camellia sinensis); glutathione peroxidase (GPX); abiotic stress
DOI ?10.3969/j.issn.1000-2561.2019.06.014
生長環境造成的非生物脅迫是農作物減產、品質下降的重要因素[1]。植物在低溫、干旱、高鹽、重金屬和農藥等非生物脅迫下產生大量活性氧(reactive oxygen species, ROS)[2-4],導致脂質過氧化、DNA和蛋白質氧化以及光合器官的損傷[5]。而酶促和非酶促抗氧化防御機制是調節植物體內ROS穩態,提高抗氧化能力的重要途徑[6-7]。
谷胱甘肽過氧化物酶(glutathione peroxidase, GPXs)作為重要的酶促抗氧化劑,依賴于硫氧還蛋白(thioredoxin, Trx)還原分解H2O2,避免植物細胞氧化損傷,提高植物的抗逆性[8-10]。已經在擬南芥[11]、水稻[12]、棉花[13]等植物中鑒定得到谷胱甘肽過氧化物酶基因家族,也驗證其在非生物脅迫氧化應激響應中的重要性。Martinez等[14]檢測番茄葉綠素熒光參數發現,LeGPX表達增加了對高鹽和熱脅迫抗性,保護光反應系統功能。大豆GPX減少脂質過氧化對細胞的損傷,緩解干旱脅迫帶來的危害[15]。轉錄組分析表明擬南芥AtGPX參與了低溫脅迫導致的氧化應激響應,通過維持ROS平衡而減少氧化損害[16]。Wang等[17]發現水稻OsGPX突變體萌發活力低,對脅迫敏感并且發育不良。Zhou等[18]利用蛋白質組學方法發現非生物脅迫導致ROS積累,而GPX等抗氧化酶活性的增加則有利于植物對抗ROS脅迫。
隨著茶樹栽培面積的擴大,由環境脅迫導致的茶芽萌發遲、長勢差,營養積累困難等問題同步增長,對茶產量和品質的影響更加嚴重[19-21]。雖然植物GPX基因在脅迫氧化應激響應中十分重要,但茶樹CsGPX在多種非生物脅迫下的表達模式的報道還少。茶樹基因組測序的完成使挖掘功能基因資源成為可能[22-23]。本研究通過生物信息學方法,基于已公開的基因組數據,對茶樹CsGPX家族基因進行了全基因組鑒定,并分析了它們在非生物脅迫下的表達模式,為CsGPX基因功能的進一步研究打下初步基礎。
1 ?材料與方法
1.1 ?植物材料與非生物脅迫處理
2018年5月中旬,在福建農林大學園藝學院教學實驗基地采取以下供試材料:
(2)按照Yue等[24]的方法,選取2年生、健壯的盆栽鐵觀音茶苗,分別進行如下處理:通過葉片噴灑濃度為100 μmol/L ABA溶液;將植株移入并保持在4?℃人工氣候室中;將茶苗從盆中小心取出,用清水洗凈根部泥土后放置在10% (w/V) PEG-6000溶液中;用250 mmol/L NaCl溶液灌溉。所有處理分別在0、3、9、24 h取樣,取頂端第2和第3片成熟葉片,用錫箔紙分裝后迅速投入液氮速凍,然后移至?80?℃保存備用。各樣品均設置3次生物學重復。
1.2 ?方法
1.2.1 ?茶樹基因組CsGPX基因鑒定方法 ?從Pfam數據庫(http://pfam.xfam.org)下載GPX基因典型保守結構域隱馬可夫模型HMM(Hidden Markov Modelle)文件Pf00255[8],利用HMMER3.0軟件分別在中國科學院昆明植物研究所公布的阿薩姆種[Camellia sinensis var. assamica (CSA; Assam type)]茶樹基因組數據庫中(http://www.plantkingdomgdb.com/tea_tree/)進行比對搜索[22]。同時從TAIR數據庫(https://www. arabidopsis.org/)下載擬南芥AtGPX蛋白序列,作為檢索序列使用本地BLAST在茶樹基因組數據庫中檢索候選蛋白(E-value≤10?10)。利用Pfam和SMART(http://smart.embl-heidelberg.de/)檢測候選蛋白結構域,獲得CsGPX蛋白序列。
2 ?結果與分析
2.1 ?茶樹CsGPX基因鑒定
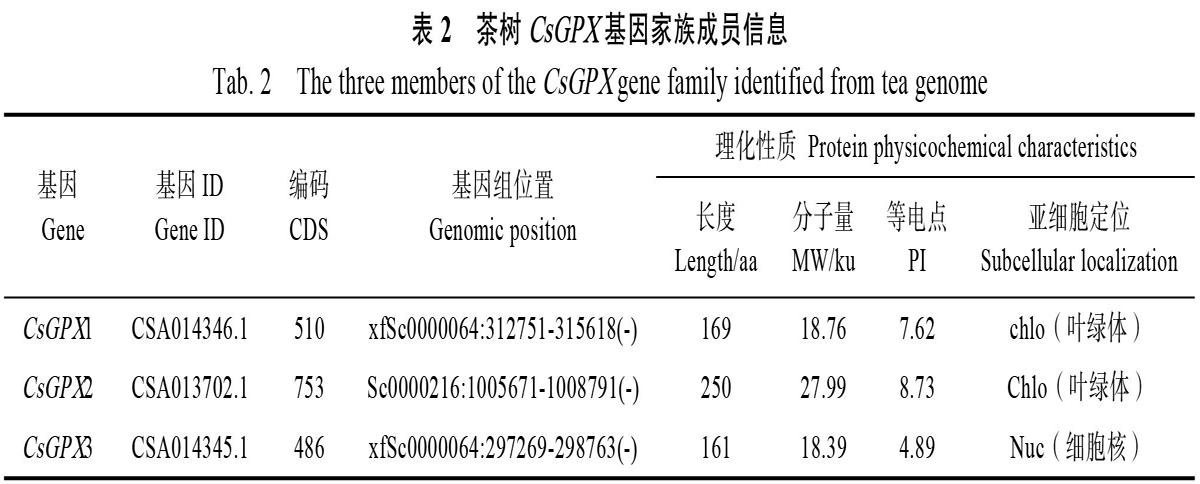
利用HMMER 3.0軟件和本地BLAST在茶樹基因組數據庫中檢索出候選蛋白,然后通過Pfam、SMART和DNAMAN比對,確定包含完整GPX結構域以及3個高度保守域(圖1A)的CsGPX基因,并依次命名為CsGPX1、CsGPX2和CsGPX3。生物信息學分析表明,CsGPX1編碼
169氨基酸殘基,分子量為18.76 ku。CsGPX2編碼250氨基酸殘基,分子量為27.99 ku。CsGPX3編碼161氨基酸殘基,分子量為18.39 ku(表2)。
2.2 ?茶樹CsGPX蛋白保守結構域和進化分析
利用Pfam和SMART對CsGPX蛋白序列檢索發現,這3條CsGPX包含了完整的GSHPx結構域,還包含了GPX家族的PLN02399和GSH_Peroxidase結構域(圖1C),主要集中在C端。多序列比對發現,CsGPX含有3段高度保守序列(圖1A),分別為GKVLLIVNVASXCG(GPX signature 1),ILAFPCNQ(GPX signature 2)和WNFXKF[27],還有3個半胱氨酸殘基(Cys)以及組成預測的催化活性位點的氨基酸殘基。
從Phytozome 12數據庫其他12個物種的GPX蛋白序列,采用N-J法與擬南芥和茶樹CsGPX構建系統進化樹,bootstrap檢驗采用1000次重復。結果顯示,CsGPX1、CsGPX2與可可TcGPX6、桃PpGPX6和擬南芥AtGPX6聚為一類,CsGPX3與西紅柿SiGPX8、蕓薹BrGPX8和擬南芥AtGPX8聚在一起(圖1B)。
2.3 茶樹CsGPX保守元件和基因結構分析
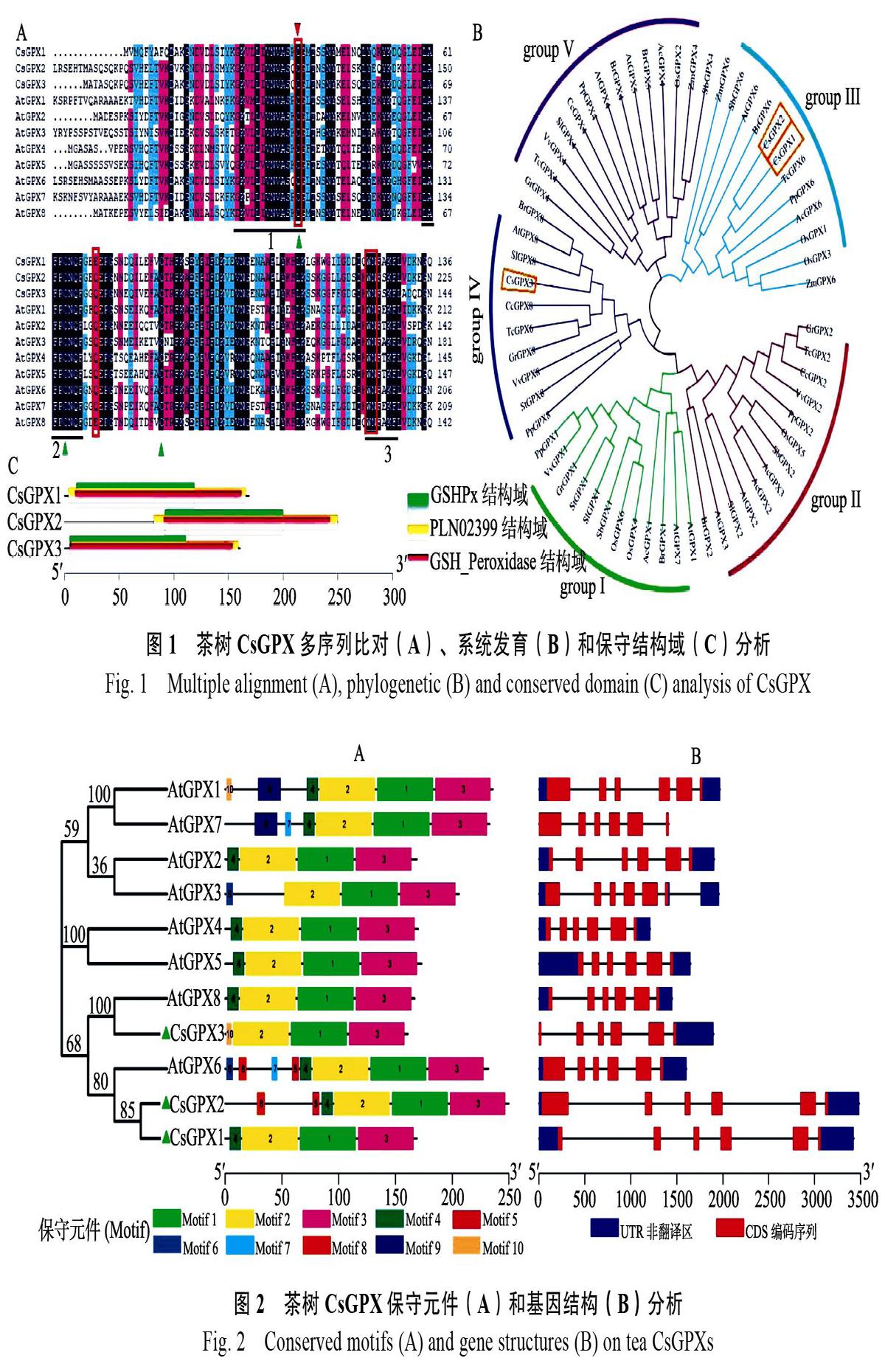
利用MEME在線軟件對CsGPX和擬南芥AtGPX蛋白序列分析發現(圖2A),所有序列都包含第1、2、3、4總共4個Motif。其中Motif 1、Motif 2和Motif 3組成了CsGPX蛋白GSHPx結構域,Motif 1、Motif 2、Motif 3和Motif 4則構成PLN02399和GSH_Peroxidase結構域,是CsGPX蛋白主要功能區域。
將CsGPX基因CDS和其對應的基因組序列通過GSDS在線分析,進一步了解CsGPX基因結構。結果顯示,這些基因結構相似,都包括5個內含子和6個外顯子(圖2B),與Zhai等人[28]的研究結果一致。
下劃線分別表示3個保守結構區域,紅色三角形表示哺乳動物中被Sec取代的半胱氨酸殘基。綠色三角表示植物GPX蛋白的3個半胱氨酸殘基,紅色方框表示組成GPX催化部位的氨基酸殘基(Cys、Gln、Trp和Asn)。AcGPX:菠蘿Ananas comosus;SbGPX:高粱Sorghum bicolor;TcGPX:可可Theobroma cacao;CcGPX:克萊門柚Citrus clementina;StGPX:馬鈴薯Solanum tuberosum;GrGPX:棉花Gossypium raimondii;AtGPX:擬南芥Arabidopsis thaliana;VvGPX:葡萄Vitis vinifera;OsGPX:水稻Oryza sativa;PpGPX:桃Prunus persica;SlGPX:西紅柿Solanum lycopersicum;ZmGPX:玉米Zea mays;BrGPX:蕓薹Brassica rapa。
2.4 ?茶樹CsGPX基因順式作用元件分析
將每條CsGPX起始密碼子ATG上游2000 bp作為啟動子區,在線軟件PlantCARE預測了該區域順式作用元件。結果顯示(表3),3條CsGPX具有的順式作用元件主要分為植物激素響應元件,如脫落酸響應元件(ABRE)、乙烯響應元件(ERE)、生長素響應元件(AuxRR-core)、茉莉酸甲酯響應元件(CGTCA-motif)、赤霉素響應元件(GARE-motif)等。植物生長發育相關順式作用元件,如O2-site、Skn-1 motif和circadian。生物和非生物脅迫響應順式作用元件,如熱脅迫響應元件(HSE)、干旱誘導元件(MBS)、逆境防御響應元件(TC-rich)、真菌誘導子響應元件(Box-W1)。預測結果表明茶樹CsGPX在調節生長發育、激素信號和應對環境脅迫方面有重要作用。
2.5 ?茶樹CsGPX基因表達分析
通過實時熒光定量PCR分析了CsGPX在鐵觀音不同組織中的表達,結果顯示CsGPX1在根中的表達水平遠高地上部組織,而CsGPX2則是地上部的表達量高于地下部根系,CsGPX3則在莖、葉中有較高的表達量(圖3)。
茶樹CsGPX在ABA、干旱(PEG)、低溫(4?℃)和高鹽(NaCl)脅迫下的表達譜見圖4,各處理下CsGPX表達的總體趨勢是上調,其中ABA誘導CsGPX上調表達的速度快于PEG和NaC。低溫處理只對CsGPX1的表達有明顯的誘導作用。不同小寫字母表示顯著差異(P<0.05)。
3 ?討論
本研究對茶樹全基因組范圍內進行了CsGPX基因鑒定,共得到3個CsGPX家族基因。生物信息學分析顯示均含有完整GPX保守結構域,以及3段保守蛋白序列GKVLLIVNVASXCG(GPX signature 1),ILAFPCNQ(GPX signature 2)和WNFXKF[29]。植物GPX活性依賴3個保守的半胱氨酸(Cys)殘基的前2個[30-31],這與動物的不同,動物GPX的催化活性位點依賴含硒代半胱氨酸(SeCys)殘基。此外,茶樹CsGPX至少包含4個Motif,構成了CsGPX蛋白主要保守功能區域[32],這也不同于動物GPX。啟動子區預測結果顯示,茶樹CsGPX的啟動子上主要有植物激素、生物和非生物脅迫等響應順式作用元件,暗示它們和其他植物中GPX基因一樣會受到多種植物激素的調控[33-35]。
植物中GPX的組織特異性表達分析顯示,作為葉綠體蛋白的擬南芥AtGPX1通常在綠色組織中大量表達[31],荷花LjGPX家族成員在不同組織中的表達水平同樣差異顯著[27]。本研究結果表明根是CsGPX1主要表達部位,CsGPX2在莖、葉、花和果實中的表達量均較高,CsGPX3則是主要在莖、葉中表達。有研究認為抗氧化基因的組織差異性表達模式有利于維持不同組織中的ROS穩態,有利于抗逆性的提高[36]。
作為抗氧化代謝網絡中的關鍵酶,GPX維持細胞ROS水平從而保護細胞膜結構完整,提高植物對脅迫氧化應激的耐受程度[37-38]。過表達擬南芥AtPGX3不僅能夠調控H2O2穩態,并且通過Ca2+通道調節氣孔參與到ABA代謝途徑,提高對干旱脅迫耐受性[35]。在冷害脅迫下,水稻OsGPX參與了根部ROS清除,OsGPX高表達及其酶活性的增加提高了水稻的冷害耐受性[39-40]。Gao等[33]采用蛋白免疫印跡和熒光定量PCR技術發現高鹽脅迫在根和葉中調節鹽芥TsGPX表達,TsGPX家族成員中TsGPX7對于抵御高鹽脅迫最為重要。非生物脅迫下的CsGPX表達譜說明其參與茶樹的氧化應激防御機制,本研究結果與已有報道結果相似[8, 12],干旱(PEG)、低溫(4?℃)和高鹽(NaCl)脅迫均能誘導提高茶樹葉片中不同CsGPX的表達水平,盡管不同的CsGPX對不同的處理的響應程度有明顯的差異。
ABA信號傳導途徑將外界環境的脅迫壓力通過一系列的ABA受體傳遞到下游代謝通道,進而調控抗性基因表達來增強植物耐受性[41]。Dong等[42]對ABA合成突變體擬南芥研究發現,ABA介導的信號網絡增強了酶催活性氧清除劑的基因轉錄水平,提高了耐鹽性。本研究中,CsGPX能迅速響應ABA處理,3 h內表達量就提高了3倍,并在處理的中后期保持較高水平。結合啟動子序列分析,3條CsGPX均存在響應ABA的順式作用元件ABRE,使得ABA可以調控CsGPX表達。通過水稻抗寒性轉錄分析揭示,在低溫脅迫下OsGPX表達量被上調,谷胱甘肽過氧化物酶是主要的抗氧化劑,ABA信號轉導途徑相關基因參與了冷應激反應,并且ABA信號通路起著主導作用[40]。Gaber[43]發現ABA作為脅迫響應基因誘導物可調節擬南芥AtGPX家族基因表達,通過在擬南芥中異源表達小麥TaGPX發現,其在非生物脅迫和ABA信號傳導中有重要作用[28]。
茶樹基因組數據的公布有利于從全基因組水平上對重要功能基因進行篩選,其結果更為全面。本研究利用生物信息學手段,在茶樹全基因組中鑒定的到3條CsGPX基因家族成員,分析其在不同非生物脅迫中的表達模式。明確了該家族基因在茶樹逆境氧化應激響應中具有重要功能,豐富了茶樹抗逆機理研究,為進一步在模式植物中對CsGPX進行功能驗證和優良品種選育提供理論基礎。
參考文獻
Hu H, Xiong L. Genetic engineering and breeding of drought-resistant crops[J]. Annual Review of Plant Biology 2014, 65: 715-741.
Shakir S K, Irfan S, Akhtar B, et al. Pesticide-induced oxidative stress and antioxidant responses in tomato (Solanum lycopersicum) seedlings[J]. Ecotoxicology, 2018(9): 1-17.
Kumar D, Yusuf M A, Singh P, et al. Modulation of antioxidant machinery in -tocopherol-enriched transgenic Brassica juncea plants tolerant to abiotic stress conditions[J]. Protoplasma, 2013, 250(5): 1079-1089.
Manquián-Cerda K, Cruces E, Escudey M, et al. Interactive effects of aluminum and cadmium on phenolic compounds, antioxidant enzyme activity and oxidative stress in blueberry (Vaccinium corymbosum L.) plantlets cultivated in vitro[J]. Ecotoxicology and Environmental Safety, 2018, 150: 320- 326.
Mates J M. Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology[J]. Toxicology, 2000, 153(1-3): 83-104.
Gill S S, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants[J]. Plant Physiology and Biochemistry, 2010, 48(12): 909-930.
You J, Chan Z. ROS regulation during abiotic stress responses in crop plants[J]. Front Plant Science, 2015, 6: 1092.
Zhou Y, Hu L, Ye S, et al. Genome-wide identification of glutathione peroxidase (GPX) gene family and their ?response to abiotic stress in cucumber[J]. 3 Biotech, 2018, 8(3): 159.
Herbette S, Menn A L, Rousselle P, et al. Modification of photosynthetic regulation in tomato overexpressing glutathione peroxidase[J]. Biochimica et Biophysica Acta (BBA) - General Subjects, 2005, 1724(1-2): 108-118.
Iqbal A, Yabuta Y, Takeda T, et al. Hydroperoxide reduction by thioredoxin-specific glutathione peroxidase isoenzymes of Arabidopsis thaliana[J]. The Federation of European Biochemical Societies Journal, 2006, 273(24): 5589- 5597.
Milla M A R, Maurer A, Huete A R, et al. Glutathione peroxidase genes in Arabidopsis are ubiquitous and regulated by abiotic stresses through diverse signaling pathways[J]. The Plant Journal, 2003, 36(5): 602-615.
Islam T, Manna M, Kaul T, et al. Genome-wide dissection of Arabidopsis and rice for the identification and expression analysis of glutathione peroxidases reveals their stress-specific and overlapping response patterns[J]. Plant Molecular Biology Reporter, 2015, 33(5): 1413-1427.
Chen M, Li K, Li H, et al. The glutathione peroxidase gene family in Gossypium hirsutum: genome-wide identification, classification, gene expression and functional analysis[J]. Scientific Reports, 2017, 7: 44743.
Martinez V, Nieves-Cordones M, Lopez-Delacalle M, et al. Tolerance to stress combination in tomato plants: New insights in the protective role of melatonin[J]. Molecules, 2018, 23(3): 535.
Xing X, Fang C, Li L, et al. Improved drought tolerance by α-naphthaleneacetic acid-induced ROS accumulation in two soybean cultivars[J]. Journal of Integrative Agriculture, 2016, 15(8): 1770-1784.
Ren L, Zhang D, Chen G, et al. Transcriptomic profiling revealed the regulatory mechanism of Arabidopsis seedlings response to oxidative stress from cryopreservation[J]. Plant Cell Reports, 2015, 34(12): 2161-2178.
Wang X, Fang G, Yang J, et al. A thioredoxin-dependent glutathione peroxidase (OsGPX5) is required for rice normal development and salt stress tolerance[J]. Plant Molecular Biology Reporter, 2017, 35(3): 333-342.
Zhou X, Chen S, Wu H, et al. Biochemical and proteomics analyses of antioxidant enzymes reveal the potential stress tolerance in Rhododendron chrysanthum Pall[J]. Biology Direct, 2017, 12: 10.
Liu S C, Jin J Q, Ma J Q, et al. Transcriptomic analysis of tea plant responding to drought stress and recovery[J]. PLoS ONE, 2016, 11(1): e0147306.
Hao X, Yang Y, Yue C, et al. Comprehensive transcriptome analyses reveal differential gene expression profiles of Camellia sinensis axillary buds at para-, endo-, ecodormancy, and bud flush stages[J]. Frontiers in Plant Science, 2017, 8: 553.
Cheruiyot E K, Mumera L M, Ng'etich W K, et al. High fertilizer rates increase susceptibility of tea to water stress[J]. Journal of Plant Nutrition, 2009, 33(1): 115-129.
Xia E, Zhang H, Sheng J, et al. The tea tree genome provides insights into tea flavor and independent evolution of caffeine biosynthesis[J]. Molecular Plant, 2017, 10(6): 866-877.
Wei C L, Yang H, Wang S, et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality[J]. Proceedings of the National Academy of Sciences, 2018, 115(18): E4151-E4158.
Yue C, Cao H, Wang L, et al. Molecular cloning and expression analysis of tea plant aquaporin (AQP) gene family[J]. Plant Physiology and Biochemistry, 2014, 83: 65-76.
Hao X, Horvath D, Chao W, et al. Identification and evaluation of reliable reference genes for quantitative real-time PCR analysis in tea plant (Camellia sinensis (L.) O. Kuntze)[J]. International Journal of Molecular Sciences, 2014, 15(12): 22155-22172.
Cao H, Wang L, Yue C, et al. Isolation and expression analysis of 18 CsbZIP genes implicated in abiotic stress responses in the tea plant (Camellia sinensis)[J]. Plant Physiology and Biochemistry, 2015, 97: 432-442.
Ramos J, Matamoros M A, Naya L, et al. The glutathione peroxidase gene family of Lotus japonicus: characterization of genomic clones, expression analyses and immunolocalization in legumes[J]. The New Phytologist 2009, 181(1): 103-114.
Zhai C, Zhao L, Yin L, et al. Two wheat glutathione peroxidase genes whose products are located in chloroplasts improve salt and H2O2 tolerances in Arabidopsis[J]. PLoS One, 2013, 8(10): e73989.
Liu W, Zhao C, Wang P, et al. The response of glutathione peroxidase 1 and glutathione peroxidase 7 under different oxidative stresses in black tiger shrimp, Penaeus monodon[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2018, 217: 1-13.
Attacha S, Solbach D, Bela K, et al. Glutathione peroxidase-like enzymes cover five distinct cell compartments and membrane surfaces in Arabidopsis thaliana[J]. Plant, Cell & Environment, 2017, 40(8): 1281-1295.
Navrot N, Collin V, Gualberto J, et al. Plant glutathione peroxidases are functional peroxiredoxins distributed in several subcellular compartments and regulated during biotic and abiotic stresses[J]. Plant Physiology, 2006, 142(4): 1364-1379.
Kim Y, Jang M, Noh H, et al. Molecular characterization of two glutathione peroxidase genes of Panax ginseng and their expression analysis against environmental stresses[J]. Gene, 2014, 535(1): 33-41.
Gao F, Chen J, Ma T, et al. The glutathione peroxidase gene family in Thellungiella salsuginea: genome-wide identification, classification, and gene and protein expression analysis under stress conditions[J]. International Journal of Molecular Sciences, 2014, 15(2): 3319-3335.
Passaia G, Queval G, Bai J, et al. The effects of redox controls mediated by glutathione peroxidases on root architecture in Arabidopsis thaliana[J]. Journal of Experimental Botany, 2014, 65(5): 1403-1413.
Miao Y, Lv D, Wang P, et al. An Arabidopsis glutathione peroxidase functions as both a redox transducer and a scavenger in abscisic acid and drought stress responses[J]. The Plant Cell, 2006, 18(10): 2749-2766.
Yu L, Ma J, Niu Z, et al. Tissue-specific transcriptome analysis reveals multiple responses to salt stress in populus euphratica seedlings[J]. Genes, 2017, 8(12): 372.
Islam T, Manna M, Reddy M K. Glutathione peroxidase of pennisetum glaucum (PgGPx) Is a functional Cd2+ dependent peroxiredoxin that enhances tolerance against salinity and drought stress[J]. PLoS One, 2015, 10(11): e143344.
Koua D, Cerutti L, Falquet L, et al. PeroxiBase: a database with new tools for peroxidase family classification[J/OL]. Nucleic Acids Research, 2009, 37(suppl_1): D261-D266. [2018-10-11]. https://doi.org/10.1093/nar/gkn680
Yoo Y, Anil Kumar N C, Park J, et al. Global analysis of differentially expressed genes between japonica and indica rice roots reveals the molecular basis for enhanced cold tolerance in japonica rice[J]. Plant Biotechnology Reports, 2017, 11(6): 461-473.
Zhao J, Zhang S, Yang T, et al. Global transcriptional profiling of a cold-tolerant rice variety under moderate cold stress reveals different cold stress response mechanisms[J]. Physiologia Plantarum, 2015, 154(3): 381-394.
Lim C W, Baek W, Jung J, et al. Function of ABA in stomatal defense against biotic and drought stresses[J]. International Journal of Molecular Sciences, 2015, 16(12): 15251-15270.
Dong W, Wang M, Xu F, et al. Wheat oxophytodienoate reductase gene TaOPR1 confers salinity tolerance via enhancement of abscisic acid signaling and reactive oxygen species scavenging[J]. Plant Physiology 2013, 161(3): 1217-1228.
Gaber A. The importance of Arabidopsis glutathione peroxidase 8 for protecting Arabidopsis plant and E. coli cells against oxidative stress[J]. GM Crops & Food, 2014, 5(1): 20-26.
