組成型過(guò)表達(dá)ido基因?qū)?-羥基異亮氨酸合成的影響及其發(fā)酵培養(yǎng)基的響應(yīng)面優(yōu)化
2019-07-26 08:24:50何繼龍李英滋韓世寶滿德恩郭脈海戰(zhàn)俊杰張成林
食品科學(xué) 2019年14期
關(guān)鍵詞:產(chǎn)量
何繼龍,李英滋,韓世寶,滿德恩,郭脈海,戰(zhàn)俊杰,張成林,,*
(1.天津科技大學(xué)生物工程學(xué)院,天津 300457;2.菱花集團(tuán)有限公司,山東 濟(jì)寧 272073)
L-異亮氨酸羥化物——4-羥基異亮氨酸具有血糖濃度依賴(lài)的促進(jìn)胰島素分泌特性,以及促進(jìn)脂肪代謝等功能,對(duì)糖尿病具有良好的治療效果[1-4]。同時(shí),4-羥基異亮氨酸還具有降血脂、保護(hù)肝功能等作用[5-7],具有廣闊的應(yīng)用前景。目前4-羥基異亮氨酸的工業(yè)化生產(chǎn)主要采用胡蘆巴種子提取法,然而此方法生產(chǎn)效率低(1 kg胡蘆巴種子僅能提取出150 mg 4-羥基異亮氨酸),致使生產(chǎn)規(guī)模小、產(chǎn)量低,嚴(yán)重限制了其應(yīng)用[8]。此外,化學(xué)合成法因在生產(chǎn)過(guò)程中使用會(huì)產(chǎn)生乙醛等有毒或易爆物且存在提取收率低等不足,仍停留在研究階段[9-13]。Kodera[13]、Ogawa[14]等在蘇云金芽孢桿菌(Bacillus thuringiensis)中發(fā)現(xiàn)異亮氨酸羥化酶(isoleucine dioxygenase,IDO),由ido基因編碼,該酶能夠以L-異亮氨酸和α-酮戊二酸為底物特異性催化生成4-羥基異亮氨酸(圖1)。在前期研究中,利用該酶開(kāi)發(fā)出靜息細(xì)胞法合成4-羥基異亮氨酸,但底物L(fēng)-異亮氨酸和α-酮戊二酸價(jià)格較高致使該方法不利用工業(yè)化生產(chǎn)[15]。Smirnov[12]、Kivero[16]等將異亮氨酸羥化酶編碼基因ido轉(zhuǎn)化至大腸桿菌并在發(fā)酵過(guò)程中添加L-異亮氨酸,實(shí)現(xiàn)前體物添加法合成4-羥基異亮氨酸,然而該方法存在L-異亮氨酸被額外消耗及耗糖高等不足。
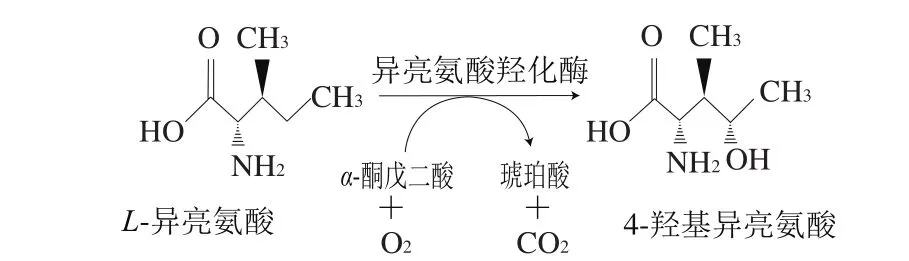
圖1 異亮氨酸羥化酶催化生成4-羥基異亮氨酸的反應(yīng)示意圖Fig. 1 Reaction route for IDO-catalyzed synthesis of 4-hydroxyisoleucine
前期研究從蘇云金芽孢桿菌TCCC 11826中克隆出ido基因(Accession No. KC884243)[17],利用該基因以L-異亮氨酸生產(chǎn)菌Corynebacterium glutamicum YILW-2為出發(fā)菌株[18],構(gòu)建了4-羥基異亮氨酸生產(chǎn)菌HIL016,實(shí)現(xiàn)發(fā)酵法合成4-羥基異亮氨酸。在HIL016菌株中,采用誘導(dǎo)型質(zhì)粒pXMJ19過(guò)表達(dá)ido基因,然而誘導(dǎo)劑IPTG通常對(duì)菌體細(xì)胞生長(zhǎng)存在抑制作用且因其價(jià)格高昂不利于工業(yè)化生產(chǎn)。針對(duì)該問(wèn)題本研究構(gòu)建組成型表達(dá)質(zhì)粒并考察ido基因組成型過(guò)表達(dá)對(duì)4-羥基異亮氨酸合成的影響,在此基礎(chǔ)上利用響應(yīng)面法優(yōu)化該菌株搖瓶發(fā)酵培養(yǎng)基。
1 材料與方法
1.1 材料與試劑
1.1.1 菌株和質(zhì)粒
本研究所用4-羥基異亮氨酸生產(chǎn)菌株谷氨酸棒桿菌HIL016(C. glutamicum YILW-2ΔaceAPppc::PtufPgltA::Ptuf/pXMJ19-ido)、大腸桿菌(Escherichia coli)DH5α以及質(zhì)粒pXMJ19和pXMJ19-ido均由本實(shí)驗(yàn)室保藏。
1.1.2 引物
本研究所用引物見(jiàn)表1。
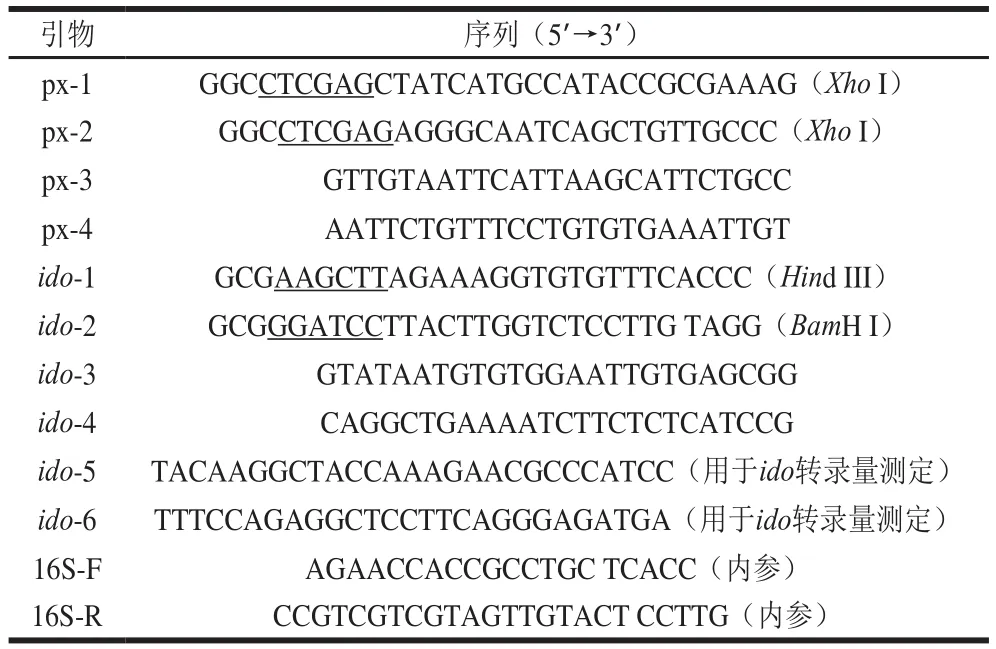
表1 實(shí)驗(yàn)所用引物Table 1 Primer sequences used in this study
1.1.3 培養(yǎng)基
LB培養(yǎng)基:胰化蛋白胨10 g/L,酵母提取物5 g/L,NaCl 10 g/L,pH 7.0~7.5,121 ℃高壓蒸汽滅菌20 min[19]。
LBG培養(yǎng)基:LB培養(yǎng)中添加5 g/L葡萄糖,pH 7.0~7.5,115 ℃高溫蒸汽滅菌15 min。
種子培養(yǎng)基:葡萄糖25 g/L,酵母粉5 g/L,(NH4)2SO45 g/L,KH2PO4·3H2O 2 g/L,MnSO4·7H2O 0.6 g/L,玉米漿40 mL/L,豆餅水解液 15 mL/L,pH 6.7~7.0,115 ℃高壓蒸汽滅菌15 min。
發(fā)酵培養(yǎng)基:葡萄糖100 g/L,(NH4)2SO43 g/L,KH2PO4·3H2O 1.5 g/L,MgSO4·7H2O 0.6 g/L,MnSO4·7H2O 0.015 g/L,F(xiàn)eSO4·7H2O 0.015 g/L,VB10.001 g/L,谷氨酸3 g/L,酵母粉0.5 g/L,玉米漿30 mL/L,pH 6.7~7.0,115 ℃高壓蒸汽滅菌15 min。
1.1.4 試劑
限制性內(nèi)切酶、T4 DNA連接酶、ExTaq DNA聚合酶、RNAiso Plus、primeScriptTMRT、SYBR?Premix Ex Taq? II等 寶生物工程(大連)有限公司;4-羥基異亮氨酸標(biāo)準(zhǔn)品 美國(guó)Sigma公司;化學(xué)試劑均購(gòu)于國(guó)藥集團(tuán)化學(xué)試劑有限公司。
1.2 儀器與設(shè)備
U3000高效液相色譜儀 美國(guó)賽默飛世爾科技公司;PTC-1148型聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction,PCR)儀 美國(guó)Bio-Rad公司;7500實(shí)時(shí)熒光定量PCR儀 美國(guó)ABI公司;PD-10除鹽柱 英國(guó)GE Healthcare公司;ZORBAX Eclipse AAA氨基酸柱美國(guó)Agilent公司。
1.3 方法
1.3.1 組成型表達(dá)質(zhì)粒pXM01的構(gòu)建
根據(jù)質(zhì)粒pXMJ19中阻遏蛋白編碼基因lacI上下游序列設(shè)計(jì)引物px-1和px-2(表1)[20]。以質(zhì)粒pXMJ19為模板,利用引物px-1和px-2進(jìn)行PCR擴(kuò)增,擴(kuò)增產(chǎn)物經(jīng)純化后利用Xho I酶切并經(jīng)瓊脂糖凝膠電泳、切膠、回收。回收產(chǎn)物經(jīng)T4 DNA連接酶連接后轉(zhuǎn)化至E. coli DH5α感受態(tài)細(xì)胞中,然后涂布于含30 μg/mL氯霉素的LB固體培養(yǎng)基,于37 ℃倒置培養(yǎng)。挑取轉(zhuǎn)化子單菌落,利用引物px-3和px-4進(jìn)行菌落PCR鑒定,將鑒定正確的質(zhì)粒命名為pXM01。
1.3.2 ido基因組成型表達(dá)質(zhì)粒pXM01-ido及菌株HIL017的構(gòu)建
以質(zhì)粒pXMJ19-ido為模板,利用引物ido-1和ido-2擴(kuò)增ido基因,擴(kuò)增產(chǎn)物經(jīng)純化后利用BamH I和Hind III雙酶切,經(jīng)瓊脂糖凝膠電泳、切膠、回收后連接至經(jīng)相同酶切的pXM01。將上述連接產(chǎn)物轉(zhuǎn)化至E. coli DH5α感受態(tài)細(xì)胞中,然后涂布于含30 μg/mL氯霉素的LB固體培養(yǎng)基,于37 ℃倒置培養(yǎng)。挑取轉(zhuǎn)化子單菌落,經(jīng)活化后提取質(zhì)粒,分別利用Hind III及BamH I和Hind III進(jìn)行單、雙酶切驗(yàn)證,并由蘇州金唯智生物科技有限公司測(cè)定質(zhì)粒中ido序列。將鑒定正確的質(zhì)粒命名為pXM01-ido。
將pXM01-ido轉(zhuǎn)化至不含質(zhì)粒pXMJ19-ido的HIL016感受態(tài)細(xì)胞(2.5 kV,25 μF,200 Ω,1 mm電擊杯),然后涂布于含30 μg/mL氯霉素的LBG固體培養(yǎng)基[21-22],于32 ℃倒置培養(yǎng)。挑取單菌落活化后提取其質(zhì)粒利用引物ido-3和ido-4進(jìn)行PCR鑒定,將鑒定正確的菌株命名為HIL017。
1.3.3 實(shí)時(shí)定量PCR
根據(jù)ido及谷氨酸棒桿菌C. glutamicum ATCC13032 16S rDNA(內(nèi)參)序列采用Primer5.0軟件設(shè)計(jì)用于實(shí)時(shí)定量PCR的引物(表1)。分別將HIL016和HIL017接種至LBG培養(yǎng)基(含30 μg/mL氯霉素),于32 ℃、220 r/min振蕩培養(yǎng)。對(duì)于HIL016,當(dāng)OD600nm為0.6~0.8時(shí)(2~3 h)添加終濃度為0.2 mmol/L的IPTG。分別于4、8、12、24 h和36 h收集培養(yǎng)物,按照說(shuō)明書(shū)提取菌體總RNA并反轉(zhuǎn)錄成cDNA。分別以接種至發(fā)酵培養(yǎng)基(0 h)的含質(zhì)粒pXMJ19的HIL016及含pXM01的HIL017細(xì)胞為對(duì)照。利用實(shí)時(shí)熒光定量PCR儀進(jìn)行檢測(cè)。采用2-ΔΔCt法對(duì)ido的轉(zhuǎn)錄量進(jìn)行分析[23]。
1.3.4 IDO酶活性測(cè)定
按1.3.3節(jié)方法收集培養(yǎng)4、8、12、24 h和36 h的HIL016和HIL017細(xì)胞,重懸于10 mL Tris-HCl緩沖液(100 mmol/L,pH 7.0),利用超聲破碎儀破碎細(xì)胞。將細(xì)胞破碎物于4 ℃、13 000×g離心30 min后取上清液,然后利用PD-10除鹽柱過(guò)濾除鹽。取上述濾液100 μL加入900 μL含10 mmol/L α-酮戊二酸、10 mmol/L L-異亮氨酸、5 mmol/L FeSO4·7H2O和10 mmol/L抗血酸的Tris-HCl緩沖液(100 mmol/L,pH 7.0),反應(yīng)30 min后測(cè)定利用高效液相色譜儀測(cè)定4-羥基異亮氨酸濃度[8],以每毫克總蛋白每分鐘催化生成的4-羥基異亮氨酸(nmol/(min·mg))表示IDO比活力。
1.3.5 Plackett-Burman(PB)試驗(yàn)設(shè)計(jì)
選取前期研究中對(duì)4-羥基異亮氨酸產(chǎn)量影響較大的9 個(gè)因素(N=9),每個(gè)因素分為高(1)和低(-1)2 個(gè)水平,進(jìn)行PB試驗(yàn)設(shè)計(jì)(表2)。按照設(shè)計(jì)方案進(jìn)行12 次試驗(yàn),響應(yīng)值為4-羥基異亮氨酸產(chǎn)量,篩選出對(duì)其有顯著影響的因素。
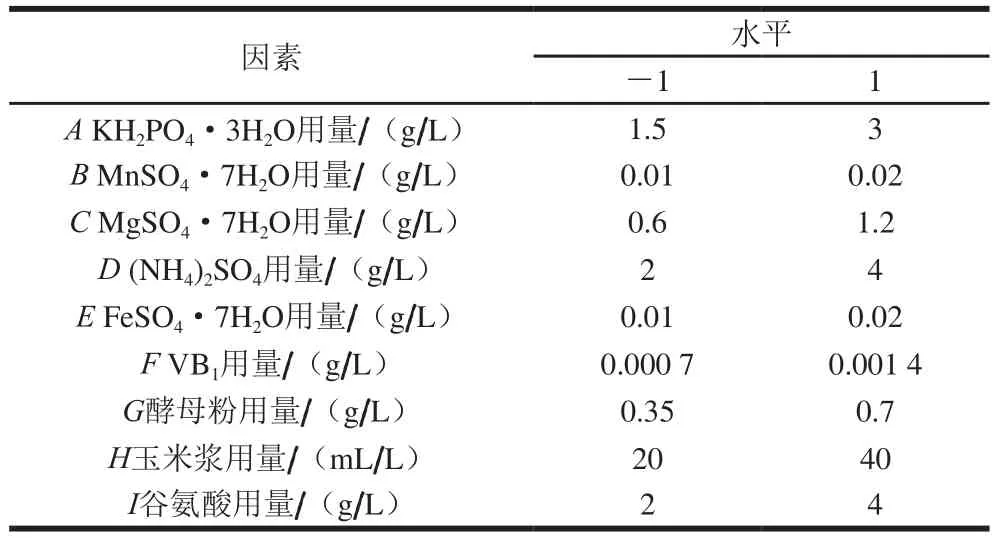
表2 PB試驗(yàn)設(shè)計(jì)因素與水平Table 2 Factors and levels used for PB experimental design
1.3.6 Box-Behnken試驗(yàn)設(shè)計(jì)
以4-羥基異亮氨酸產(chǎn)量為響應(yīng)值,利用軟件Design Experts V8.0.6,對(duì)1.3.5節(jié)篩選出的對(duì)4-羥基異亮氨酸產(chǎn)量影響較大的因素玉米漿、谷氨酸和FeSO4·7H2O用量,設(shè)計(jì)3因素3水平的響應(yīng)面試驗(yàn)(表3),按照設(shè)計(jì)方案共進(jìn)行15 次試驗(yàn)。

表3 Box-Behnken試驗(yàn)設(shè)計(jì)因素及水平Table 3 Factors and levels used for Box-Behnken design
1.3.7 搖瓶發(fā)酵實(shí)驗(yàn)
將活化后的4-羥基異亮氨酸生產(chǎn)菌株以10%(體積分?jǐn)?shù),下同)接種量接種至含30 mL種子培養(yǎng)基(含30 μg/mL氯霉素)的500 mL搖瓶中,于32 ℃、220 r/min振蕩培養(yǎng)10~12 h。將種子培養(yǎng)物以10%接種量接種至含30 mL發(fā)酵培養(yǎng)基(含30 μg/mL氯霉素)的500 mL搖瓶中,以苯酚紅作為pH值指示劑,用氨水維持pH 7.0~7.5,于32 ℃、220 r/min振蕩培養(yǎng)48 h[24]。對(duì)于HIL016,當(dāng)OD600nm為0.6~0.8時(shí)添加終濃度為0.2 mmol/L的IPTG。
1.3.8 菌株HIL017中pXM01-ido質(zhì)粒的穩(wěn)定性檢測(cè)
將菌株HIL017于LBG液體培養(yǎng)基傳代,每次傳代培養(yǎng)12 h。取不同傳代次數(shù)的培養(yǎng)物,適當(dāng)稀釋后涂布于LBG固體培養(yǎng)基,于32 ℃倒置培養(yǎng)。挑取100 個(gè)單菌落,分別接種至不含氯霉素和含30 μg/mL氯霉素的LBG固體培養(yǎng)基,于32 ℃倒置培養(yǎng)。按式(1)計(jì)算HIL017中pXM01-ido質(zhì)粒的穩(wěn)定性[25]:

挑取含氯霉素LBG固體培養(yǎng)基中的單菌落活化后按1.3.6節(jié)方法進(jìn)行搖瓶發(fā)酵實(shí)驗(yàn),測(cè)定其4-羥基異亮氨酸產(chǎn)量。
1.3.9 氨基酸及生物量檢測(cè)
發(fā)酵結(jié)束后,取1 mL發(fā)酵液,于4 ℃、8 000×g離心5 min后取上清液。經(jīng)2,4-二硝基氟苯衍生后利用高效液相色譜儀測(cè)定4-羥基異亮氨酸、L-異亮氨酸、L-纈氨酸和賴(lài)氨酸濃度。檢測(cè)條件為:ZORBAX Eclipse AAA氨基酸柱,50%乙腈-50 mmol/L醋酸銨二元梯度洗脫。發(fā)酵液經(jīng)離心后,用生理鹽水洗滌菌體沉淀3 次,后用適量生理鹽水重懸。利用分光光度計(jì)測(cè)定其發(fā)酵液OD600nm,根據(jù)公式(2)計(jì)算菌體生物量(以干質(zhì)量計(jì)):
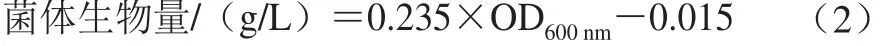
1.4 數(shù)據(jù)分析
每組實(shí)驗(yàn)均設(shè)置3 個(gè)平行并重復(fù)3 次,利用SPSS 13.0統(tǒng)計(jì)軟件對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行分析。
2 結(jié)果與分析
2.1 ido基因組成型表達(dá)質(zhì)粒及菌株的構(gòu)建
pXMJ19為谷氨酸棒桿菌常用表達(dá)質(zhì)粒,該質(zhì)粒含tac啟動(dòng)子和lacO元件,基因的轉(zhuǎn)錄受lacI編碼的LacI蛋白阻遏。按1.3.1節(jié)構(gòu)建lacI缺失的組成型表達(dá)質(zhì)粒pXM01。利用鑒定引物進(jìn)行PCR擴(kuò)增,結(jié)果如圖2A所示,PCR產(chǎn)物經(jīng)瓊脂糖凝膠電泳后,出現(xiàn)堿基數(shù)約為1 000 bp的條帶,與理論值(1 188 bp)接近;而以pXMJ19為模板擴(kuò)增出堿基數(shù)約為2 000 bp的條帶,與預(yù)期值(2 160 bp)接近,表明lacI成功從pXMJ19敲除。
將基因ido的PCR擴(kuò)增產(chǎn)物按1.3.2節(jié)方法連接至pXM01。將獲得的質(zhì)粒進(jìn)行酶切驗(yàn)證,結(jié)果如圖2B所示,質(zhì)粒經(jīng)單酶切獲得堿基數(shù)約為6 000 bp的條帶,與pXM01(5 635 bp)和ido(723 bp)堿基數(shù)之和接近;質(zhì)粒經(jīng)雙酶切獲得堿基數(shù)分別為5 500 bp和750 bp的條帶,分別與pXM01和ido堿基數(shù)接近,表明ido成功連接至pXM01,即pXM01-ido質(zhì)粒構(gòu)建成功。對(duì)該質(zhì)粒中ido基因測(cè)序發(fā)現(xiàn),其堿基序列未發(fā)生突變。
將重組質(zhì)粒pXM01-ido轉(zhuǎn)化至HIL016感受態(tài)細(xì)胞,提取轉(zhuǎn)化子質(zhì)粒進(jìn)行PCR鑒定,以不含ido基因的pXM01為對(duì)照。如圖2C所示,以從轉(zhuǎn)化子提取的重組質(zhì)粒為模板擴(kuò)增出堿基數(shù)約為850 bp的片段,而以pXM01為模板擴(kuò)增出堿基數(shù)約為150 bp的片段,與其理論值(分別為848 bp和155 bp)接近,表明pXM01-ido成功轉(zhuǎn)化至HIL016,即HIL017構(gòu)建成功。
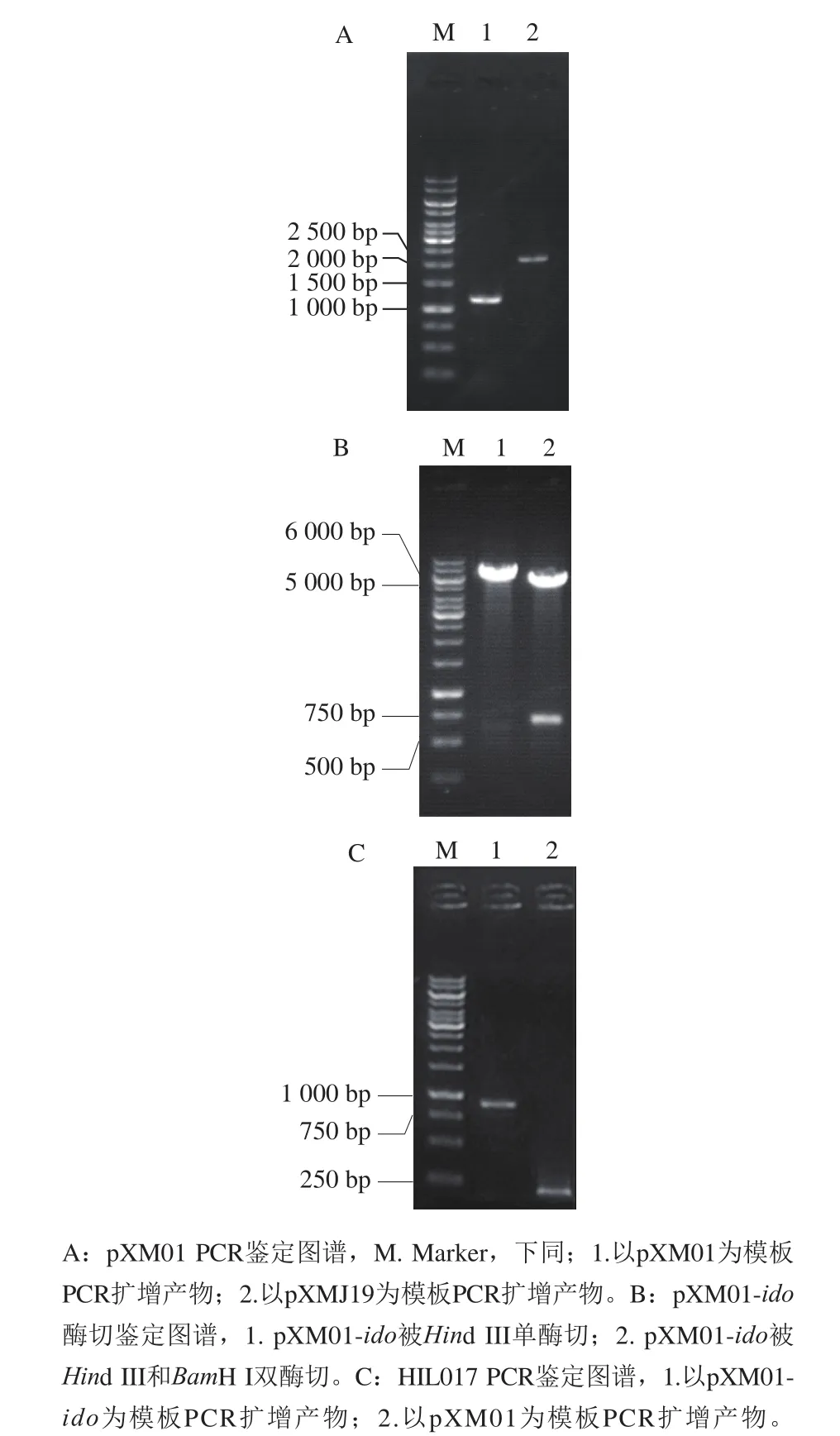
圖2 pXM01 PCR鑒定、pXM01-ido酶切鑒定及HIL017 PCR鑒定圖譜Fig. 2 Identification of pXM01, pXM01-ido and HIL017
2.2 菌株HIL016和HIL017的ido基因轉(zhuǎn)錄量及IDO比活力分析
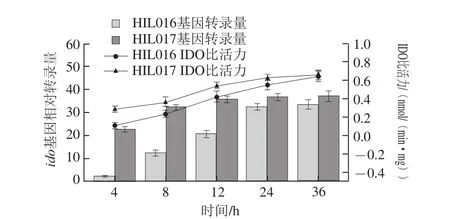
圖3 菌株HIL016和HIL017的ido相對(duì)轉(zhuǎn)錄量及IDO比活力Fig. 3 Transcription level of ido and specific activity of IDO in HIL016 and HIL017
如圖3所示,隨著培養(yǎng)時(shí)間的延長(zhǎng),菌株HIL016和HIL017的ido相對(duì)轉(zhuǎn)錄量及IDO比活力均逐漸提高,24 h后趨于穩(wěn)定。然而4~12 h時(shí),HIL017的ido相對(duì)轉(zhuǎn)錄量及IDO比活力均高于HIL016,24 h后二者無(wú)顯著差異。表明利用構(gòu)建的阻遏蛋白基因lacI敲除質(zhì)粒pXM01實(shí)現(xiàn)了ido基因的組成型表達(dá),且該質(zhì)粒更有益于發(fā)酵前期ido基因的過(guò)表達(dá)。
2.3 ido基因組成型過(guò)表達(dá)對(duì)4-羥基異亮氨酸合成的影響
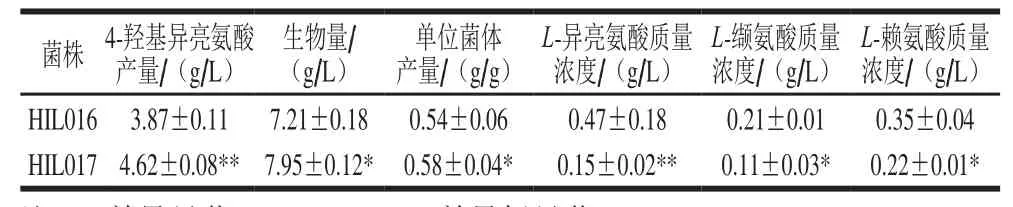
表4 菌株HIL016和HIL017搖瓶發(fā)酵主要參數(shù)Table 4 Shake-flask fermentation parameters for HIL016 and HIL017
為考察ido基因組成型過(guò)表達(dá)對(duì)4-羥基異亮氨酸合成的影響,利用HIL017進(jìn)行搖瓶發(fā)酵實(shí)驗(yàn),以ido基因誘導(dǎo)型過(guò)表達(dá)菌株HIL016為對(duì)照,結(jié)果如表4所示,經(jīng)發(fā)酵48 h HIL017的4-羥基異亮氨酸產(chǎn)量和生物量分別為4.62 g/L和7.95 g/L,較對(duì)照菌株HIL016高19.4%和10.3%。誘導(dǎo)劑IPTG對(duì)細(xì)胞生長(zhǎng)有一定的抑制作用,與HIL016采用誘導(dǎo)型質(zhì)粒相比,HIL017采用的pXM01質(zhì)粒中阻遏蛋白編碼基因lacI被敲除,無(wú)需誘導(dǎo)即可組成型表達(dá)ido基因,避免了IPTG對(duì)細(xì)胞生長(zhǎng)的抑制作用,故其生物量高于HIL016。由表4可知,HIL017的4-羥基異亮氨酸單位菌體產(chǎn)量較HIL016高7.4%,表明組成型表達(dá)ido基因可提高菌株HIL017的4-羥基異亮氨酸合成效率。此外,HIL017的副產(chǎn)物(L-異亮氨酸、L-纈氨酸和L-賴(lài)氨酸)生成量均顯著低于HIL016,其原因可能是L-異亮氨酸和L-纈氨酸的合成均需丙酮酸[26],由于組成型表達(dá)ido基因顯著提高了IDO比活力,使得更多的L-異亮氨酸合成4-羥基異亮氨酸,從而消耗更多的丙酮酸用于合成L-異亮氨酸,故L-纈氨酸生成量降低。L-異亮氨酸和L-賴(lài)氨酸均以L-天冬氨酸為前體物[27],L-異亮氨酸的過(guò)量消耗意味著更多的L-天冬氨酸用于L-異亮氨酸合成,故L-賴(lài)氨酸合成量降低。本研究考察了tac啟動(dòng)子組成型表達(dá)ido基因?qū)?-羥基異亮氨酸合成的影響,目前已報(bào)道的用于谷氨酸棒桿菌的組成型啟動(dòng)子包括Ptuf、Psod、PgapA和PCP_2454[28-31]。在后續(xù)研究中,擬利用上述啟動(dòng)子調(diào)控ido基因轉(zhuǎn)錄,以考察其轉(zhuǎn)錄水平與IDO酶活性以及4-羥基異亮氨酸產(chǎn)量的關(guān)系。
2.4 菌株HIL017中pXM01-ido質(zhì)粒的穩(wěn)定性分析
對(duì)HIL017進(jìn)行傳代并測(cè)定其質(zhì)粒穩(wěn)定性及4-羥基異亮氨酸產(chǎn)量,如圖4所示,經(jīng)20 次傳代,pXM01-ido的穩(wěn)定性(98.4%)及HIL017的4-羥基異亮氨酸產(chǎn)量(4.61 g/L)未見(jiàn)顯著降低,表明菌株HIL017中pXM01-ido穩(wěn)定性好。
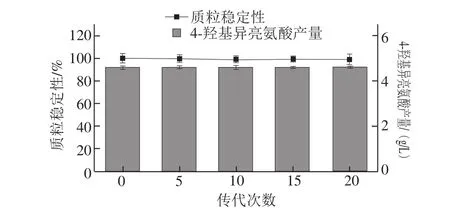
圖4 HIL017中pXM01-ido穩(wěn)定性及4-羥基異亮氨酸產(chǎn)量Fig. 4 Stability of pXM01-ido and production of 4-hydroxyisoleucine by HIL017
2.5 PB試驗(yàn)結(jié)果
選取KH2PO4·3H2O等9 種成分用量為因素,選用N為9的PB試驗(yàn)設(shè)計(jì)。根據(jù)預(yù)試驗(yàn),設(shè)定每個(gè)因素的高水平為低水平的2 倍,進(jìn)行12 次試驗(yàn),以4-羥基異亮氨酸產(chǎn)量作為響應(yīng)值,結(jié)果如表5所示。
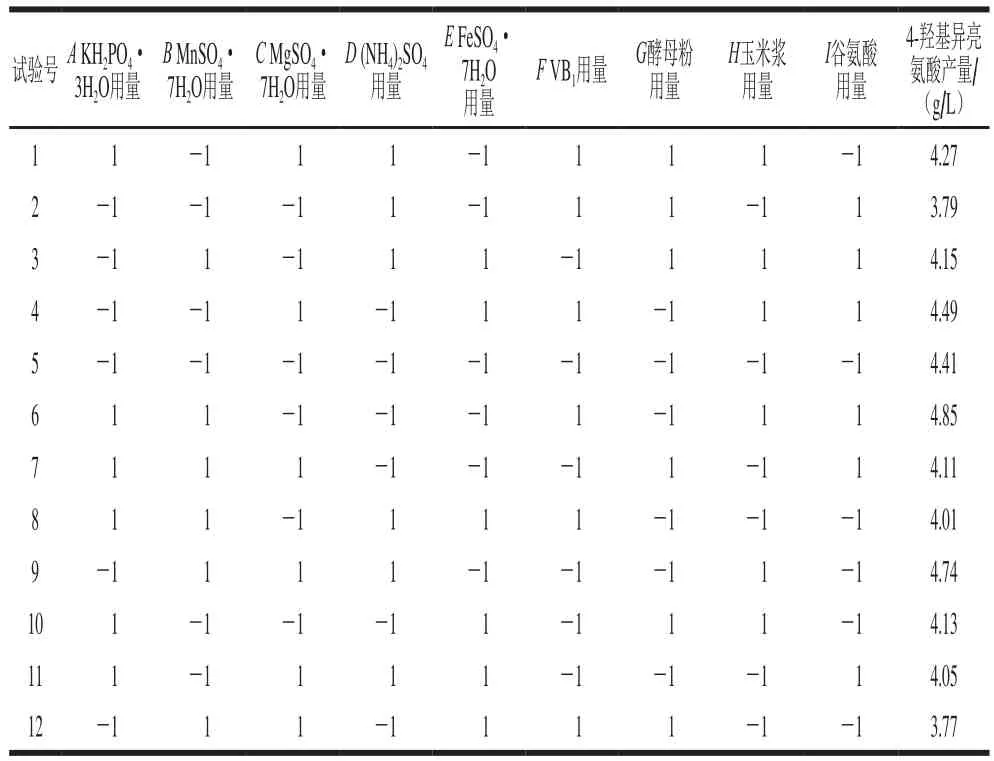
表5 PB試驗(yàn)設(shè)計(jì)及響應(yīng)值Table 5 PB design with response
利用Design Experts V8.0.6分析軟件對(duì)表5中的數(shù)據(jù)進(jìn)行分析,獲得各因素的偏回歸系數(shù)及其顯著性。如表6所示,9 個(gè)因素對(duì)應(yīng)響應(yīng)值影響的顯著性順序?yàn)橛衩诐{>谷氨酸>FeSO4·7H2O>MgSO4·7H2O>酵母粉>KH2PO4·3H2O>VB1>(NH4)2SO4>MnSO4·7H2O,F(xiàn)eSO4·7H2O、玉米漿與谷氨酸用量3 個(gè)因素對(duì)4-羥基異亮氨酸的產(chǎn)量影響極顯著(P<0.01)。由其效應(yīng)值可知,F(xiàn)eSO4·7H2O和谷氨酸用量為負(fù)效應(yīng),說(shuō)明其在高水平時(shí)不利于4-羥基異亮氨酸的合成;而玉米漿為正效應(yīng),說(shuō)明其在高水平時(shí)有利于4-羥基異亮氨酸的合成。因此,在后續(xù)實(shí)驗(yàn)中,在FeSO4·7H2O和谷氨酸高水平基礎(chǔ)上減少其用量,在玉米漿低水平基礎(chǔ)上增加其用量。對(duì)于其他不具顯著影響的因素,正效應(yīng)因素取1水平,負(fù)效應(yīng)因素取-1水平。

表6 PB試驗(yàn)設(shè)計(jì)各因素的回歸系數(shù)及其顯著性Table 6 PB design factors regression coefficients and significance test
2.6 最陡爬坡試驗(yàn)結(jié)果
根據(jù)因素玉米漿、谷氨酸和FeSO4·7H2O用量效應(yīng)的大小確定最陡爬坡試驗(yàn)相應(yīng)變化的方向及步長(zhǎng),設(shè)計(jì)最陡爬坡試驗(yàn)。如表7所示,從第1組到第3組4-羥基異亮氨酸產(chǎn)量逐漸提高,而從第4組后下降,即第3組為最陡爬坡的拐點(diǎn)。故選擇第3組(玉米漿、谷氨酸和FeSO4·7H2O用量分別為35 mL/L、3.0 g/L和0.016 g/L)作為下一步響應(yīng)面試驗(yàn)因素水平的中心點(diǎn)。
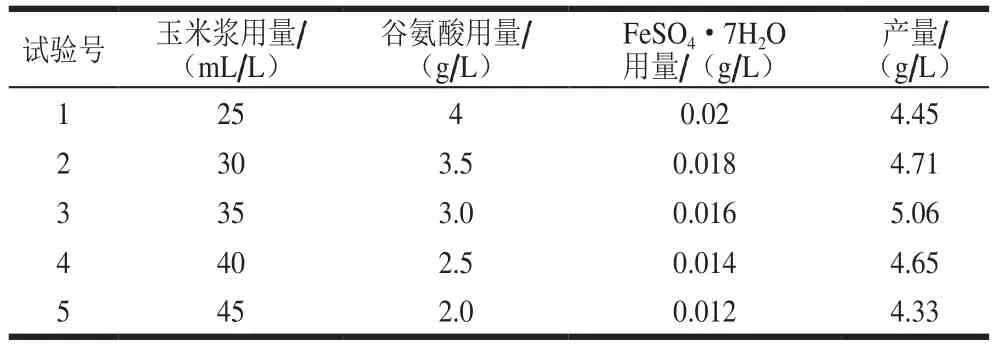
表7 最陡爬坡試驗(yàn)設(shè)計(jì)及結(jié)果Table 7 Design and result of the steepest ascent tests
2.7 Box-Behnken響應(yīng)面試驗(yàn)結(jié)果
Box-Behnken試驗(yàn)結(jié)果如表8所示。利用軟件Design Experts V8.0.6,對(duì)表8的數(shù)據(jù)進(jìn)行多元回歸分析,得到回歸方程為:
Y=5.49+0.12A+0.082B+0.025C+0.068AB-0.037AC+0.018BC-0.37A2-0.14B2-0.03C2
回歸方程的方差分析結(jié)果如表9所示,模型的P值為0.005,說(shuō)明模型的差異性極顯著;相關(guān)系數(shù)R2為0.961 5,表明該模型能夠解釋96.15%的響應(yīng)值變化,校正相關(guān)系數(shù)R2Adj為0.892 1,即僅有變異6.94%不能由該模型來(lái)解釋?zhuān)f(shuō)明模型與試驗(yàn)擬合良好;變異系數(shù)為1.5%,證明試驗(yàn)結(jié)果可信度高;失擬項(xiàng)P為0.418(>0.05),表明該模型失擬不顯著。在模型中,A和A2對(duì)4-羥基異亮氨酸影響極顯著,B和B2對(duì)4-羥基異亮氨酸影響顯著,其余因素對(duì)4-羥基異亮氨酸影響不顯著。
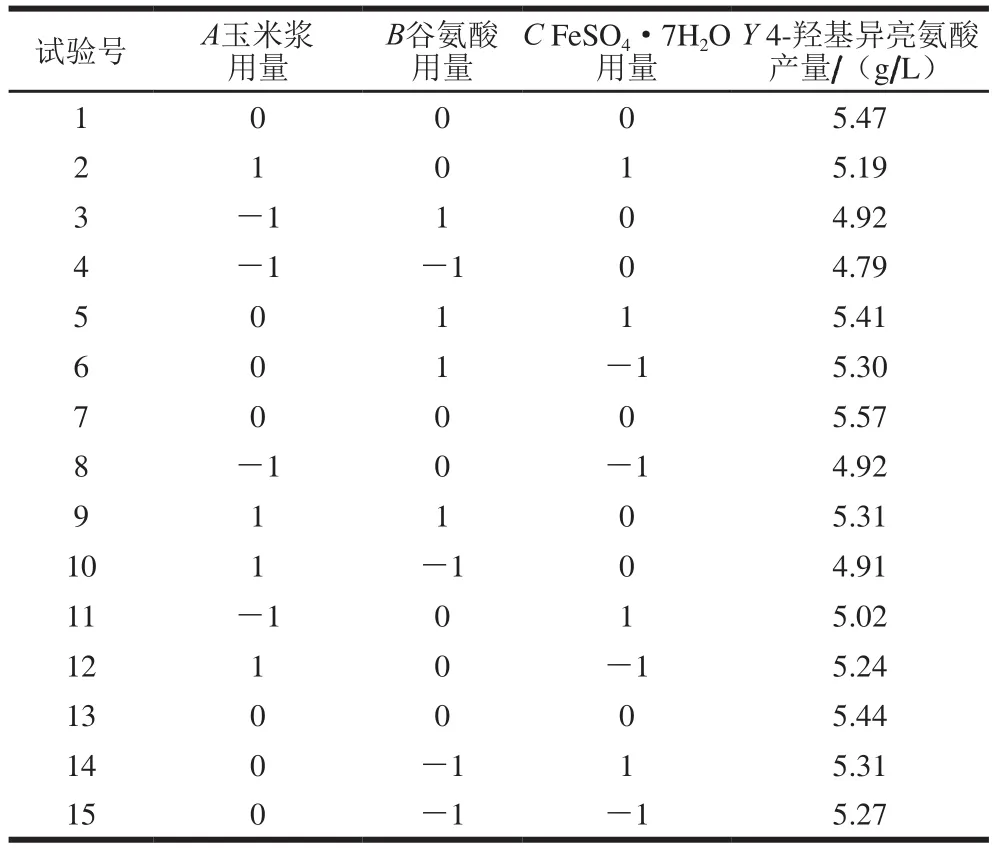
表8 Box-Behnken試驗(yàn)設(shè)計(jì)及結(jié)果Table 8 Box-Behnken design with results

表9 回歸模型的方差分析Table 9 Analysis of variance of regression model
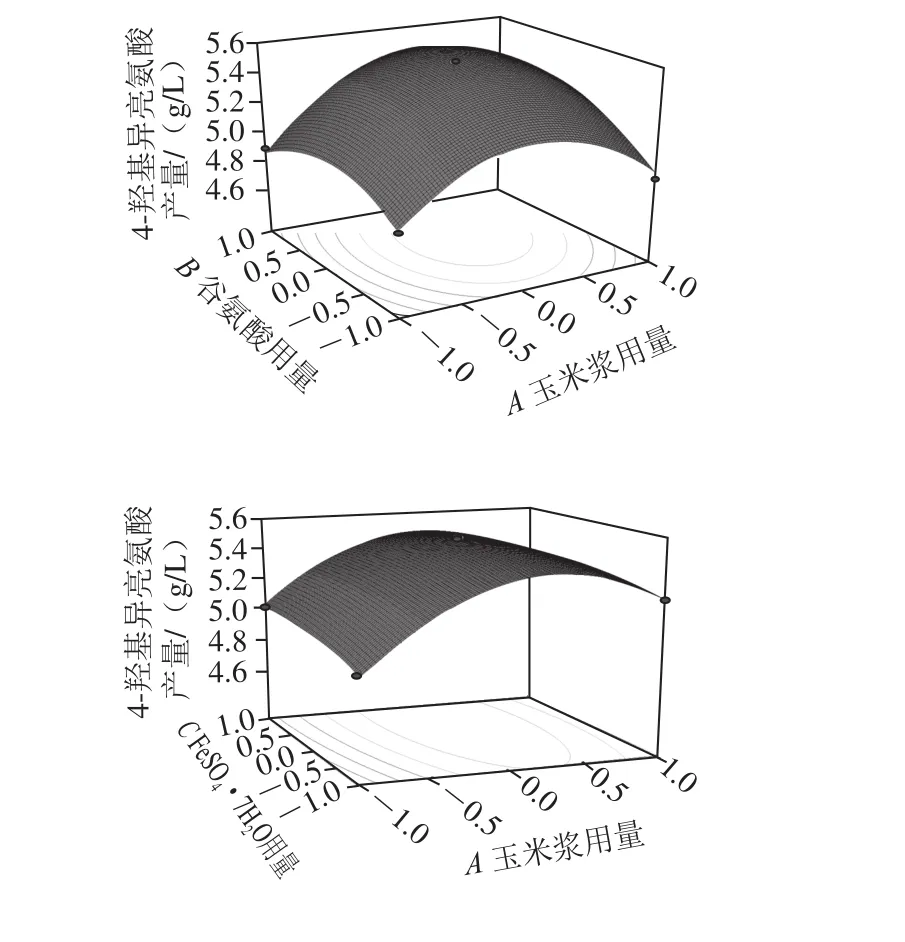

圖5 各因素交互影響4-羥基異亮氨酸產(chǎn)量的響應(yīng)面和等高線圖Fig. 5 Response surface and contour plots for 4-hydroxyisoleucine production influenced by interaction among corn syrup, glutamic acid and FeSO4·7H2O
由圖5可知,F(xiàn)eSO4·7H2O的含量一定時(shí),隨著培養(yǎng)基中玉米漿和谷氨酸用量的增加,4-羥基異亮氨酸產(chǎn)量先增加后降低,在玉米漿和谷氨酸用量分別為34.1 mL/L和2.98 g/L時(shí)達(dá)到最大值5.57 g/L。當(dāng)谷氨酸用量一定時(shí),玉米漿和FeSO4·7H2O用量之間的交互作用并不顯著;而當(dāng)玉米漿用量一定時(shí),谷氨酸和FeSO4·7H2O用量之間的交互作用依然不顯著。通過(guò)Design Experts V8.0.6分析軟件,得到的優(yōu)化結(jié)果為玉米漿34.1 mL/L、谷氨酸2.98 g/L、FeSO4·7H2O 0.016 7 g/L,4-羥基異亮氨酸的理論值為5.57 g/L。
2.8 優(yōu)化培養(yǎng)基效果驗(yàn)證
利用上述優(yōu)化后的培養(yǎng)基進(jìn)行搖瓶發(fā)酵實(shí)驗(yàn),以驗(yàn)證模型的有效性。結(jié)果表明,在最佳培養(yǎng)基條件下,經(jīng)48 h搖瓶發(fā)酵,4-羥基異亮氨酸產(chǎn)量達(dá)到5.53 g/L,與理論值5.57 g/L接近,說(shuō)明該模型能很好地預(yù)測(cè)4-羥基異亮氨酸產(chǎn)量,證明了模型的有效性。利用優(yōu)化后的培養(yǎng)基使得HIL017的4-羥基異亮氨酸產(chǎn)量較優(yōu)化前提高19.7%。
3 結(jié) 論
構(gòu)建了ido組成型表達(dá)質(zhì)粒pXM01-ido及菌株HIL017,其4-羥基異亮氨酸產(chǎn)量較ido誘導(dǎo)型過(guò)表達(dá)菌株HIL016高19.4%,生物量和單位菌體產(chǎn)量也顯著提高,表明ido組成型過(guò)表達(dá)效果更佳。通過(guò)PB試驗(yàn)確定了HIL017發(fā)酵培養(yǎng)基中玉米漿、谷氨酸和FeSO4·7H2O用量為主要影響因素,利用最陡爬坡試驗(yàn)確定這3 種因素的中心值分別為35 mL/L、3.0 g/L和 0.016 g/L。響應(yīng)面法確定的最優(yōu)用量為玉米漿34.1 mL/L、谷氨酸2.98 g/L、FeSO4·7H2O 0.016 7 g/L,此時(shí)4-羥基異亮氨酸理論產(chǎn)量為5.57 g/L。驗(yàn)證實(shí)驗(yàn)結(jié)果表明,所建模型能很好地預(yù)測(cè)4-羥基異亮氨酸產(chǎn)量,最佳條件下為5.53 g/L,較優(yōu)化前提高19.7%。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當(dāng)代水產(chǎn)(2021年10期)2021-12-05 16:31:48
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
今日農(nóng)業(yè)(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中國(guó)果業(yè)信息(2019年10期)2019-11-13 01:21:34
中國(guó)化肥信息(2019年2期)2019-01-18 15:24:35
中國(guó)化肥信息(2019年1期)2019-01-17 21:31:12
中國(guó)化肥信息(2019年4期)2019-01-17 18:47:06
