C-Jun氨基末端激酶與Activin A/Smads通路在體外腦缺血損傷中的相互作用
2019-08-29 02:34:54何金婷梁文昭王姣琦徐忠信
中國實驗診斷學 2019年8期
關鍵詞:信號
何金婷,莽 靖,董 玥,徐 磊,梁文昭,王姣琦,徐忠信
(吉林大學中日聯誼醫院 神經內科,吉林 長春130033)
激活素A(ActA),是目前較為公認的神經保護因子[1]。研究認為,ActA主要通過Smads依賴的信號轉導通路發揮生物學作用,但其活化水平受多種因子的調控,包括卵泡雌激素(FSH)、Smad錨捉蛋白(SARA)等[2,3]。 c-Jun氨基末端激酶(JNK),作為絲裂原活化蛋白激酶(MAPK)家族的成員之一,在上皮—間充質轉化和癌變過程中與TGF-β/Smads信號通路存在交叉對話[4]。作為轉化生長因子β(TGF-β)超家族的成員之一,ActA介導的ActA/Smads信號通路與JNK之間是否也存在著表達調控尚不清楚。本研究通過外源性ActA及JNK抑制劑SP600125干預,初步探討了ActA/Smads與JNK之間的表達調控關系,為ActA/Smads抗缺血性腦損傷神經保護作用的調控提供潛在位點。
1 材料和方法
1.1 主要材料
大鼠腎上腺嗜鉻細胞瘤PC12細胞購自國家實驗細胞資源共享服務平臺。鼠神經生長因子(NGF)購自美國Promega公司。外源性ActA蛋白購自美國Sigma公司。JNK拮抗劑SP600125購自美國BioSource公司。小鼠抗大鼠β-actin抗體購自美國Santa公司,兔抗大鼠Smad3、磷酸化Smad3抗體購自美國ThermoFisher公司;兔抗大鼠JNK1、磷酸化JNK1購自Abcam公司。辣根過氧化物酶標記的羊抗小鼠、羊抗兔二抗購自北京博奧森生物技術有限公司。
1.2 實驗方法
1.2.1PC12細胞的培養及神經元樣轉化 PC12細胞常規培養在含10%馬血清和10%胎牛血清的高糖DMEM培養基。根據細胞生長情況及培養液顏色,每2至3天更換一次培養液。當細胞生長接近80 %融合時,使用含有0.25 %的胰酶進行消化傳代。利用含有50 ng/ml鼠神經生長因子的去血清DMEM高糖培養液連續培養7天,誘導PC12細胞神經元樣轉化[5]。
1.2.2氧糖剝奪模型的復制 利用含有1 mM連二亞硫酸鈉(NaS2O4)的無糖DMEM培養液于37 ℃,5 % CO2、95 % N2的培養箱中培養神經元樣PC12細胞,復制氧糖剝奪(Oxygen Glucose Deprivation,OGD)模型[6]。OGD計時0和1 h。
1.2.3ActA/Smads及JNK信號的干預 為探討ActA/Smads通路與JNK1信號之間的相互作用,本研究首先將神經元樣PC12細胞給與外源性ActA(50 ng/ml)孵育24 h后行OGD處理[7],以探討ActA信號活化對JNK1磷酸化激活的影響。其次,將溶解在DMSO中的SP600125以10 μM的濃度處理神經元樣PC12細胞30 min[8],隨即行OGD處理,以探討抑制JNK磷酸化激活對ActA/Smads信號活化的影響。
1.2.4Western blot蛋白水平表達檢測 將各組提取的總蛋白按照BCA法測定蛋白濃度后調整上樣量。上樣蛋白在100 ℃水中煮沸 5 min,隨后按照操作步驟進行電泳、轉膜、封閉操作。使用含5 %脫脂奶粉的TBST溶液稀釋兔抗大鼠Smad3(1∶1000),磷酸化Smad3(1∶1000)和小鼠抗大鼠β-actin(1∶1000)抗體。一抗4 ℃孵育過夜,次日TBST洗膜,使用羊抗兔或羊抗小鼠的二抗,按照1∶1000的比例室溫孵育1 h,TBST洗膜后ECL顯影壓膠片。凝膠圖像分析系統掃描膠片,吸光度分析計算每條目的條帶與相應內參條帶的灰度值,兩者的比值表示目的蛋白的表達水平。
1.3 統計學分析
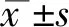
2 結果
2.1 ActA/Smads信號負性調控JNK磷酸化活化
利用Western blot技術對OGD 0 h和1 h組JNK1總蛋白及磷酸化蛋白的表達進行檢測。結果如圖1-A和B,OGD1h組JNK1總蛋白及磷酸化蛋白的表達水平較OGD 0h組均升高。給與外源性ActA孵育24 h后行OGD處理,結果如圖1-C,空白對照組OGD 0 h組相比,OGD 1 h時Smad3總蛋白及磷酸化水平分別升高了27.0%和61.6%,p-JNK1表達升高了27.1%,差別有統計學意義(P<0.05)。與空白對照組各OGD時間點相比,外源性ActA處理組在OGD 0 h及1 h時Smad3磷酸化蛋白水平分別升高了98.7%和44.7%,JNK1磷酸化蛋白水分別減低了67.8%和56.4%,差別有統計學意義(P<0.05)。
2.2 抑制JNK活化上調Act A/Smads通路活性
Western blot檢測JNK1總蛋白及磷酸化蛋白的表達,結果如圖2-A和B,與DMSO組及對照組相比,SP600125抑制劑組JNK1總蛋白表達無明顯變化,但JNK1的磷酸化水平顯著降低。說明SP600125可有效抑制JNK1磷酸化激活。Western blot在對SP600125處理后神經元樣PC12細胞ActA/Smads通路活化水平進行檢測后發現,抑制劑組Smad3總蛋白在OGD 0h及1h時較相應的DMSO組相應時間點升高47.1%和38.3%,磷酸化Smad3升高142.5%和60.5%,差別有統計學意義(P<0.05,如圖2-C和D)。
3 討論
隨著靜脈溶栓和血管內介入治療技術的開展,近年來,針對缺血性腦卒中患者急性期的治療已經取得了長足的進步[9]。然而,受到缺血時間窗的限制,早期再灌注治療的臨床適應癥較少,獲益人群非常有限[10]。近期研究發現,早期神經保護策略的啟動可以在一定程度上為更多的急性缺血性腦卒中患者血管內治療贏得時間[11]。然而,在缺乏外源性干預的情況下,內源性神經保護的作用時間有限,致使損傷機制占優,常常掩蓋了內源性神經保護的效應。因此,通過對影響內源性神經保護作用時程及效力的潛在位點進行研究和適時干預,將為缺血性腦卒中新治療方法的研發提供新思路。
ActA是目前較公認的神經保護因子。作為轉化生長因子β超家族的成員之一,它主要通過ActA/
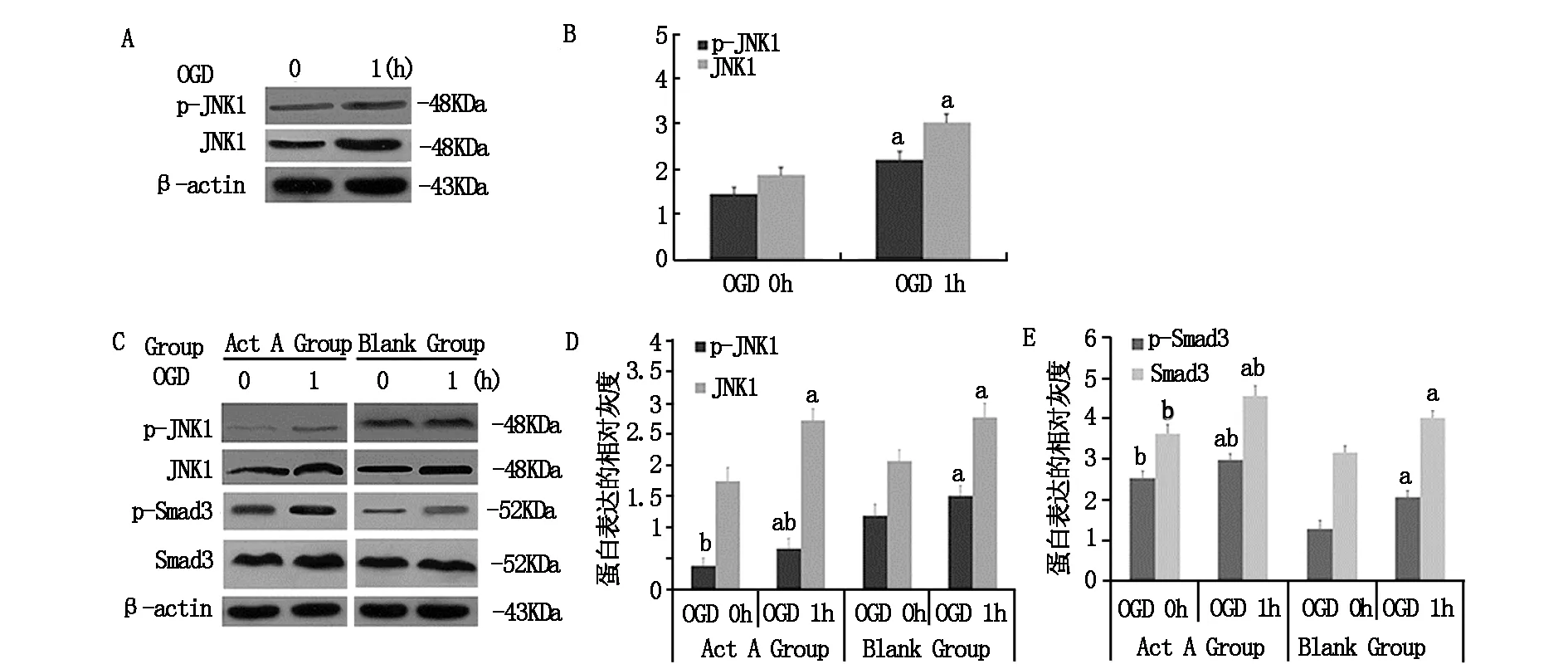
(A)JNK1總蛋白及磷酸化蛋白表達檢測;(B)JNK1總蛋白及磷酸化蛋白表達的灰度分析;(C)JNK1、Smad3總蛋白及磷酸化蛋白表達檢測;(D)JNK1總蛋白及磷酸化蛋白表達的灰度分析;(E)Smad3總蛋白及磷酸化蛋白表達的灰度分析。(a與相應的OGD 0 h組比較P<0.05,b與空白組相應的OGD 時間比較P<0.05,n≥3/組)
圖1 ActA/Smads信號負性調控JNK磷酸化活化
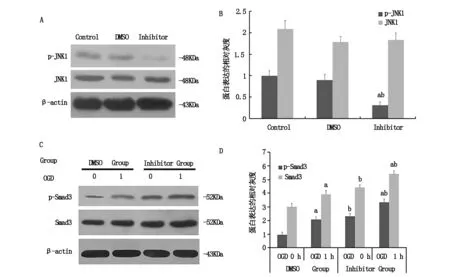
(A)JNK1總蛋白及磷酸化蛋白表達檢測;(B)JNK1總蛋白及磷酸化蛋白表達的灰度分析(a與對照組比較P<0.05,b與DMSO組比較P<0.05,n≥3/組);(C)Smad3總蛋白及磷酸化蛋白表達檢測;(D)Smad3總蛋白及磷酸化蛋白表達的灰度分析。(a與OGD0 h組比較P<0.05,b與DMSO組相應OGD時間比較P<0.05,n≥3/組)Smads通路發揮內源性神經保護作用。由缺血缺氧性損傷介導高表達的ActA,經跨膜信號轉導磷酸化活化細胞內的Smad2/3。磷酸化的Smad2/3再與Smad4形成異二聚體轉入細胞核,調控下游靶基因的表達[12,13]。這樣,磷酸化的Smad2或Smad3的表達水平可一定程度上評估ActA/Smads信號的活化強度。然而,ActA/Smads信號的神經保護作用持續時間較短,活化強度有限[14]。事實上,ActA/Smads信號產生的生物學效能并不單單只是下游Smads信號的級聯反應結果,它還受多種信號通路、細胞因子,從配體、到受體、再到Smads磷酸化等多位點的綜合調控[15]。靶向這些位點的作用機制研究,將為延長ActA內源性神經保護作用時程,擴大作用強度,提升內源性神經保護效力提供潛在的干預位點。
圖2 抑制JNK活化上調Act A/Smads通路活性
前期研究發現,JNK作為MAPK家族的成員之一,在缺血性腦卒中和神經系統退行性疾病中具有促進神經細胞凋亡的作用[16]。JNK1作為其主要的亞基,在腦組織中高表達。磷酸化激活的JNK1可進一步磷酸化活化下游的c-Jun[17]。體內外實驗發現,磷酸化的c-Jun進入細胞核與DNA結合,通過調控靶基因轉錄進一步促進細胞死亡[17]。上述研究提示JNK信號具有促進細胞凋亡的作用。最近研究發現JNK與Smads信號存在交叉對話,急性肝損傷后JNK參與了TGF-β介導的Smad2/3信號的磷酸化活化[18]。但在缺血性腦損傷中JNK與ActA介導的Smads信號之間的相互作用仍不清楚。
為探討該問題,本研究利用神經元樣PC12細胞氧糖剝奪OGD模型,體外模擬腦神經細胞缺血缺氧性損傷。結果發現,OGD1h后JNK1總蛋白及磷酸化蛋白表達升高,提示缺血缺氧性損傷可早期激活JNK信號(圖1-A)。給與外源性ActA 后,JNK1磷酸化水平顯著降低,Smad3磷酸化水平升高,提示由外源性ActA介導的ActA/Smads信號活化,對JNK1信號具有負性調控作用。進一步,通過使用JNK化學抑制劑SP600125,競爭ATP結合位點,有效抑制JNK1磷酸化后,Smad3的磷酸化水平升高。說明JNK信號的磷酸化激活對ActA/Smads信號具有負性調控作用。總結以上研究結果,一方面,JNK信號隨OGD損傷早期激活,負性調控ActA/Smads信號下游Smads位點磷酸化活化。另一方面,由OGD損傷介導的ActA表達上調及ActA/Smads信號激活[7],又對JNK信號的活化起到抑制作用。JNK與ActA/Smads信號的這種交互作用,使得JNK成為ActA/Smads信號調控的一個有效潛在位點。通過下調腦缺血性損傷后JNK信號的活化,我們可以有效上調ActA/Smads通路的磷酸化激活水平,從而進一步抑制損傷介導的JNK內源性激活,實現對ActA/Smads信號的正性調控作用。上述信號調控機制的研究有望通過提高ActA信號的活化水平,為擴大內源性神經保護在缺血性腦卒中的應用提供前期實驗基礎。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
