羅布麻K+通道編碼基因AvAKT1的克隆與表達分析
2019-08-30 02:27:00夏曾潤王文穎劉亞琪王鎖民
草業學報 2019年8期
關鍵詞:植物
夏曾潤,王文穎,劉亞琪,王鎖民*
(1.蘭州大學草地農業生態系統國家重點實驗室,蘭州大學農業農村部草牧業創新重點實驗室,蘭州大學草地農業科技學院,甘肅 蘭州 730020;2.農業農村部富硒產品開發與質量控制重點實驗室,富硒食品開發國家地方聯合工程實驗室,安康市富硒產品研發中心,陜西 安康 725000)
K+是植物生長發育必需的三大營養元素之一,是植物體內含量最豐富的無機單價陽離子和60多種酶的活化劑,參與離子穩態、氣孔運動、膜電位調節等諸多關鍵性生理生化代謝過程。研究表明,植物可通過維持體內高的K+/Na+并發揮K+高效的滲透調節作用來抵御鹽和干旱等非生物脅迫[1]。可見,有效吸收K+并維持體內K+濃度相對穩定對植物適應逆境至關重要。
植物主要通過K+轉運蛋白[2]和K+通道[3]兩大系統從外界環境中吸收K+。普遍認為,Shaker家族的K+通道是介導植物K+吸收和轉運、維持細胞內K+平衡的主要通道。1992年Sentenac等[4]首次從擬南芥(Arabidopsisthaliana) cNDA文庫中篩選到Shaker家族內整流K+通道基因AKT1(ArabidopsisK+transporter 1),隨后相繼從水稻(Oryzasativa)、大麥(Hordeumvulgare)、番茄(Lycopersiconesculentum)、陸地棉(Gossypiumhirsutum)等多種植物中克隆得到[5-8]。Shaker家族K+通道多肽包含1個較短的N末端結構域、6個跨膜片段結構(S1~S6)組成的核心功能區域和1個較長的C末端區域。其中,在S5與S6之間有一個高度保守的膜孔道區域-P結構域(pore loop),其TXXTXGYGD序列是判斷K+通道結構的標志;S4區域含有帶正電的Arg/Lys殘基,是K+通道的電壓感應器,通過應答膜電位的變化改變通道構象,從而控制通道孔的開閉;C末端結構域位于胞質內,含有一個環核苷酸結合位點(cyclic nucleotide-binding domain,cNBD),并且在最靠C末端形成富含疏水酸性殘基區(a domain rich in hydrophobic and acidic residues, KHA),是調節離子通道活性的重要部位[9-12]。此外,AKT1類K+通道在cNBD下游有錨蛋白域(ankyrin-related domain,ANKY),此結構有助于通道與細胞骨架的結合,可促進蛋白之間的相互作用[13-14]。
擬南芥Shaker家族K+通道是目前研究最為透徹的植物K+吸收通道,已有9個家族成員被分離鑒定。內整流K+通道AKT1的組織特異性較強,主要在成熟根表皮、皮層和內皮層細胞等部位表達,具有雙親和性,主要負責根系從土壤中吸收K+,并在調節根系生長發育中也起著重要作用[2,15-16]。akt1突變體的K+吸收能力明顯下降,其根細胞原生質體內向電流消失,在低鉀環境下表現出明顯的敏感表型[17-18]。此外,Shaker家族成員AtKC1能與AKT1形成異源復合K+通道,調控AKT1通道的活性,進而影響低鉀環境中擬南芥根部的K+吸收和積累過程[19-20]。Pilot等[21]發現,擬南芥內整流K+通道KAT1和KAT2主要在保衛細胞中表達,通過介導保衛細胞K+的跨膜流動來調控細胞滲透勢和氣孔運動。也有研究顯示,K+通道在鹽脅迫下可介導低親和性Na+的吸收[22]。然而,Qi等[23]的研究表明,擬南芥中細胞質Na+濃度的提高抑制了AKT1對K+的吸收;在中度鹽脅迫(50 mmol·L-1NaCl)下,擬南芥atakt1-2突變體與野生型植株相比,其Na+濃度及凈吸收速率均沒有顯著差異[24]。AKT1的突變使植物對水分脅迫的響應增強,水培介質中添加聚乙二醇(PEG)時,akt1植株較野生型水分損失少,并表現為蒸騰作用降低,水分消耗減小,在響應脫落酸(ABA)時akt1植株氣孔迅速關閉[25]。越來越多的研究表明,內整流K+通道在參與植物對K+的吸收、轉運以及非生物脅迫響應過程中發揮重要作用。
羅布麻(Apocynumvenetum)系夾竹桃科(Apocynaceae)羅布麻屬(Apocynum)多年生宿根草本植物或小灌木,廣泛分布于干旱半干旱及鹽堿地區,生態幅度廣,具有很強的抗旱耐鹽性,是我國西北荒漠地區一種兼具生態、經濟和藥用價值的新興多用途植物資源,具有良好的產業化前景和巨大的開發潛力[26]。前期研究發現,土培條件下,羅布麻體內K+的積累量與高富鉀植物空心蓮子草(Alternantheraphiloxeroides)、商陸(Phytolaccaacinosa)相當[26];水培條件下,羅布麻在低鉀環境下能保持與正常供鉀等同的K+吸收和利用效率,在鹽或干旱逆境中其可維持體內高且穩定的K+含量和K+/Na+[27],這表明羅布麻體內可能存在高活性的K+吸收途徑。鑒于此,本研究從羅布麻中分離到內整流K+通道AvAKT1的編碼基因,并分析其在低鉀、鹽及滲透脅迫條件下的表達豐度,以期為逆境條件下,羅布麻維持K+穩態的分子機制研究奠定基礎。
1 材料與方法
1.1 試驗材料
羅布麻種子采自新疆阿勒泰戈寶茶股份有限公司種質資源圃。種子經0.5%的高錳酸鉀溶液消毒10 min,用蒸餾水沖洗干凈后在常溫黑暗條件下浸泡24 h,將其均勻撒播于盛有蛭石的穴盤(5 cm×5 cm×5 cm)中,培養至3周齡,移入水培盒(19 cm×13.5 cm×7.5 cm)培養1周。期間澆灌Hoagland營養液[2 mmol·L-1KNO3, 0.5 mmol·L-1KH2PO4, 0.5 mmol·L-1MgSO4·7H2O, 0.25 mmol·L-1Ca(NO3)2·4H2O, 1.25 mmol·L-1CaCl2·2H2O, 0.06 mmol·L-1Fe-citrate, 50 mmol·L-1H3BO3, 10 mmol·L-1MnCl2·4H2O, 1.6 mmol·L-1ZnSO4·7H2O, 0.6 mmol·L-1CuSO4·5H2O, 0.05 mmol·L-1Na2MoO4·2H2O, pH=5.7],營養液每2 d更換一次。培養室晝/夜溫度為25 ℃/22 ℃,光照時間為16 h·d-1,光強度約為230 μmol·m-2·s-1,空氣相對濕度60%~70%。
1.2 材料處理
生長4周齡的羅布麻幼苗,分別做以下處理:1)對照:澆灌Hoagland營養液;2)鹽處理:用含有25 mmol·L-1NaCl的Hoagland營養液處理6 h;3)滲透脅迫:用山梨醇調節滲透勢大小為-0.2 MPa的Hoagland營養液處理6 h;4)鉀處理:先用含有0.01 mmol·L-1K+的Hoagland營養液(分別用2 mmol·L-1HNO3和0.5 mmol·L-1H3PO4代替2 mmol·L-1KNO3和0.5 mmol·L-1KH2PO4,補充0.01 mmol·L-1KCl,用Tris溶液調整pH為5.7)處理3 d,隨后分別用含有0.1和5.0 mmol·L-1K+的Hoagland營養液處理0、6、48 h。處理液每12 h更換一次。處理結束后,分別收集根、莖、葉,迅速于液氮中冷凍用于RNA提取。
1.3 總RNA的提取和cDNA的合成
參照UNIQ-10柱式Trizol總RNA抽提試劑盒(生工生物,上海)說明書提取總RNA。參照PrimeScriptTM1st strand cDNA Synthesis Kit試劑盒(TaKaRa,大連)進行反轉錄,合成cDNA第一鏈。根據SMARTerTMRACE cDNA Kit試劑盒(Clontech,USA)進行RACE反轉錄,合成5′-cDNA和3′-cDNA第一鏈。
1.4 AvAKT1基因的克隆
通過同源多重比對已分離得到的高等植物AKT1核苷酸序列,確定保守序列,利用DNAMAN 6.0和Primer 5.0設計簡并引物P1和P2 (表1)。以鹽處理下羅布麻根系的cDNA為模板,使用PrimeSTAR?高保真DNA聚合酶(TaKaRa,大連)擴增AvAKT1核心片段。PCR擴增在20 μL反應體系中進行,反應程序為:98 ℃預變性2 min;98 ℃變性10 s、53 ℃退火15 s、72 ℃延伸60 s,35個循環;72 ℃再延伸10 min,4 ℃保存。回收純化得到的PCR產物連接pMD19-T載體(TaKaRa,大連),轉入大腸桿菌(Escherichiacoli) DH5α感受態細胞,經藍白斑篩選、鑒定陽性克隆后送至生工生物(上海)股份有限公司測序。
根據測序所得核心片斷序列分別設計5′端和3′端外側特異引物P3、P5和巢式特異引物P4、P6 (表1)分別與試劑盒提供的引物P7、P8配對,采用Phusion?超保真DNA聚合酶(Thermo,USA)進行外側和巢式PCR擴增。外側PCR反應程序:98 ℃預變性30 s;98 ℃變性10 s、55 ℃退火30 s、72 ℃延伸55 s(5′-RACE)和75 s (3′-RACE),35個循環;72 ℃再延伸10 min,4 ℃保存。巢式PCR反應程序:98 ℃預變性30 s;98 ℃變性10 s、55 ℃退火30 s、72 ℃延伸45 s (5′-RACE)和60 s (3′-RACE),35個循環;72 ℃再延伸10 min,4 ℃保存。將測序得到的核心、5′-RACE和3′-RACE序列進行拼接,獲得AvAKT1全長cDNA序列。
1.5 生物信息學分析
將編碼的氨基酸序列在NCBI (www.ncbi.nlm.nih.gov/BLAST)網站上進行Blast比對分析,檢索該基因與其他物種的同源性;序列的翻譯、開放閱讀框(open reading frame,ORF)的分析及氨基酸序列的多重比對通過生物軟件DNAMAN實現;該基因編碼蛋白的等電點和分子量通過在線軟件ProtParam (http://www.expasy.org/tools/protparam.html)預測;利用TMpred (http://www.ch.embnet.org/software/TMPRED_form.html)預測蛋白的跨膜結構域。采用MEGA 6.0軟件中的臨近法構建系統進化樹,校驗參數Bootstrap重復1000次。
1.6 AvAKT1基因表達模式分析
參考SYBR?Premix Ex Taq II試劑盒(TaKaRa,大連)說明書,以P9、P10為擴增引物,推測目的片段長度120 bp,采用ABI 7500熒光定量PCR儀(ABI,美國)檢測不同處理下羅布麻AvAKT1的表達量;以羅布麻肌動蛋白編碼基因AvACT(未發表數據)為內參基因,以P11、P12為擴增引物,推測目的片段長度112 bp。qRT-PCR反應程序為:95 ℃ 30 s;95 ℃ 5 s、60 ℃ 34 s,40個循環。溶解曲線反應條件為:95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。采用2-ΔΔct法[28]計算AvAKT1的相對表達量。每個樣品重復3次。
1.7 數據分析
采用SPSS 18.0進行統計分析,結果以平均值±標準誤表示。用Excel 2010、DNAMAN 6.0、TMHMM、MEGA 6.0等軟件作圖。
2 結果與分析
2.1 AvAKT1基因的克隆
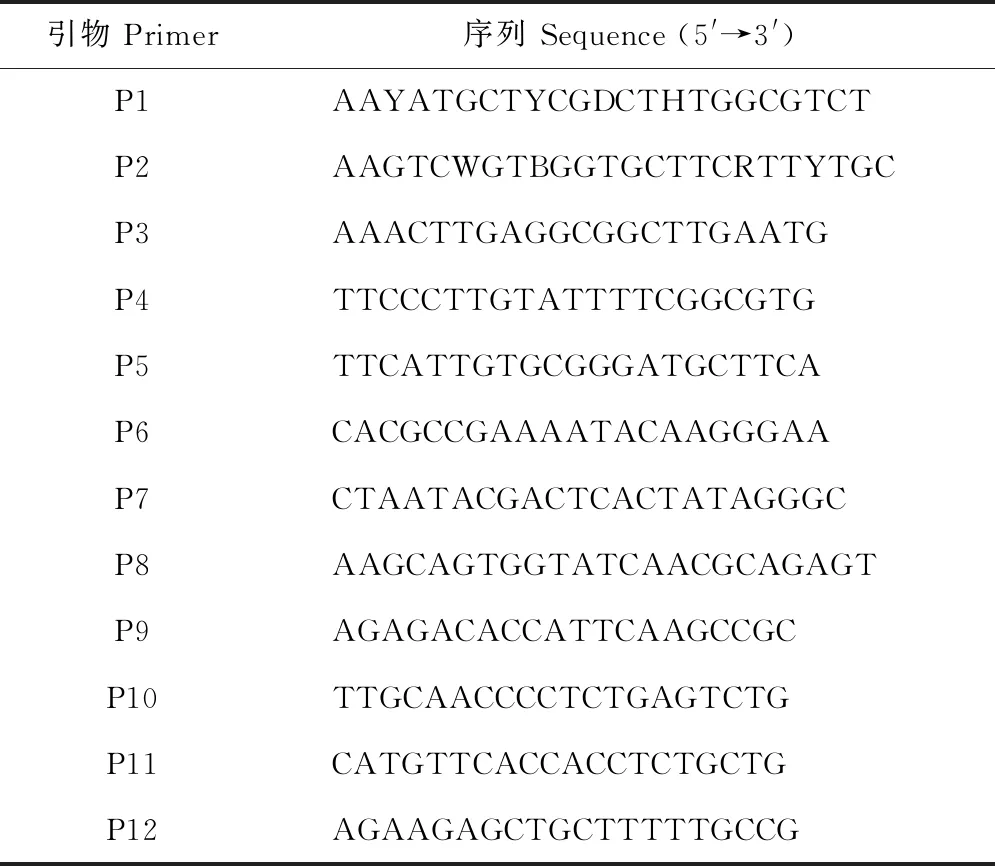
表1 基因克隆和表達分析所用引物序列Table 1 Primer sequences used for gene cloning and expression analysis
以羅布麻根系cDNA為模板,設計簡并引物進行擴增,獲得約749 bp的核心片段(圖1A);根據得到的核心片段設計引物,以RACE產物為模板,進行3′-RACE和5′-RACE擴增,分別獲得長度約為1875 bp (圖1B)和939 bp (圖1C)的片段。測序結果經Blast比對分析,3個cDNA片段均與其他高等植物的AKT1有較高同源性,確定為羅布麻AKT1的編碼基因片段。
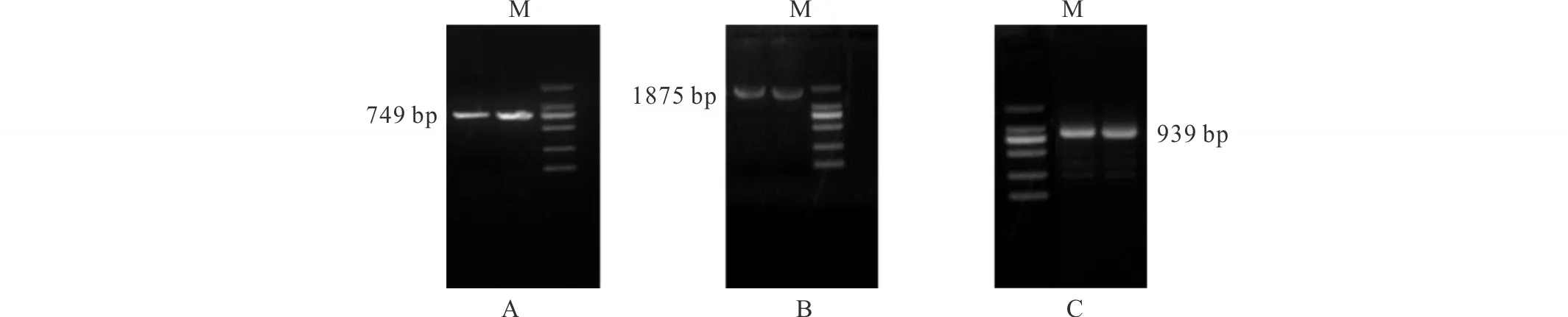
圖1 羅布麻AvAKT1基因PCR產物凝膠電泳檢測Fig.1 Agarose gel electrophoresis of PCR products of AvAKT1 from A. venetum A: 核心片段擴增The RT-PCR product of the partial fragment; B: 3′-RACE; C: 5′-RACE; M: DL 2000 DNA marker.
2.2 AvAKT1基因序列及結構分析
用BioEdit軟件拼接5′端、核心片段和3′端的核苷酸序列,得到一條全長為2906 bp的cDNA序列,該基因命名為AvAKT1。分析表明,AvAKT1的cDNA序列包括2700 bp的開放閱讀框、55 bp的5′非翻譯區(5′-untranslated region,5′UTR)以及151 bp含有poly-A尾巴的3′非翻譯區(3′-untranslated region,3′UTR),推測其編碼一個由899個氨基酸構成的多肽(圖2)。在線軟件ProtParam分析得出AvAKT1編碼蛋白的理論等電點為7.05,蛋白質分子量為101.4 kDa。

圖2 羅布麻AvAKT1的cDNA與預測的氨基酸序列Fig.2 The cDNA sequence and its predicted amino acid residues of AvAKT1 from A. venetum 方框處為起始密碼子(ATG)和終止密碼子(TGA)。The initiation codon (ATG) and termination codon (TGA) were framed with lines.
對AvAKT1基因編碼的氨基酸序列進行疏水跨膜結構域和保守域預測分析,發現其包含6個高度保守的跨膜區(TM 1~TM 6),在TM 5和TM 6之間有一個高度保守的膜孔道區域(pore loop),C端包含一個推測的環核苷酸結合區(cNBD)和錨蛋白重復域(ANKY),且富含疏水性、酸性殘基的KHA結構域(圖3),符合典型的Shaker-like內整流K+通道結構特征。
2.3 同源序列和系統進化分析
運用DNAMAN軟件分析發現該蛋白與其他高等植物的AKT1氨基酸序列的同源性在60%以上。其中,與煙草(Nicotianatabacum)NtAKT1的同源性最高,達76%;與大豆(Glycinemax)GmAKT1、擬南芥AtAKT1和小花堿茅(Puccinelliatenuiflora)PutAKT1的同源性分別為70%、64%和62% (圖3)。
利用MEGA 6.0對AvAKT1與其他植物Shaker K+通道家族構建的系統進化樹表明,Shaker K+通道家族分為5個亞族,AvAKT1隸屬于第Ⅰ亞族(AKT1亞族),與煙草NtAKT1屬同一進化分枝,與雙子葉植物擬南芥AtAKT1、大豆GmAKT1親緣關系較近(圖4)。
2.4 AvAKT1的表達模式分析
正常條件下羅布麻AvAKT1主要在根中表達,在植株地上部的表達量極少,尤其在葉中幾乎不表達(圖5)。采用qRT-PCR檢測不同濃度K+、鹽和滲透脅迫處理下羅布麻根中AvAKT1表達水平的變化,結果表明,K+饑餓后添加5.0 mmol·L-1K+顯著誘導了AvAKT1的表達(圖6A);25 mmol·L-1NaCl和-0.2 MPa滲透脅迫處理時,AvAKT1的表達量在6 h受到顯著誘導(圖6B)。

圖3 羅布麻AvAKT1與其他植物AKT1氨基酸序列多重對比Fig.3 Multiple sequence alignment of AvAKT1 with its homologs in other higher plants At: 擬南芥; Gm: 大豆; Nt: 煙草; Put: 小花堿茅。各基因編碼氨基酸的GenBank登錄號如下: AtAKT1(NM_128222), GmAKT1(XP_003549784), NtAKT1(XP_009619489), PutAKT1(GU327382)。TM 1~TM 6為6個跨膜域。At: A. thaliana; Gm: Glycine max; Nt: Nicotiana tabacum; Put: Puccinellia tenuiflora. Sources of Shaker-like K+ channel family and their GenBank accession numbers are as follows: AtAKT1(NM_128222), GmAKT1(XP_003549784), NtAKT1(XP_009619489), PutAKT1(GU327382). TM 1-TM 6 indicates the structures of the predicted trans-membrane segments of AvAKT1.
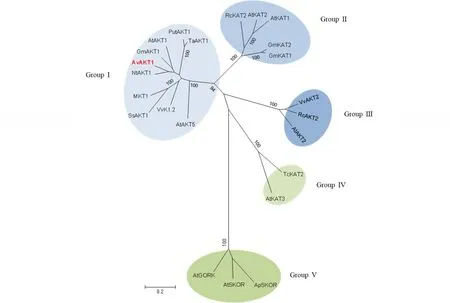
圖4 AvAKT1與其他植物Shaker K+通道家族成員的系統進化分析Fig.4 Phylogenetic analysis of AvAKT1 (marked in red) with related Shaker-like K+ channel family Ap: 空心蓮子草A. philoxeroides; At: 擬南芥A. thaliana; Gm: 大豆G. max; Nt: 煙草N. tabacum; Put: 小花堿茅P. tenuiflora; Rc: 蓖麻Ricinus communis; Ta: 小麥Triticum aestivum; Tc: 可可Theobroma cacao; Vv: 葡萄Vitis vinifera; Zm: 玉米Zea mays. 各基因編碼氨基酸的GenBank登錄號如下Sources of Shaker-like K+ channel family and their GenBank accession numbers are as follows: AtAKT1 (NM_128222), GmAKT1 (XP_003549784), NtAKT1 (XP_009619489), PutAKT1 (GU327382), TaAKT1 (AF207745), ZMK1 (CAA68912), VvK1.2 (FR669116), AtAKT5 (NP_194976), AtKAT1 (At5g46240), AtKAT2 (At4g18290), RcKAT2 (XP_002519693), GmKAT1 (XP_003541662), GmKAT2 (XP_003547208), AtAKT2 (At4g22200), VvAKT2 (XP_002268924), RcAKT2 (XP_002529533), AtKAT3 (At4g32650), TcKAT2 (EOY29638), AtSKOR (At3g02850), AtGORK (At5g37500), ApSKOR (AFO70199). 左下角標尺0.2表示系統進化樹上進化枝的長短The scale bar 0.2 corresponds to the distance of clade on the evolutionary tree.
3 討論
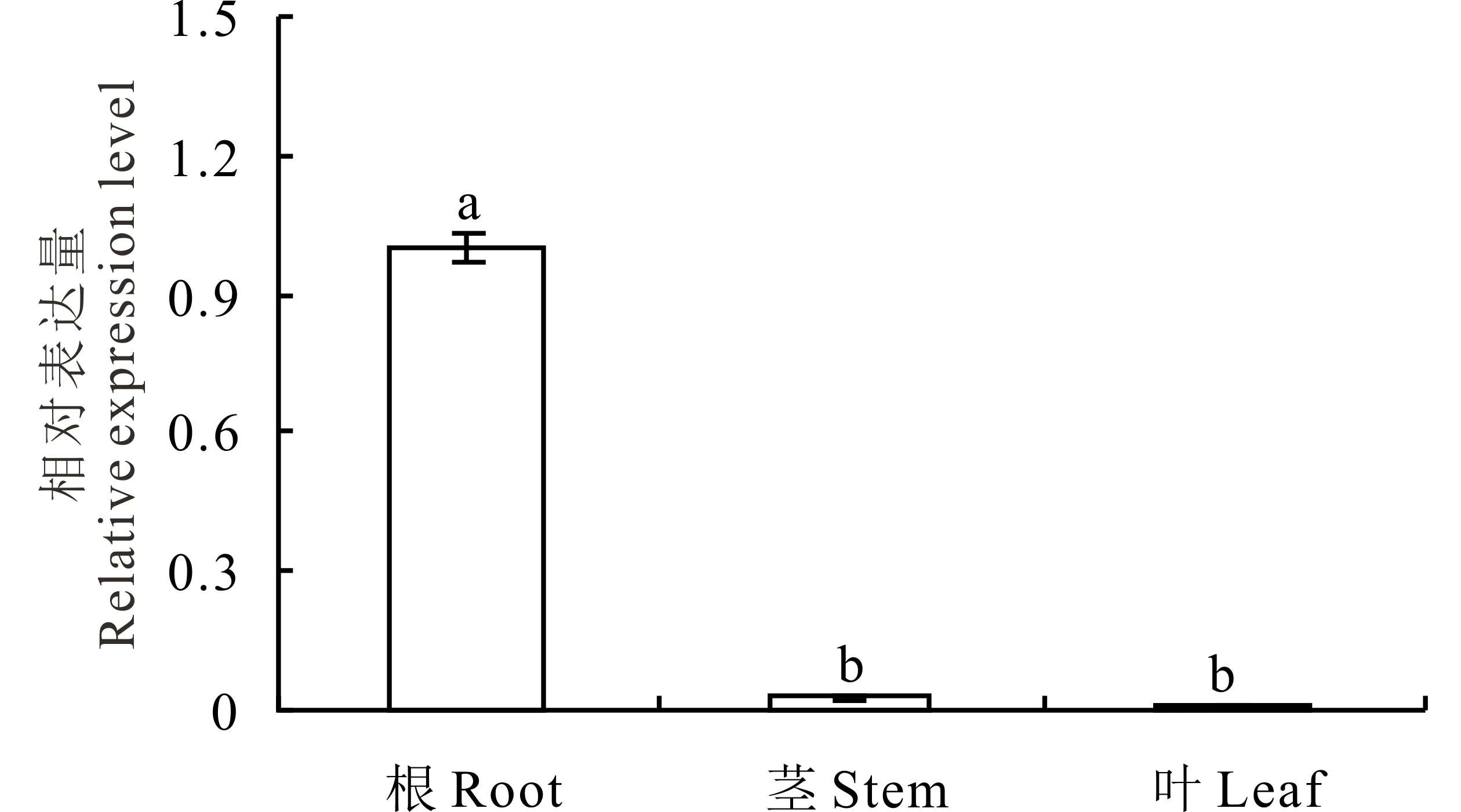
圖5 羅布麻AvAKT1基因在根、莖和葉中的組織特異性Fig.5 The tissue expression characteristics of AvAKT1 in root, stem and leaf不同字母代表在P<0.05水平上差異顯著(Duncan檢驗)。Columns with different letters indicant significant differences at P<0.05 (Duncan’s test).
植物通過根質膜上的K+通道和轉運蛋白從環境中吸收K+。K+通道介導低親和性K+的吸收,是特異性選擇K+通過質膜的跨膜蛋白,也是離子通道中最龐大的家族,可分為Shaker、TPK與Kir-like三大家族[9,29-30]。研究顯示,Shaker家族的成員在植物K+吸收和轉運過程中發揮關鍵作用,對植物適應逆境至關重要[3,31]。
本研究從羅布麻根系中克隆并獲得Shaker家族K+通道蛋白基因AvAKT1,其氨基酸序列的C末端包含Shaker家族K+通道的cNBD和KHA特征結構域,在cNBD下游還存在AKT1類K+通道區別于KAT1類的ANKY結構域,并在第5和第6跨膜區間具有Shaker家族K+通道高度保守和特異性序列TXXTXGYGD及P環孔道區域(圖3),這些特征序列是鑒別Shaker家族K+通道蛋白的重要標準之一,也是K+通道蛋白功能的重要序列單元[12,14,32]。氨基酸序列比對和系統進化樹分析顯示,AvAKT1為第Ⅰ亞族(AKT1亞類)內整流K+通道蛋白,與煙草的同源性最高(圖3,圖4)。
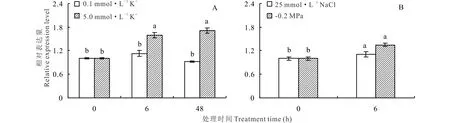
圖6 不同處理下羅布麻根中AvAKT1相對表達量分析Fig.6 The expression level of AvAKT1 in root from A. venetum under different conditions
本研究結果顯示,AvAKT1主要在羅布麻根中高豐度表達(圖5);K+饑餓后,5.0 mmol·L-1K+顯著誘導了羅布麻AvAKT1在根中的表達,0.1 mmol·L-1K+對其表達量幾乎沒有影響(圖6A),表明AvAKT1主要在羅布麻根部發揮作用。已有文獻報道[33-34], AKT1在維持植物離子穩態平衡及增強植物逆境脅迫耐受性等方面起著重要作用。在水稻中超表達OsAKT1后,轉基因植株在干旱條件下生長良好,且根中積累較多的K+;相反,osakt1突變體植株表現為生長不良,且根中K+含量明顯減少[35]。Golldack等[6]研究表明,鹽處理下水稻根吸收Na+與OsAKT1的表達水平有直接關系。在水稻中超表達小花堿茅K+通道蛋白PutAKT1顯著提高了水稻的耐鹽性[36]。Duan等[37]研究發現,鹽脅迫下鹽地堿蓬(Suaedasalsa)SsAKT1的高豐度表達與植株體內K+的積累密切相關。最新研究結果表明[38],ZxAKT1對維持荒漠旱生植物霸王(Zygophyllumxanthoxylum)的K+吸收、K+/Na+穩態平衡至關重要。本試驗中,25 mmol·L-1NaCl和-0.2 MPa滲透脅迫處理6 h時,AvAKT1的表達豐度顯著上調(圖6B),并且發現-0.2 MPa滲透脅迫下羅布麻葉中K+濃度顯著上升、地上部K+/Na+顯著提高,添加25 mmol·L-1NaCl后Na+對滲透勢的貢獻大幅增加,滲透脅迫對幼苗生長的危害得到緩解[27],據此推測AvAKT1可能在調控羅布麻體內K+和Na+選擇性吸收、維持K+濃度穩定,進而增強其滲透調節能力中發揮重要作用。
4 結論
本研究通過RT-RCR和RACE技術從羅布麻根中分離得到內整流K+通道AvAKT1編碼基因,并采用實時定量PCR的方法分析了其在低鉀、鹽及滲透脅迫條件下的表達豐度變化特征。結果表明,AvAKT1基因編碼的氨基酸序列結構符合典型的Shaker-like內整流K+通道結構特征,與其他高等植物AKT1類K+通道蛋白的同源性在60%以上;羅布麻AvAKT1主要在根中表達,受到5.0 mmol·L-1K+、25 mmol·L-1NaCl及-0.2 MPa滲透脅迫的顯著誘導。這可為研究逆境條件下,羅布麻維持K+穩態的分子機制研究奠定基礎。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
