Tim-3對早期梅毒患者外周血CD56dimNK細胞的影響及初步機制①
2019-09-03 03:30:48王政潔閆越穎張杰娜趙培慶
中國免疫學雜志 2019年16期
張 偉 王政潔 閆越穎 張杰娜 趙培慶
(菏澤市立醫院皮膚性病科,菏澤274000)
梅毒是由梅毒螺旋體(Treponema pallidum,Tp)引起的性傳播疾病。最新數據表明,我國梅毒發病率已達32.86/10萬,居全國甲乙類法定傳染病發病第3位,已成為嚴重影響我國公共衛生與人民健康的社會問題[1]。梅毒螺旋體含有豐富的脂蛋白,可作為病原相關分子模式(PAMPs)通過CD14和Toll樣受體1(TLR1)以及TLR2依賴性信號通路激活巨噬細胞和樹突狀細胞(DC)[2,3]。然而梅毒螺旋體表面由于獨特的外膜結構存在,導致缺乏暴露的脂蛋白成分,因此這些PAMPs不容易被TLRs或其他模式識別受體(PRRs)識別,從而造成梅毒螺旋體在組織中可長期復制并傳播[4]。研究發現,患者感染的皮膚局部除了CD4+和CD8+T細胞外,還有大量CD56+NK細胞浸潤,其可能的機制是通過分泌IFN-γ參與皮膚巨噬細胞的激活[5],提示NK細胞在梅毒患者的疾病進程中發揮重要作用。
T細胞免疫球蛋白與黏蛋白結構域分子3(T cell immunoglobulin and mucin domain containing 3,Tim-3)是重要的免疫調節分子。在人類活化的CD4+(Th1)和CD8+細胞毒性T細胞(TC)上首次發現;其表達范圍較廣,包括NK細胞在內幾乎所有免疫細胞上都有Tim-3的表達[6]。Tim-3的功能也異常復雜,與免疫系統的激活和抑制都密切相關[7]。但到目前為止,Tim-3在梅毒患者NK細胞上的表達與功能皆未知。本研究中,我們采用流式細胞術等實驗方法檢測梅毒患者外周血中Tim-3的表達情況并探討其初步功能,以期明確Tim-3在梅毒等性傳播疾病發生發展中的確切生物學作用。
1 材料與方法
1.1 病例獲取、試劑及儀器 早期梅毒患者外周血標本由菏澤市立醫院皮膚性病科門診收集,并與患者簽署研究知情同意書。患者經梅毒螺旋體明膠凝集試驗 (TPPA)及快速血漿反應素試驗 (RPR)確診,且未經治療的早期梅毒患者78例[其中一期患者34例,二期患者40例,隱性梅毒患者4例;其中男62例,年齡(31.0±16.9)歲,女16例,年齡27.0±22.4)歲]。正常對照外周血標本80例取自菏澤市立醫院健康體檢中心,排除任何感染性疾病、自身免疫性疾病等可能影響Tim-3及NK細胞表達情況的疾病。流式抗體CD3、CD16、CD56為Biolegend公司產品(Cat# 300327,Cat#302011,Cat#304605,Biolegend,USA),Human Tim-3為R&D公司產品(#344823,R&D,USA),Lysing Buffer 為BD公司產品(#555899,BD,USA),RPMI1640培養基購自Gibco公司,PBS溶液購自Hyclone公司,人IL-2重組蛋白購自CST公司(#8907,CST,USA),人IL-12及IL-18重組蛋白購自R&D公司(Catalog #219 IL,Catalog #119 BP,R&D,USA),多功能酶標儀產自賽默飛世爾公司(VarioskanFlash),流式細胞儀產自美國BD公司(Aria2),Tim-3及STAT1干擾慢病毒載體由上海吉凱基因有限公司構建(GeneChem,Shanghai)。
1.2 方法
1.2.1 外周血NK細胞的檢測 取患者及正常對照EDTA-K2抗凝外周血100 μl置于流式管底部,依次加入3 μl CD3、CD16、CD56、Tim-3流式抗體,室溫避光30 min;加入1×Lysing Buffer 2 ml,渦旋混勻,置于室溫避光15 min;1 000 r/min離心5 min后棄上清,然后用含0.5%小牛血清的PBS洗滌一次;棄上清,加入約1 ml PBS,充分混勻,上機檢測。
1.2.2 外周血NK細胞分離與培養 按儀器要求調整液流,使液流穩定;然后用微球調節斷點,先調節Ampl,使Gap線位于中上且下面兩個數值保持穩定;分選時,把實際Drop1和Gap值輸入設定窗口,并點擊Sweet Spot鎖定。常規制備無菌管路,以CD3-、CD16+、CD56bright或CD56dim設門分別分選CD56bright及CD56dim兩群NK細胞,細胞分選成功后,加入IL-2(1 000 U/ml)刺激NK細胞增殖,并在含有雙抗的細胞培養基中繼續培養。
1.2.3 ELISA檢測血清及培養上清中細胞因子的分泌 采集患者及正常對照5 ml無抗凝劑外周血標本,待標本完全凝集后充分離心(1 500 r/min離心20 min),以徹底去除可能影響檢測的雜質。然后取上清50 μl進行細胞因子檢測,并設陽性對照、陰性對照及空白對照孔。具體實驗步驟參照試劑說明書,最后用多功能酶標儀在吸光度450 nm處進行讀數,讀取各孔OD值;繪制標準曲線,計算患者及正常對照血清中細胞因子的濃度。
1.2.4 流式細胞儀檢測胞內蛋白IFN-γ水平 收獲培養的細胞1 000 r/min離心5 min,棄上清;加入500 μl 固定液對細胞進行固定,置室溫10 min,1 000 r/min離心5 min,棄上清;加入1.5 ml破膜劑置室溫10 min,1 000 r/min離心5 min,棄上清;加入含0.5%小牛血清的PBS重懸細胞,加入適量抗IFN-γ流式抗體室溫孵育30 min;過400目細胞篩,上機檢測。
1.2.5 Western blot檢測培養細胞中信號通路蛋白的變化 利用蛋白裂解液(含RIPA)抽提細胞總蛋白質,裂解完成后酶標儀BCA法測蛋白濃度,進行蛋白電泳、轉膜,最后將膜與磷酸化STAT1 抗體(#9167,CST,USA),STAT1抗體(#14994,CST,USA)及GAPDH抗體進行孵育,膠片曝光檢測蛋白表達量, GAPDH為內參對照,與目的蛋白比對后得到相對表達量。
1.3 統計學分析 本研究統計學方法利用GraphpadPrism7.0.統計作圖軟件(GraphPad Software Inc.)進行統計學分析,數據采用t檢驗,相關性分析采用Spearman′s rank方法進行統計學分析。P<0.05表示差異有統計學意義。
2 結果
2.1 早期梅毒患者NK細胞數量變化 早期梅毒患者與正常人外周血經NK細胞標記抗體標記后(CD3-CD56+)進行流式分析,結果如圖1所示,患者NK細胞比值(10.8±4.6)與正常對照(9.97±5.02)之間無統計學差異(P>0.05)。
2.2 早期梅毒患者NK細胞上Tim-3表達水平變化 流式結果顯示梅毒患者NK細胞上Tim-3的表達水平較正常對照明顯上調,差異有統計學意義(圖2,P<0.05);進一步分析發現,梅毒患者CD56dimNK上Tim-3表達差異更顯著(P<0.01)在CD56brightNK上Tim-3表達則差異較小(P=0.058)。一般情況下,CD56dim約占NK細胞總數的90%,表達中度親和力的IL-2受體(IL-2R),其主要功能為細胞毒性作用,具有較強的殺傷活性;CD56bright則主要起免疫調節作用。本實驗結果初步提示Tim-3可能通過抑制CD56dim生物學功能,從而導致梅毒患者疾病的發展進程。
2.3 早期梅毒患者CD56dimNK細胞上Tim-3表達水平與血清中IFN-γ相關性分析 為進一步明確CD56dimNK細胞上Tim-3的生物學功能,我們分析了患者Tim-3表達與其血清中IFN-γ分泌水平的相關性,結果如圖3所示,Tim-3表達與其血清中IFN-γ 分泌水平呈明顯負相關性(P<0.01,r=-0.450 6)。
2.4 Tim-3影響CD56dimNK細胞中INF-γ的分泌水平 分選式流式細胞儀以CD56dim設門后對該群細胞進行分選,分選出的細胞置于5%CO2、37℃細胞培養箱中培養,并以IL-2(1 000 U/ml)刺激該群細胞增殖。待細胞擴增后以1×104密度接種于24孔板中,繼續培養24 h后,轉染Tim-3干擾慢病毒載體及對照載體,流式細胞儀及ELISA檢測IFN-γ水平,結果如圖4所示,干擾Tim-3表達后,不管是CD56dimNK細胞中的IFN-γ表達水平,還是培養上清中的IFN-γ水平皆明顯上調。

圖1 早期梅毒患者NK細胞數量與正常人的比較Fig.1 Comparison of number of NK cells between patie-nts with early stage syphilis and control groupNote: A.Analysis of the ratio of NK cells by flow cytometry;B.There was no significant difference for the ratio of circulating NK cells between patients with early syphilis and controls(P>0.05).
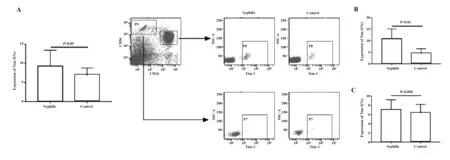
圖2 早期梅毒患者NK細胞上Tim-3表達水平明顯上調Fig.2 Up-regulation of Tim-3 on NK cells in patients with early stage syphilisNote: A.The expression level of Tim-3 on peripheral NK cells was significantly different from controls in patients with early stage syphilis (P<0.05);B.The expression level of Tim-3 was up-regulated on CD56dimNK cells in patients with early stage syphilis (P<0.01);C.There was no significant difference for Tim-3 on CD56brightNK cells between patients with early stage syphilis and controls (P=0.058).

圖3 早期梅毒患者CD56dimNK細胞中Tim-3表達水平與血清中IFN-γ水平負相關Fig.3 Tim-3 expression level on CD56dimNK cells from patients with early syphilis was negatively correlated with the serum IFN-γ level

圖4 干擾CD56dimNK細胞中Tim-3的表達后,IFN-γ分泌水平上調Fig.4 Up-regulation of IFN-γ after interference with Tim-3 on CD56dimNK cellsNote: A.Detection of Tim-3 interference determined by Western blot;B.After interfering with Tim-3 on CD56dimNK cells,the level of IFN-γ secretion was up-regulated (P<0.01);C.The supernatant IFN-γ level was up-regulated after interfering with Tim-3 on CD56dimNK cells (P<0.01).
2.5 Tim-3通過JAK-STAT1通路調控IFN-γ分泌Western blot檢測梅毒患者CD56dimNK細胞JAK-STAT1通路蛋白變化,結果如圖5所示,p-STAT1蛋白在IL-12和IL-18的刺激下較正常對照明顯下調,干擾CD56dimNK細胞中Tim-3的表達后則JAK-STAT1通路蛋白表達上調,同時上清中IFN-γ分泌水平增高(P<0.01)。為進一步明確該通路在Tim-3介導IFN-γ分泌中的作用,我們干擾CD56dimNK細胞中Tim-3表達的同時,也對STAT1蛋白進行干擾,結果顯示IFN-γ升高現象消失(P>0.05),提示Tim-3主要是通過JAK-STAT1通路調控IFN-γ分泌。

圖5 Tim-3通過JAK-STAT1通路調控IFN-γ分泌Fig.5 Tim-3 regulates IFN-γ secretion via JAK-STAT1pathwayNote: A.p-STAT1 protein in CD56dimNK cells is significantly down-regulated in patients with syphilis;B.Up-regulation of p-STAT1 protein in CD56dimNK cells after Tim-3-sh transfected;C.Detection of STAT1 interference using Western blot method.Tim-3 regulates IFN-γ secretion via the JAK-STAT1 pathway;D.IFN-γ was up-regulated in the culture supernatant in CD56dimNK cells after interference with Tim-3 (P<0.01).Tim-3 regulates IFN-γ secretion via the JAK-STAT1 pathway;E.There was no significant difference for IFN-γ in CD56dimNK cells when both interference with STAT1 and Tim-3 (P>0.05).
3 討論
人類NK細胞是一群異質性細胞群體,一般分為CD56dim和CD56bright兩個亞群。 外周血中CD56dim約占NK細胞總數的90%,其功能主要為細胞毒性反應,在各類感染性疾病中發揮重要作用。相反,CD56bright只占外周血中NK細胞總數的10%,該類細胞主要生物學功能是免疫調節作用,可能在自身免疫性疾病中發揮關鍵作用[8,9]。與 CD56brightNK細胞不同,CD56dimNK細胞表達CD16 (Fcγ受體) 和KIR 受體[10]。在抗病原體感染方面,NK細胞一方面通過細胞與細胞直接接觸來殺傷受病原體感染的細胞,另一方面通過分泌細胞因子(如IFN-γ)來控制病原體在細胞中的增殖[11],因此NK細胞在抗擊病原體感染的第一線發揮重要的生物學作用。但相關研究表明早期梅毒患者IFN-γ分泌水平往往低于正常人,從而導致病原體清除障礙。
人Tim基因家族主要包括Tim-1、 Tim-3和 Tim-4,該家族成員與免疫調節、感染等密切相關。課題組前期研究表明Tim-4可以調控巨噬細胞免疫學功能,在SLE、糖尿病、膿毒血癥等炎癥性疾病中發揮關鍵生物學作用[12-15]。但Tim-3與其他兩個成員不同,它幾乎在所有免疫細胞中皆有表達,可以調控幾乎所有細胞的功能。但到目前為止它通過調控NK細胞參與梅毒等性傳播疾病的機制還不清楚。有研究報道梅毒患者外周血NK細胞數量較正常對照低[16],但在本研究中,我們發現NK細胞總數在梅毒患者與正常對照之間無統計學差異,可能的原因是檢測群體差異導致。有趣的是,我們發現CD56dimNK細胞上Tim-3的表達水平差異非常顯著,由于Tim-3是重要的免疫調節分子,因此我們認為Tim-3可能通過調控CD56dimNK細胞在梅毒感染中發揮關鍵生物學功能。
CD56dim主要表達中度親和力的IL-2受體(IL-2R),主要功能是行使細胞毒性作用,具有較強的殺傷活性。有研究表明乙肝患者外周血中CD56dimNK細胞可分泌大量IFN-γ,進而導致乙肝病毒持續感染[17],表明CD56dimNK細胞可能通過調控IFN-γ的分泌水平參與感染性疾病的病理進程。由于我們發現早期梅毒患者CD56dimNK細胞中Tim-3表達水平與血清中IFN-γ水平明顯負相關,提示Tim-3可能是該進程中的關鍵調控分子。已有研究表明Tim-3在成熟NK細胞上高表達,體外實驗發現NK細胞上的Tim-3具有共刺激功能。在腫瘤模型中,Tim-3的表達與預后負相關,主要功能是抑制抗腫瘤作用,阻斷Tim-3可逆轉某些腫瘤中NK細胞的衰竭狀態,表明Tim-3通過調控NK細胞促進了腫瘤的進展過程[18]。在感染性疾病如HIV患者中,有學者發現表達Tim-3的CD8+T細胞在功能上呈現耗竭狀態,Tim-3表達水平也與病毒載量正相關[19]。在乙型肝炎病毒和丙型肝炎病毒感染者中,高表達的Tim-3也與T細胞功能耗竭相關,抗Tim-3治療后可以使T細胞毒性功能部分恢復,并能顯著降低病毒滴度[20]。Tim-3在某些胞內細菌如結核分枝桿菌感染中,Tim-3的表達可促進人類T細胞功能增強,利用小鼠模型發現Tim-3有助于在結核分枝桿菌感染過程中維持T細胞的耗竭狀態。機制上,T細胞上的Tim-3與其配體Gal-9相互作用,后者又可刺激巨噬細胞,從而增強抗菌功效。因此Tim-3對免疫功能的調控具有雙向性[21]。本研究中,CD56dimNK細胞上高表達的Tim-3生物學功能到底如何?我們首先分析了梅毒患者Tim-3表達水平與血清中IFN-γ分泌水平的相關性,結果發現Tim-3的表達與血清中IFN-γ分泌水平呈明顯負相關性。IFN-γ等Th1型細胞因子可增強T細胞免疫功能,在促進梅毒螺旋體清除以及皮損愈合方面起重要作用。因此,我們初步認為CD56dimNK細胞上高表達的Tim-3可能抑制了IFN-γ的分泌,從而導致梅毒螺旋體的清除障礙。為進一步深入探討Tim-3通過調控CD56dimNK細胞對IFN-γ分泌水平的影響,我們分選出CD56dimNK細胞,同時轉染Tim-3干擾慢病毒載體及對照載體,結果發現干擾Tim-3表達后,不管是CD56dim中的IFN-γ表達,還是培養上清中的IFN-γ水平皆明顯上調。該結果更進一步明確了我們的理論推測。文獻結果表明IFN-γ的分泌主要受JAK-STAT1通路的正向調控[22]。基于以上理論基礎,我們檢測了梅毒患者CD56dimNK細胞中JAK-STAT1通路蛋白表達變化,結果顯示梅毒患者CD56dimNK細胞中JAK-STAT1通路蛋白在IL-12和IL-18的刺激下較正常對照明顯下調,干擾CD56dimNK細胞中Tim-3的表達后則JAK-STAT1通路蛋白表達上調,同時上清中IFN-γ分泌水平增高,該結果進一步明確Tim-3主要通過調控JAK-STAT1通路進而影響CD56dimNK細胞中IFN-γ的分泌水平。為進一步明確該通路在Tim-3介導IFN-γ分泌中的作用,我們干擾CD56dimNK細胞中Tim-3表達的同時,也對STAT1蛋白進行干擾,結果顯示IFN-γ升高現象消失。以上結果更進一步表明Tim-3通過JAK-STAT1通路調控IFN-γ分泌,進而影響梅毒患者的疾病進程。
總之,在本研究中我們發現Tim-3通過下調JAK-STAT1通路蛋白,進而導致IFN-γ分泌減少。由于IFN-γ是梅毒螺旋體清除過程中的重要細胞因子,因此高表達的Tim-3水平可能抑制了梅毒螺旋體的清除進程,最終影響了梅毒患者的疾病進程。
綜上所述,我們的結果初步闡釋了Tim-3可能參與梅毒疾病進程的分子機制,為闡明梅毒發生發展提供了新的理論基礎,可能在梅毒的預后判斷中發揮重要作用。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
學苑創造·A版(2015年11期)2016-01-14 09:03:27
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
中國中醫藥現代遠程教育(2014年16期)2014-03-01 04:28:54
