EB病毒相關伯基特淋巴瘤差異基因表達分析
2019-09-10 07:22:44鄭雨薇肖華王維文劉雯
青島大學學報(醫學版) 2019年4期
鄭雨薇 肖華 王維文 劉雯

[摘要] 目的 從分子層面分析EB病毒(EBV)相關伯基特淋巴瘤(BL)的差異表達基因(DEGs),為其診療提供新的思路。方法 從美國國立生物技術信息中心(NCBI)獲取EBV陽性與陰性BL相關基因芯片數據集GSE100458。通過morpheus篩選出DEGs,分別將上調和下調基因輸入Database for Annotation Visualizationand Integrated Discovery(DAVID)數據庫,進行GO分析;進一步采用cytoscape篩選出10個TOP基因,實時熒光定量PCR方法驗證其在EBV陽性與陰性BL細胞系中表達情況。結果 分析EBV陽性和EBV陰性的BL細胞系的前400個DEGs,包括200個上調基因和200個下調基因,上調基因主要集中在生長因子活性通路、肝素結合通路、細胞外間隙通路;下調基因主要集中在伽馬微管蛋白結合通路、內質網膜通路、逆行性小囊泡運輸通路。對DEGs進行蛋白質相互作用分析后,篩選出核心基因乙酰輔酶A羧化酶β(ACACB)等10個基因。實時熒光定量PCR檢測結果顯示,在EBV陽性與陰性BL組間,CDH1、PRPF19、UBE2N、F2、H6PD、VEGFA、YARS共7個基因的轉錄表達差異有統計學意義(t=3.878~32.601,P<0.05),ACACB、CTTN、IGF1共3個基因表達差異無統計學意義(P>0.05)。結論 EBV感染可導致宿主細胞基因表達譜的改變,生物信息學分析法可初步篩選出DEGs和相互作用的蛋白,為進一步深入研究提供有價值的信息和思路。
[關鍵詞] 皰疹病毒4型;伯基特淋巴瘤;基因表達;生物信息學
[中圖分類號] R373.9 ?[文獻標志碼] A ?[文章編號] ?2096-5532(2019)04-0423-06
[ABSTRACT] Objective To analyze the differentially expressed genes (DEGs) in EB virus (EBV)-associated Burkitt lymphoma (BL) at molecular level, and to provide new ideas for its diagnosis and treatment. ?Methods The EBV-positive and-negative BL-related gene microarray data set GSE100458 was obtained from the National Center for Biotechnology Information (NCBI). DEGs were screened out by Morpheus, and the up-regulated and down-regulated genes were entered into the Database for Annotation Visualization and Integrated Discovery (DAVID) for GO analysis. Ten top genes were identified by cytoscape, and their expression in EBV-positive and EBV-negative BL cell lines was verified by real-time fluorescence quantification PCR. ?Results The top 400 DEGs of EBV-positive and EBV-negative BL cell lines were analyzed, including 200 up-regulated genes and 200 down-regulated genes. The up-regulated genes were mainly concentrated in the growth factor activity pathway, heparin binding pathway, and extracellular gap pathway. The down-regulated genes were mainly concentrated in the gamma-tubulin binding pathway, endoplasmic reticulum pathway, and retrograde small vesicle transport pathway. After protein interaction analysis of DEGs,10 core genes such as acetyl coenzyme A carboxylase beta (ACACB) were screened out. Quantitative real-time PCR results showed that there were significant differences in transcriptional expression of 7 genes (CDH1, PRPF19, UBE2N, F2, H6PD, VEGFA, and YARS) between EBV-positive and EBV-negative BL groups (t=3.878-32.601,P<0.05), and there were no significant diffe-rences in the expression of ACACB, CTTN, and IGF1 (P>0.05). Conclusion EBV infection can lead to changes in the gene expression profiles of host cells. Bioinformatics analysis can preliminarily screen out DEGs and interacting proteins, providing valuable information and ideas for further research.
[KEY WORDS] herpesvirus 4, human; Burkitt lymphoma; gene expression; computational biology
EB病毒(EBV)是重要的DNA腫瘤病毒,與伯基特淋巴瘤(BL)、霍奇金病、某些T細胞源和B細胞源性非霍奇金淋巴瘤、鼻咽癌以及胃癌等多種人類淋巴細胞及上皮細胞腫瘤的發生發展均密切相關[1]。地方性BL感染的EBV有兩種不同類型的毒株,即1型和2型EBV,兩種類型毒株特異性有所不同[2]。1型EBV毒株廣泛分布在世界各地,盡管淋巴細胞樣細胞系中1型EBV轉化率比2型高,?但2型EBV在非洲地區更為常見;兩種類型EBV均在非洲地方性BL中被發現,并且流行于撒哈拉以南的非洲健康人群中。然而,尚無研究對比初發地方性BL中1型和2型EBV的突變情況,病毒變異是否影響腫瘤的發生尚不清楚[3]。基因表達匯編(GEO)是基因表達數據資源庫,其中匯聚大量的表達譜數據,是多種生物信息學分析均需要使用的重要資源[4-5]。本文通過GEO數據庫得到有意義的基因,進而驗證EBV影響的BL的發生機制。
1 資料與方法
1.1 數據收集
從美國國立生物技術信息中心(NCBI)GEO數據庫下載GSE100458基因表達譜,包括4種細胞系的14個克隆,其中Mutu、Akata和Awia細胞系分別包含2個EBV陽性和2個EBV陰性克隆;Elijah細胞系包含1個EBV陽性和1個EBV陰性克隆。GSE100458基于GPL23270 [HG-U133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array平臺分析得來。
1.2 差異表達基因(DEGs)鑒定
采用R語言和Limma包篩選出DEGs,符合P值<0.01和|logFC|≥2標準。通過morpheus進行基因表達聚類分析,根據噪聲比大小篩選出DEGs[6]。
1.3 DEGs的GO分析
獲得DEGs后,將DEGs列表提交到線上軟件Database for Annotation,Visualization and Integrated Discovery(DAVID,https://david.ncifcrf.gov)v6.8,分別進行上調基因和下調基因的GO分析[7]。GO(http://www.geneontology.org)是分析DEGs和基因產物功能的工具,包括生物過程、分子功能和細胞成分[8]。最終,通過GO分析選擇被強化的DEGs的功能,以P<0.05為差異有統計學意義。
1.4 蛋白質-蛋白質相互作用(PPI)網絡分析
采用Search Tool for theTetrieval of Interacting Genes(STRING)數據庫分析DEGs之間的相互作用關系,以解釋更深刻的分子機制。首先將DEGs列表提交到STRING上,采用組合分數>0.4選取實驗證實的相互作用,然后用cytoscape軟件分析構建PPI網絡系統。根據蛋白質與蛋白質相互作用度篩選出頂級中樞基因。
1.5 DEGs表達
1.5.1 細胞培養 將EBV陽性淋巴瘤細胞系Raji、Daudi,EBV陰性淋巴瘤細胞系Ramos,置于37 ℃、含體積分數0.05的CO2、99%濕度培養箱常規培養,細胞匯合度達90%時棄培養液,收集2個不同批次細胞用于RNA提取。
1.5.2 RNA提取 用TRIzol法提取3種細胞系RNA,RNA濃度在200~1 000 μg/L、純度A260/A280在2.0左右為可使用范圍[9]。
1.5.3 cDNA合成 采用cDNA逆轉錄試劑盒滅活基因組DNA,反應體系為10 μL,內含:5×gDNA Eraser Buffer 2 μL,gDNA Eraser 1 μL,RNA 1 μg,剩余用RNase Free dH2O補足。反應條件42 ℃、2 min。向上述反應體系中加入:PrimeScript RT Enzyme Mix 1 μL,RT Primer Mix 1 μL,5×Prime Script Buffer 4 μL,RNase Free dH2O 4 μL;反應條件為:37 ℃孵育15 min,85 ℃反應5 s。
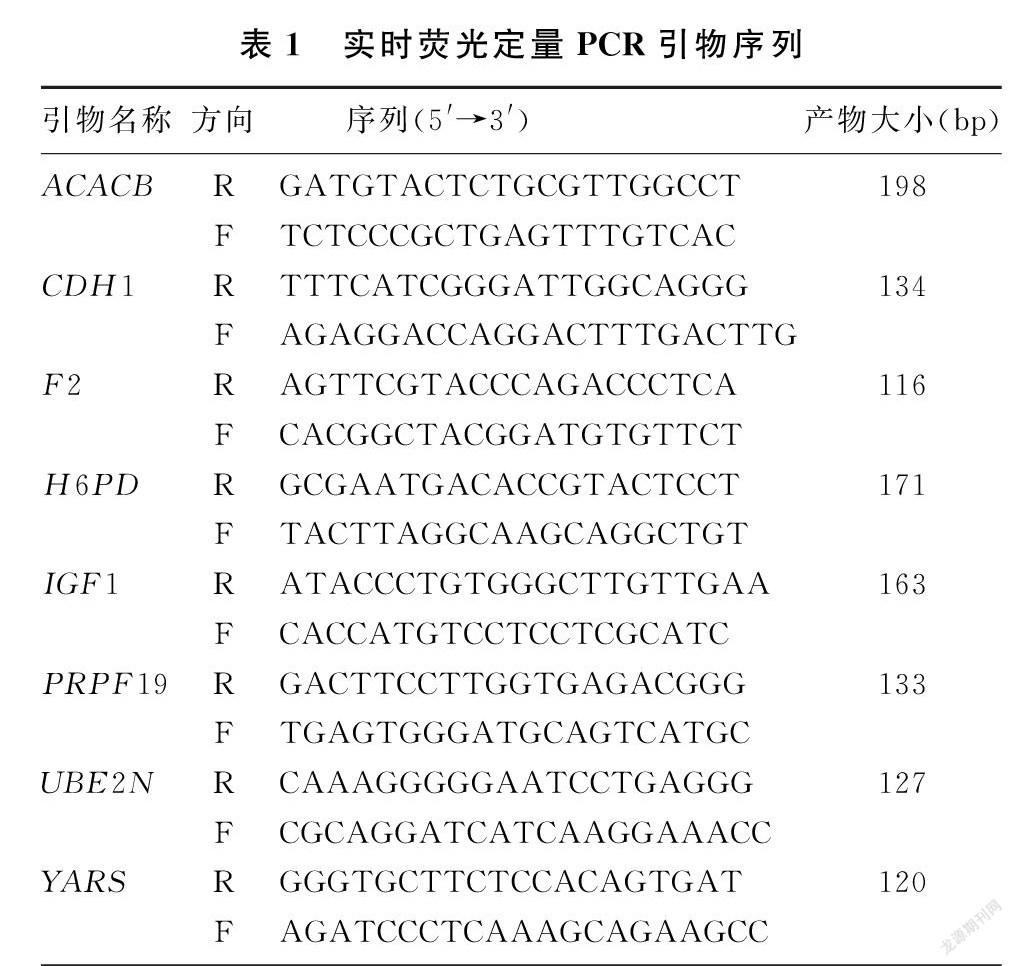
1.5.4 實時熒光定量PCR 擴增引物在Pubmed上進行設計,引物序列見表1。反應體系為20 μL,包括:SYBR GreenⅠreal-time PCR Master Mix 10 μL,上下游引物分別0.5 μmol/L,cDNA模板2 μL,無RNase水6 μL。PCR擴增條件為:95 ℃預變性15 min;94 ℃、15 s,60 ℃、 30 s,70 ℃、30 s,擴增40個循環,每次循環結束后進行熒光檢測。同時擴增內參照基因GAPDH以檢測RNA的完整性和反轉錄效果。擴增結束后,95 ℃、1 min,55 ℃、30 s后逐步升溫至95 ℃進行溶解曲線分析,確定擴增產物的特異性。
1.6 統計學分析
采用GraphPad Prism(version 6.01)軟件進行統計學分析,計量資料結果以±s形式表示,數據間比較采用t檢驗。以P<0.05為差異有統計學意義。
2 結 ?果
2.1 聚類分析熱圖
從GEO數據集中篩出的GSE100458由7個EBV陽性BL細胞系和7個EBV陰性BL細胞系組成,根據Morpheus軟件,以噪聲比絕對值≥0.5的標準,篩選出200個上調基因和200個下調基因。DEGs聚類表達譜見圖1A。
2.2 GO分析
利用DAVID在線軟件對400個DEGs分類分析結果顯示,上調基因主要參與調控生長因子活性通路(GO:0008083)[10]、調控肝素結合通路(GO:0008201)、細胞外間隙通路(GO:0005615)、片狀偽
2.3 PPI分析
在蛋白質間相互作用圖中,顏色越亮、面積越大,說明與其他蛋白質關聯度越大[11](圖1D)。從中篩選10個關聯度最大的蛋白,其全稱、縮寫、關聯度見表2。
2.4 基因表達驗證
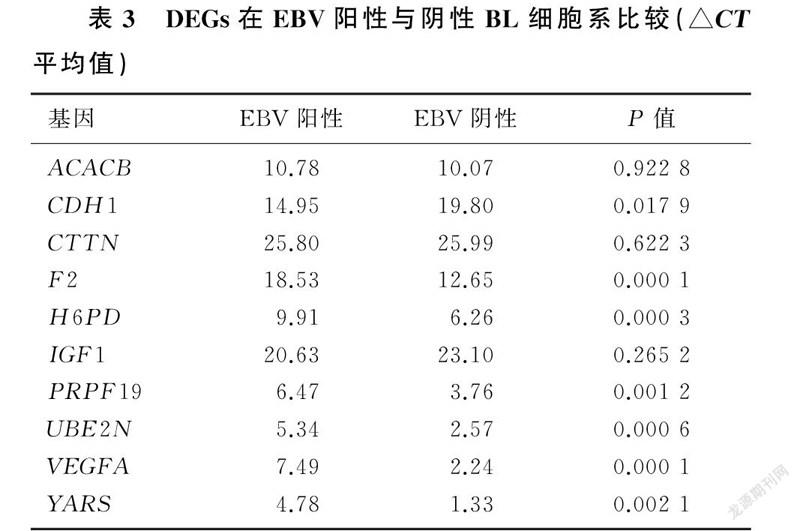
實時熒光定量PCR結果顯示,EBV陽性與陰性BL細胞系中ACACB、CTTN、IGF1表達差異無統計學意義(t=0.103~1.294,P>0.05),與網絡數據不相符;CDH1、PRPF19、UBE2N表達差異有統計學意義(t=3.878~9.730,P<0.05),網絡數據和實驗數據相符合;F2、H6PD、VEGFA、YARS表達差異均有統計學意義 (t=7.055~32.601,P<0.01),網絡數據和實驗數據不一致。見表3。
3 討 ?論
生物信息學分析可為認識蛋白結構及功能研究提供線索。隨著生物技術和信息技術的發展,新的生物信息學具有經濟、便利、快捷的特點,可以為科學研究提供有價值的信息。本研究利用生物信息學軟件,對EBV陽性和EBV陰性的BL細胞系相關特征進行了研究,初步篩選出表達差異基因,研究蛋白質與蛋白質之間相互作用的關系,為深入開展相關研究提供了理論依據。
本實驗研究表明,上調基因參與的生長因子活性通路(GO:0008083)刺激腫瘤的發生。孕酮受體為乳癌生長因子活性的潛在指標,有研究表明原發性乳癌孕酮受體缺乏與預后不良相關,低孕酮受體狀態可作為乳癌腫瘤細胞中活化生長因子信號傳導的指標,因此可以用于侵襲性腫瘤表型對抗激素治療[12]。據此,上調基因參與的生長因子活性通路可能對BL的治療和預后有著重要的作用。上調基因參與的調控肝素結合通路(GO:0008201)可促進神經母細胞瘤分化[13]。本文研究中的上調基因參與調控的肝素結合通路也可能與促進BL的分化生長有關,刺激了腫瘤的發生。上調基因參與的細胞外間隙通路(GO:0005615)為細胞之間的電耦合,由稱為間隙連接的質膜相互作用而介導[14]。哺乳動物心臟的主要連接蛋白是連接蛋白43,連接蛋白43含量的降低是心臟病的一般致病特征,并且其他連接蛋白類型的表達水平的變化可能有助于改變患病心臟的電生理功能。細胞外間隙通路影響細胞的電生理功能,這是否意味著電生理功能也對BL的發生發展有著重要作用,還需要進一步的實驗證明。上調基因參與片狀偽足通路(GO:0030027),轉移到遠處位點的腫瘤細胞在其通過血管期間受到血流動力學剪切力,片狀偽足通路可以減少其通過血液轉移的阻力[15]。BL通過淋巴途徑轉移,片狀偽足通路對淋巴液的剪切力可能對其也有一定的影響,導致了BL的轉移,上調了腫瘤的發生發展。上調基因參與鈣介導的信號通路(GO:005084),有研究結果表明線粒體裂變與細胞溶質鈣信號通路形成正反饋環,促進肝細胞癌細胞自噬[16]。線粒體裂變與細胞溶質鈣信號通路形成的正反饋環也可能參與了BL細胞的自噬,但這并不一定意味著其會下調BL的發生。上調基因參與鉀離子跨膜運輸通路(GO:0071805),有學者進行的熒光和膜片鉗實驗表明,電壓可以驅使分子插入和離開脂質雙層,從而打開和關閉鉀離子的傳輸,進而影響腫瘤的形成[17]。下調基因參與伽馬微管蛋白結合通路(GO:0043015),在果蠅卵母細胞分裂的前中期(Ⅰ期)中雙極紡錘體組裝和動粒微管附著需要伽馬微管蛋白通路[18-19]。這表明伽馬微管蛋白通路可能與祖細胞的分裂增殖有關,進而抑制腫瘤的生成。下調基因參與poly(A)?RNA結合通路(GO:0044822),Poly(A)RNA結合蛋白以高親和力和特異性結合聚腺苷RNA,并出現了許多新的作用,這些作用有助于基因表達的微調[20]。結合本研究結果認為其應該與BL的下調有關。下調基因參與內質網膜通路(GO:0005789),有研究表明蘿卜硫素是一種天然存在的化學預防劑,通過線粒體介導的細胞凋亡有效抑制肝細胞癌細胞的增殖,內質網膜通路被認為可能是蘿卜硫素誘導肝癌細胞凋亡的最重要機制[21]。內質網膜通路參與了腫瘤細胞凋亡,這與BL發生的下調有關。下調基因參與蛋白酶體調節粒子通路(GO:0008541),蛋白質穩態失衡導致細胞功能障礙、細胞死亡和病變。真核生物中的蛋白質降解途徑主要分為蛋白酶體介導的降解和溶酶體介導的降解,目前蛋白酶體可作為癌癥的治療靶標[22],今后BL的治療可能要與蛋白酶體調節粒子通路相聯系。下調基因參與逆行性小囊泡運輸通路(GO:0006890),高爾基體是分泌途徑的中心分選和生物合成中樞,并且使用囊泡運輸來回收酶[23]。若逆行性小囊泡運輸通路機制發生損傷,則高爾基體功能出現損傷進而影響腫瘤的發生發展。
本文通過對DEGs進行功能和通路分析及蛋白質相互作用分析,篩選出ACACB、VEGFA、YARS、IGF1、CTTN、F2、CDH1、UBE2N、H6PD以及PRPF19基因作為該網絡的關鍵基因,這些關鍵基?因主要參與了生長因子激活信號通路、鉀離子的跨膜轉運信號通路、伽馬微管蛋白結合信號通路等,并可能在BL的發生發展過程中發揮重要的作用。CDH1、PRPF19、UBE2N在網絡數據中表現出的差異與本文結果相符合。CDH1為抑癌基因,其突變與多種腫瘤相關,CDH1敲除可能會影響乳腺上皮細胞的增殖分化,干擾T細胞的激活化生[24]。推測CDH1可能影響了T細胞的代謝通路,與免疫應答通路有相互作用關系,其突變大大增加了BL發生的可能性。PRPF19突變可導致RNA末端的剪接紊亂并下調RNA表達,推測EBV可能通過抑制腫瘤細胞中PRPF19的表達,干擾正常細胞的分化代謝,進而影響BL細胞的凋亡[25]。UBE2N是一種泛素結合酶,泛素蛋白酶體途徑是目前己知的所有真核生物體內具有高度選擇性的最為重要的蛋白質降解途徑,其突變在BL中影響了蛋白質的降解[26]。本文結果顯示,F2、H6PD、VEGFA、YARS的表達差異有統計學意義,但是其網絡數據分析的結果和實驗驗證結果不相符,可能與樣本量不足有關。F2為一種凝血因子,EBV可抑制其在BL中的表達,影響凝血因子的級聯反應,可能與部分BL病人的出血傾向有關[27]。本文結果還顯示,H6PD在EBV陽性細胞系中低表達,H6PD與細胞中物質代謝有關,EBV可能通過抑制其表達,導致正常細胞中物質代謝紊亂,影響BL細胞的凋亡。VEGF是一種血管內皮細胞的特異性有絲分裂原,在體外可促進血管內皮細胞的生長,在體內可誘導血管增生,增加BL的新生血管。YARS是氨酰-tRNA合成酶,參與tRNA的合成,影響BL的核酸合成代謝,對BL可能有重要意義[28]。
綜上所述,EBV在BL中引起的上述基因表達差異可能影響BL的細胞間連接而影響其轉移,影響DNA的損傷后修復功能而影響BL的發生。但本文研究由于樣本量的限制,結果可能存在一定的局限性,需要擴大樣本量,進一步探討與BL發生發展相關的DEGs。
[參考文獻]
[1] LICCIARDI F, VAN DEN BOOGAARD M, DELLE PIANE M, et al. EBV-related hodgkin lymphoma in an ICF2 patient: is EBV susceptibility a hallmark of this ICF subtype[J]? J Clin Immunol, 2019,39:1-3.
[2] KAYMAZ Y, ODUOR C I, YU H B, et al. Comprehensive transcriptome and mutational profiling of endemic burkitt lymphoma reveals EBV type-specific differences[J]. Molecular Cancer Research, 2017,15(5):563-576.
[3] UCCINI S, AL-JADIRY M F, CIPPITELLI C A, et al. Burkitt lymphoma in Iraqi children: a distinctive form of sporadic disease with high incidence of EBV+ cases and more frequent expression of MUM1/IRF4 protein in cases with head and neck presentation[J]. Pediatric Blood & Cancer, 2018,65(12): e27399.
[4] WANG Anping, ZHANG Guibin. Differential gene expression analysis in glioblastoma cells and normal human brain cells based on GEO database[J]. Oncology Letters, 2017,14(5):6040-6044.
[5] 董政權,魏壘. 骨關節炎基因差異譜的生物信息學分析[J]. 中國組織工程研究, 2019,23(3):335-340.
[6] CHAI Yi, TAN Feng, YE Sumin, et al. Identification of core genes and prediction of miRNAs associated with osteoporosis using a bioinformatics approach[J]. Oncology Letters, 2019,17(1):468-481.
[7] GAUDET P, DESSIMOZ C. Gene ontology: pitfalls, biases, and remedies[J]. Methods Mol Biol, 2016,1446:189-205.
[8] VESZTROCY A W, DESSIMOZ C. A gene ontology tutorial in python[J]. Methods Mol Biol, 2017,1446:221-229.
[9] 趙丹蕊. EB病毒相關胃癌中RASSF1A基因甲基化狀態和表達的研究[D]. 青島:青島大學, 2017.
[10] LEE H Y, LI C C, HUANG C N, et al. INHBA overexpression indicates poor prognosis in urothelial carcinoma of urinary bladder and upper tract[J]. Journal of Surgical Oncology, 2015,111(4):414-422.
[11] GROMIHA M M, YUGANDHAR K, JEMIMAH S. Protein-protein interactions: scoring schemes and binding affinity[J]. Current Opinion in Structural Biology, 2017,44:31-38.
[12] CUI X J, ZHANG P, DENG W L, et al. Insulin-like growth factor-Ⅰ inhibits progesterone receptor expression in breast cancer cells via the phosphatidylinositol 3-kinase/akt/mammalian target of rapamycin pathway: progesterone receptor as a potential indicator of growth factor activity in breast[J]. Molecular Endocrinology, 2003,17(4):575-588.
[13] GAVIGLIO A L, KNELSON E H, BLOBE G C. Heparin-binding epidermal growth factor-like growth factor promotes neuroblastoma differentiation[J]. FASEB Journal, 2017,31(5):1903-1915.
[14] SEVERS N J. Pathophysiology of gap-junctions in heart-disease[J]. Journal of Cardiovascular Electrophysiology, 1994,5(5):462-475.
[15] XIONG Niya, LI Shun, TANG Kai, et al. Involvement of caveolin-1 in low shear stress-induced breast Cancer cell motility and adhesion: roles of FAK/Src and ROCK/p-MLC pathways[J]. Biochimica et Biophysica Acta-Molecular Cell Research, 2017,1864(1):12-22.
[16] HUANG Qichao, CAO Haiyan, ZHAN Lei, et al. Mitochondrial fission forms ?a positive feedback loop with cytosolic Calcium signaling pathway to promote autophagy in hepatocellular carcinoma cells[J]. Cancer Letters, 2017,403:108-118.
[17] SI Wen, LI Zhanting, HOU Junli. Voltage-driven reversible insertion into and leaving from a lipid bilayer: tuning transmembrane transport of artificial channels[J]. Angewandte Chemie-International Edition, 2014,53(18):4578-4581.
[18] HUGHES S E, BEELER J S, SEAT A, et al. Gamma-Tubulin is required for bipolar spindle assembly and for proper kinetochore microtubule attachments during prometaphase Ⅰ in drosophila oocytes[J]. PLoS Genetics, 2011,7(8): e1002209.
[19] MOUTINHO-PEREIRA S, DEBEC A, MAIATO H. Microtubule cytoskeleton remodeling by acentriolar microtubule-organizing centers at the entry and exit from mitosis in Drosophila somatic cells[J]. Mol Biol Cell, 2009,20(11):2796-2808.
[20] WIGINGTON P, WILLIAMS C. Poly(a)RNA-binding proteins and polyadenosine RNA:new members and novel functions[J]. Wiley Interdisciplinary Reviews RNA, 2014,5(5):601-622.
[21] ZOU Xiang, QU Zhongyuan, FANG Yi, et al. Endoplasmic reticulum stress mediates sulforaphane-induced apoptosis of HepG2 human hepatocellular carcinoma cells[J]. Molecular Medicine Reports, 2017,15(1):331-338.
[22] JANG H H. Regulation of protein degradation by proteasomes in cancer[J]. J Cancer Prev, 2018,23(4):153-161.
[23] COTTAM N P, UNGAR D. Retrograde vesicle transport in the Golgi[J]. Protoplasma, 2012,249(4):943-955.
[24] 周學付,陳國星,彭曉飛. 粵北地區遺傳性彌漫型胃癌與CDH1種系突變關系初探[J]. 消化腫瘤雜志(電子版), 2018,10(2):101-104.
[25] ZHANG Yuzhe, LI Yu′e, LIANG Xiao, et al. Crystal structure of the WD40 domain of human PRPF19[J]. Biochemical and Biophysical Research Communications, 2017,493(3):1250-1253.
[26] DIKSHIT A, ZHANG J Y. UBE2N plays a pivotal role in maintaining melanoma malignancy[J]. Oncotarget, 2018,9(100):37347-37348.
[27] GIROLAMI A, COSI E, FERRARI S, et al. Prothrombin: another clotting factor after FV that is involved both in blee-ding and thrombosis[J]. Clinical and Applied Thrombosis-Hemostasis, 2018,24(6):845-849.
[28] SADJJADI S M, EBRAHIMIPOUR M, SADJJADI F S. Comparison between Echinococcus granulosus sensu stricto (G1) and E. canadensis (G6) mitochondrial genes (cox1 and nad1) and their related protein models using experimental and bioinformatics analysis[J]. Computational Biology and Chemistry, 2019,79(79):103-109.
(本文編輯 黃建鄉)
