水分脅迫下竹葉花椒的生理響應
2019-09-10 07:22:44王紀輝侯娜粱美
江蘇農業學報 2019年3期
王紀輝 侯娜 粱美


摘要:為揭示水分脅迫對竹葉花椒(Zanthoxylum armatum DC)生理指標的影響,以及竹葉花椒耐旱機制,以1年生花椒實生苗為試驗材料,采用盆栽試驗,對水分脅迫后竹葉花椒幼苗的超氧化物歧化酶活性、過氧化物酶活性、相對含水量、丙二醛含量、葉綠素含量、可溶性蛋白質含量的變化規律進行了研究。結果表明,干旱脅迫下竹葉花椒超氧化物歧化酶、過氧化物酶活性保持良好,丙二醛含量緩慢增加,相對含水量和葉綠素含量逐漸下降,可溶性蛋白質含量呈“V”型變化。相關性分析結果表明,部分指標間表現出顯著正(負)相關,部分指標間呈一定的相關性但不顯著。主成分分析結果表明,過氧化物酶活性、丙二醛含量、相對含水量、葉綠素含量、可溶性蛋白質含量與竹葉花椒耐旱性關系密切,可作為評價其耐旱性的指標。
關鍵詞:竹葉花椒;水分脅迫;生理響應
中圖分類號: Q945.78
文獻標識碼:A
文章編號: 1000-4440( 2019) 03-0676-06
竹葉花椒(Zanthoxylum armatum DC)是蕓香科( Rutaceae)花椒屬落葉小喬木,花椒中酰胺類物質和揮發油含量豐富,因而麻香味十分濃郁,是受廣大消費者青睞的佐料[1]。花椒因具有溫中散寒、除濕、止痛等功能,又被納入傳統中藥材的行列,成為藥食兩用的重要植物[2]。近幾年來,花椒產業發展十分迅速,已成為中國西南、西北地區的重要經濟樹種[3]。花芽分化是花椒產量高低的決定性因素,確保一定的花芽數量,是花椒高產穩產的基礎[4]。但不良環境會減弱花椒花芽分化程度,造成花椒大幅度減產[5]。因此,研究水分脅迫對竹葉花椒生理指標的影響,為初步闡明竹葉花椒耐旱機理,解決其目前面臨的干旱問題奠定基礎。研究發現,干旱脅迫下作物超氧化物歧化酶( SOD)協同過氧化物酶(POD)共同發揮作用,清除自由基[6]。干旱脅迫阻礙葉綠素合成,葉綠素分解速率大于合成速率,葉綠素含量整體上表現為下降趨勢;干旱脅迫導致植物細胞膜系統被破壞,膜脂發生過氧化,丙二醛(MDA)含量增加,相對含水量(RWC)逐漸下降,而可溶性蛋白質(SP)合成速率大于分解速率,其含量整體上呈增加趨勢[7]。研究結果表明,水分脅迫在一定程度上能提高SOD和POD活性,加速膜脂過氧化,增加MDA含量,降低葉片相對含水量,刺激蛋白酶合成新的可溶性蛋白質來適應環境[8-10]。張寅媛等的研究結果表明,干旱脅迫能迫使景天科植物葉片相對含水量下降,細胞膜系統遭到破壞,MDA含量增加,脅迫初期SOD活性升高、后期下降,可溶性蛋白質含量增加[11]。胡宏遠等研究發現,水分脅迫能抑制葉綠素合成,加速其分解,并降低凈光合速率[12]。干旱脅迫能阻礙作物光合作用,降低凈光合速率[13-14]。目前,有關竹葉花椒對水分脅迫的生理響應方面的研究報道較少。本研究以1年生竹葉花椒幼苗葉片為試驗材料,揭示水分脅迫下竹葉花椒的生理響應,旨在為竹葉花椒栽培、品種選擇以及耐旱性能等研究提供科學依據。
1 材料與方法
1.1 試驗材料
竹葉花椒1年生實生苗取自貴州大學林學院苗圃基地,屬于延遲脫水耐旱類型。2016年2月在半拱形覆膜日光溫室中育苗。盆栽試驗于2017年2月在貴州大學林學院科研基地半拱形覆膜日光溫室中進行,取園土、腐殖質、砂子按質量比4:1:1混合,裝入高27 cm、直徑26 cm的塑料盆中,每盆裝土12 kg,土壤田間最大持水量為24.12%。每盆栽植l株,正常澆水,保持幼苗正常生長。移栽后2個月,移到遮雨棚中進行控水處理。設置4個水分梯度處理:對照( CK),土壤含水量為田間最大持水量的70% -80%;輕度脅迫(Tl),土壤含水量為田間最大持水量的50% - 60%;中度脅迫(T2),土壤含水量為田間最大持水量的40% - 50%;重度脅迫(T3),土壤含水量為田間最大持水量的30% - 40%。每個處理10盆,3次重復。采用稱量法控水和補水,每天18:00稱量,補充當天消耗的水分,使土壤含水量控制在設定范圍之內。
1.2 測定指標與方法
達到設定的水分梯度后,維持20 d。采集各處理植株上相同節位(從基部向上第3-5節)、相同部位發育成熟的葉片,帶回實驗室立即測定葉片相對含水量(RWC),其余樣品裝入密封袋,置于4℃冰箱中保存,3d內完成所有指標測定,每個指標重復測定3次。
SOD活性采用氮藍四唑光還原法測定,POD活性采用愈創木酚法測定,MDA含量采用硫代巴比妥酸法測定,葉片相對含水量采用烘干法測定,葉綠素含量采用高俊鳳等的方法[15]測定,可溶性蛋白質含量采用考馬斯亮藍G-250染色法測定。
1.3 數據處理
采用Origin9.1軟件制圖,用SPSS19.0中的Duncan's進行差異顯著性分析。
2 結果與分析
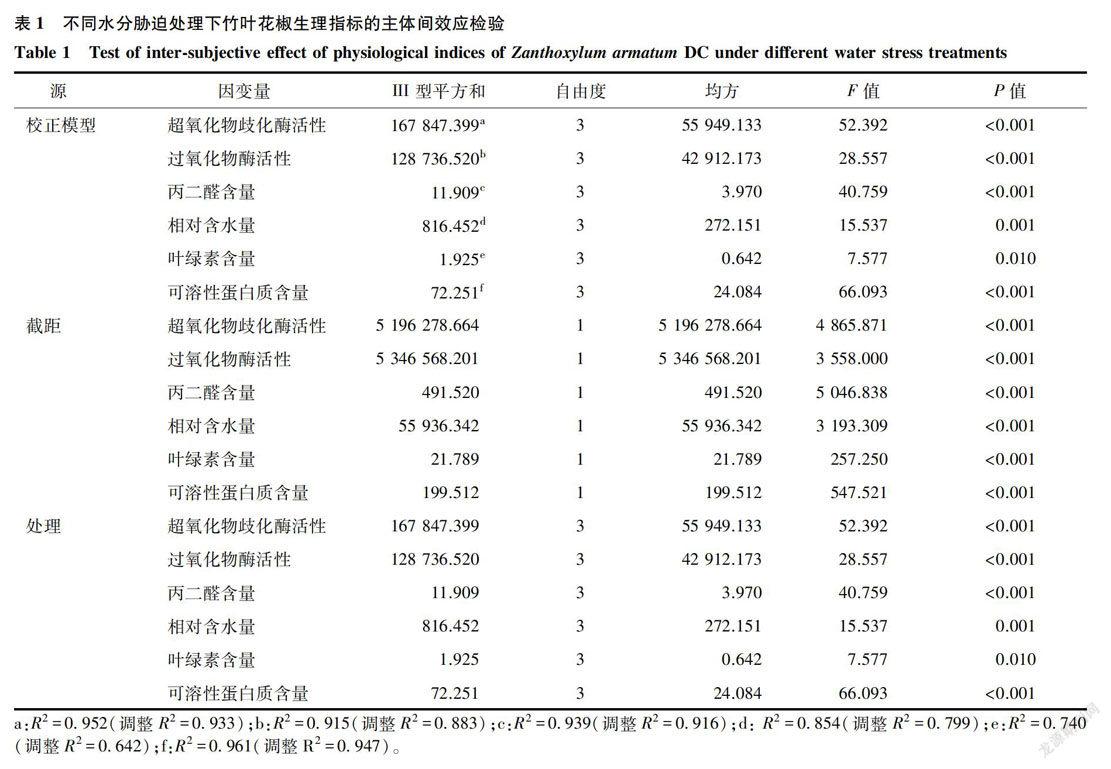
2.1 水分脅迫對竹葉花椒生理指標的影響
由圖1可知,隨著水分脅迫加深,SOD活性先升高后下降,從對照到中度脅迫,SOD活性逐漸增大,之后下降且下降幅度較大。在中度脅迫(T2)下SOD活性達到最高值785.03 U/(g.h),在重度脅迫(T3)下SOD活性達到最低值481. 97U/(g.h),與對照相比分別增加和降低26.44%、22.38%。輕度脅迫(Tl)和T2處理之間無顯著差異,它們與CK、T3之間均存在顯著差異。隨水分脅迫加深,POD活性呈現逐漸升高的變化趨勢。重度脅迫下POD活性達到762.75U/(g.min),與CK相比增加53.83%。隨著土壤水分含量的降低,MDA含量逐漸增加。脅迫初期MDA含量增加速率較快,隨著脅迫程度的增加,MDA含量增加趨勢有所減緩,重度脅迫下丙二醛含量為7. 48 μmol/g,與CK相比增加54.55%。在水分脅迫過程中,SOD、POD活性雖有所增大,但并不能完全清除細胞組織內積累的自由基和過氧化物,因此在水分脅迫逐漸加大時MDA含量逐漸增加,但速率下降。
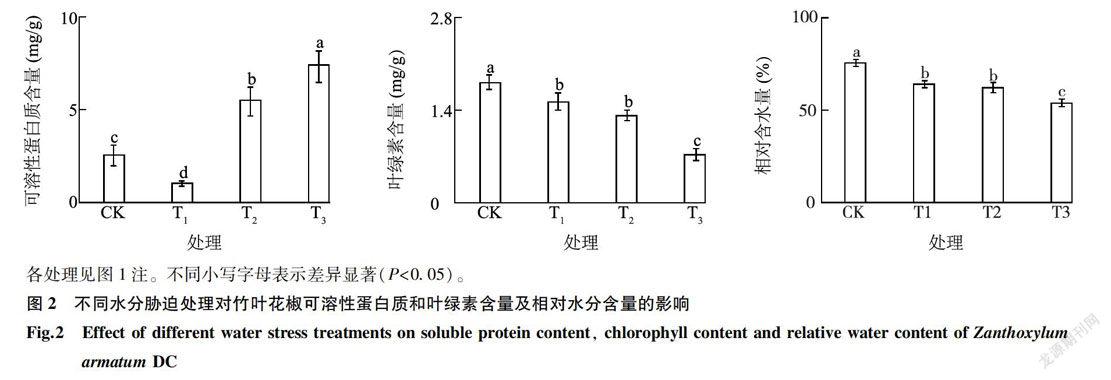
隨著水分脅迫程度逐漸增加,可溶性蛋白質含量呈現增加趨勢(圖2)。從對照到輕度脅迫過程中,可溶性蛋白質含量下降;從輕度脅迫到重度脅迫過程中,可溶性蛋白質含量逐漸增加,且增加幅度較大。其原因可能是在脅迫初期植物體內正常的蛋白質合成被抑制,隨著水分脅迫加劇進而誘導植物自身合成新的蛋白質,導致可溶性蛋白質含量增加。在水分脅迫過程中,葉綠素含量逐漸降低。脅迫初期,葉綠素含量下降幅度較小,差異不顯著。重度脅迫下葉綠素含量顯著下降至1 mg/g以下,原因可能是水分脅迫加速了葉綠體的降解,并且進一步抑制了葉綠素合成酶的活性,導致葉綠素合成量下降。花椒植株受到水分脅迫時,葉片相對含水量逐漸降低,對照花椒葉片相對含水量為80. 69%,T1、T2及T3處理葉片相對含水量分別為68.41%、66.43%、57.57%,與對照相比,葉片相對含水量分別下降17.95%,21.47%、40.16%,差異均達到顯著水平。
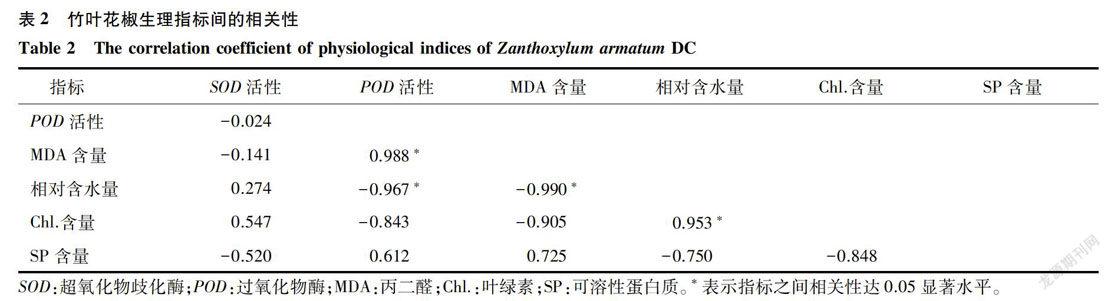
2.2 竹葉花椒生理指標間的相關性分析
由表1可知,不同處理對竹葉花椒葉綠素含量的影響達顯著水平,對其余5個生理指標的影響達極顯著性水平。由表2可知,相對含水量(RWC)與POD活性、MDA含量呈顯著負相關關系,相關系數分別為-0.967*、-0.990*( P
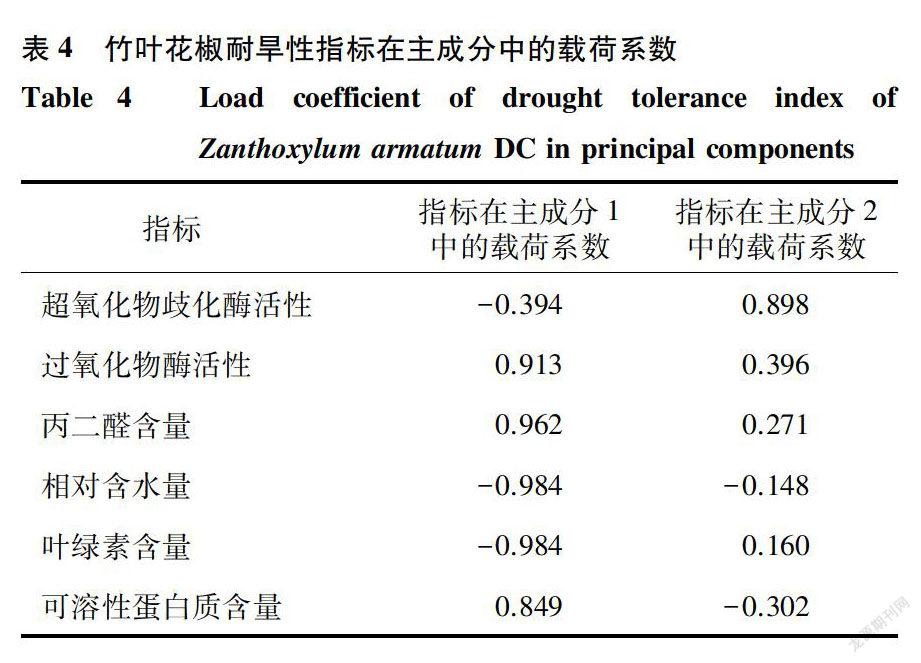
對竹葉花椒植株SOD活性、POD活性、MDA含量等6項生理指標進行主成分分析。結果(表3)表明,特征值大于l的主成分有2個,這2個主成分的累積貢獻率達到95.784%。依據各生理指標在主成分1及主成分2中載荷系數絕對值大小,判斷主成分包含的生理指標。第一主成分在POD活性、MDA含量、相對含水量、葉綠素含量、可溶性蛋白質含量5項指標上有較大的載荷系數,因此該主成分與這5項指標有較強的相關性:第二主成分在SOD活性上有較大的載荷系數,該主成分與SOD活性有較強的相關性(表4)。可見,POD活性、MDA含量、相對含水量、葉綠素含量、可溶性蛋白質含量這5項生理指標貢獻率最大,其綜合作用結果對竹葉花椒耐旱性的影響顯著,可作為評價其耐旱性指標。胡曼筠等[16]對華中冬青相對電導率、丙二醛含量、可溶性蛋白質含量、可溶性糖含量、游離脯氨酸含量、超氧化物歧化酶活性、葉綠素含量及類胡蘿卜素含量8項生理指標進行主成分分析,結果表明相對電導率、丙二醛含量、可溶性蛋白質含量、超氧化物歧化酶活性、葉綠素含量5項指標與華中冬青抗逆性關系密切。這與本研究結果相似,但又有所不同,可能與植物種類不同有關。
3 討論
植物處于正常環境之中時,其細胞內自由基的產生和清除處于動態平衡狀態,然而植物一旦受到不良環境侵害,這種動態平衡就會被破壞,自由基大量積累,細胞代謝紊亂,導致植物遭到傷害,而此時植物體內會迅速啟動保護系統,保護酶活性被激發,催化新陳代謝產生的超氧陰離子自由基歧化為02和H202,進而緩解因不良環境脅迫而引起的細胞傷害[17-18]。本研究中,水分脅迫下SOD活性呈先升后降的趨勢變化,這與張翠梅等[19]以及賈鑫等[20]的研究結果一致。SOD活性的這種變化說明竹葉花椒體內防御機制被刺激,抗性生理活動加快,SOD活性升高以應對體內超氧陰離子的增加以免細胞受損,脅迫后期可能是超出了SOD歧化能力,SOD活性下降;POD活性緩慢升高,這與劉世鵬等[21]在棗樹上的研究結果相似。說明干旱脅迫發生時,竹葉花椒通過及時提高抗氧化酶活性積極適應外界環境變化,然而不同抗氧化酶的響應速度不盡相同,SOD反應迅速,其酶活性增加較快,而POD活性則是穩步上升。干旱脅迫可破壞類囊體膜結構,導致竹葉花椒葉片產生更多自由基(02和H202),使膜脂中不飽和脂肪酸過氧化,MDA等過氧化物質積累。張翠梅等[19]及潘昕等[22]的研究結果表明,水分脅迫能引起植物細胞膜破壞,導致MDA含量增加。
滲透調節物質的積累是植物適應干旱脅迫的主要方式之一。大量研究結果顯示可溶性蛋白質含量與植物抗旱性呈正相關關系。本研究中,可溶性蛋白含量在脅迫前期下降,在中度和重度脅迫下,其含量逐漸升高,這與張慶華等[9]研究結果存在差異。脅迫前期可溶性蛋白含量下降的原因可能是輕度干旱脅迫下蛋白質的分解速率大于合成速率。中度和重度脅迫下可溶性蛋白質含量增加意味著在干旱響應中有新的蛋白質合成[23]。
干旱脅迫不僅會對竹葉花椒生長產生不利影響,還會對光合色素造成損害.進而影響光合能力[24]。目前關于逆境脅迫下葉綠素含量變化的研究結果尚存在一定差異。本研究中隨水分脅迫加深,葉綠素含量逐漸降低,這與李陽等[25]研究結果有一定差異。葉綠素含量下降的原因可能是干旱脅迫下,在類囊體膜上產生較多的活性氧簇( ROS),這些活性氧簇可破壞光合色素。
植物葉片中相對含水量是植物重要的耐旱性能指標,在一定程度上反映葉片持水保水能力[23]。隨水分脅迫加深,竹葉花椒葉片相對含水量呈逐漸下降趨勢,且不同脅迫程度之間存在顯著差異,與張寅媛等[8]研究結果稍有差異,這可能與葉片結構不同有關。
綜上所述,水分脅迫下,竹葉花椒生理指標表現出不同的生理響應,不同脅迫程度之間呈現顯著差異。脅迫初期竹葉花椒通過提高SOD、POD活性增強植株耐旱性能,合成新的可溶性蛋白質適應外界環境變化。水分脅迫引發細胞膜系統發生膜脂過氧化,導致MDA含量增加,引起葉片相對含水量降低,破壞葉綠素分子結構及葉綠體膜系統和光合反應中心。
參考文獻:
[l]王紀輝,陳應福,侯娜,等,源白不同產地青花椒主要特征品質分析[J].食品工業,2018,39(8):322-325.
[2]王紀輝,梁美,侯娜.青花椒良種黔椒2號的選育及栽培技術[J].南方農業學報,2018,49(7):1383-1388.
[3] 呂小軍,楊途熙,何小紅,等,冬季低溫對花椒抗寒性生理指標的影響[J].西北農業學報,2013,22(7):143-148.
[4] 劉淑明,孫丙寅,劉杜玲.不同種源花椒抗寒性研究[J].西北農業學報,2010,19( 11):119-124.
[5]楊途熙,魏安智,李曉,等,越冬過程中花椒抗寒性與組織水和滲透調節物質的變化[J].植物生理學通訊,2010,46(6):579-582.
[6]楊艷麗,楊勇,李大紅,等,轉桃PpCuZnSOD基因大豆的耐旱性[J].江蘇農業學報,2018,34(5):978-983.
[7]劉杜玲,劉淑明,不同花椒品種抗旱性比較研究[J].干旱地區農業研究,2010,28(6):183-189.
[8]李雪妹,劉暢,單羽,等.PEG預處理對水分脅迫下水稻根系抗氧化酶同T酶及其表達的影響[J].江蘇農業科學,2018, 46(7):54-57.
[9] 張慶華,向發云,曾祥國,等.干旱脅迫下不同品種草莓生理響應及抗旱性評價[J].北方園藝,2018(2):30-37.
[10]李佳,劉立云,李艷,等,保水劑對干旱脅迫檳榔幼苗生理特征的影響[J].南方農業學報,2018,49(1):104-108.
[11]張寅媛,劉英,白龍.干旱脅迫對4種景天科植物生理生化指標的影響[J].草業科學,2014,31(4):724-731.
[12]胡宏遠,王振平.水分脅迫對赤霞珠葡萄光合特性的影響[J].節水灌溉,2016(2):18-22,27.
[13]曹巖坡,戴素英,代鵬,等,水分脅迫對不同抗旱性豇豆生長及光合特性的影響[J].河北農業科學,2018,22(1):11-14.
[14]王玉麗,孫居文,荀守華,等,干旱脅迫對東岳紅光合特性、葉綠素熒光參數及葉片相對含水量的影響[J].山東農業科學,2017,49(4):46-50.
[15]高俊鳳,孫群,曹翠玲.植物生理學實驗指導[M].西安:陜西科技出版社,2006:29-30.
[16]胡曼筠,金曉玲,曾雯,等,華中冬青雌雄株抗寒生理指標篩選及評價[J].中南林業科技大學學報,2018,38(5):59-64.
[17] CONCEICAO V S.Regulation of chlorophyll biosynthesis and deg-radation by salt stress in sunflower levaes[J].ScientiaHorticulture ,2004, 103:93-99.
[18] JANDA T,KSA E,SZALAI G,et al.lnvestigation of antioxidantactivity in mize during low temperature stress[J].Acta BiologicaSzegedrensis, 2005 ,49( 1/2): 53-54.
[19]張翠梅,師尚禮,吳芳,干旱脅迫對不同抗旱性苜蓿品種根系生長及生理特性影響[J].中國農業科學,2018,51(5):868-882。
[20]賈鑫,孫窗舒,李光躍,等,干旱脅迫對蒙古黃芪生長和生理生化指標及其黃芪甲苷積累的影響[J].西北植物學報,2018,38(3):501-509.
[21]劉世鵬,劉濟明,陳宗禮,等.模擬干旱脅迫對棗樹幼苗的抗氧化系統和滲透調節的影響[J].西北植物學報,2006,26(9): 1781-1787.
[22]潘昕,邱權,李吉躍,等.干旱脅迫對青藏高原6種植物生理指標的影響[J].生態學報,2014,34( 13):3558-3567.
[23]馮曉敏,張永清.水分脅迫對糜子植株苗期生長和光合特性的影響[J].作物學報,2012,38(8):1513-1521.
[24]王婷,海梅榮,羅海琴,等.水分脅迫對馬鈴薯光合生理特性和產量的影響[J].云南農業大學學報,2010,25(5):737-742.
[25]李陽,齊曼·尤努斯,祝燕,水分脅迫對大果沙棗光合特性及生物量分配的影響[J].西北植物學報,2006,26( 12): 2493 -2499.
(責任編輯:張震林)
