小肽螯合鐵緩解櫻桃蘿卜苗期高溫脅迫的生理機制
2019-09-12 01:34:26賈春花張林琳劉之廣孫玲麗
植物營養與肥料學報 2019年8期
賈春花,張林琳,張 民,*,劉之廣*,鄭 磊,孫玲麗,孫 正
[1 土肥資源高效利用國家工程實驗室/山東農業大學資源與環境學院,山東泰安 271018;2 養分資源高效開發與綜合利用國家重點實驗室/金正大生態工程集團股份有限公司,山東臨沭 276700;3 眾德肥料 (平原) 有限公司,山東平原 253100;4 中國海洋大學,山東青島 266005]
櫻桃蘿卜 (Raphanus sativusL. var.radculusPers.)是中國主要的四季蘿卜之一,生育期短,外形、色澤美觀,口感爽脆,營養豐富,單位面積產量高,經濟效益好。櫻桃蘿卜最適生長溫度為5~25℃,不耐炎熱,在高溫環境中生長受阻、易發生病蟲害,產量低、品質差。農業生產中,櫻桃蘿卜溫室栽培極易受高溫脅迫。高溫脅迫會改變植物細胞膜結構,使膜上不飽和脂肪酸含量增加、蛋白質失活,進而損傷磷脂雙分子層流動性,影響膜脂的三級及四級結構等[1]。另外,高溫脅迫可破壞葉綠體中的電子運輸,改變能量分布,影響與碳代謝相關酶的活性,尤其是Rubisco酶[2],同時引起活性氧爆發[3-4],導致膜脂過氧化反應和電解質外滲,影響植物正常的光合作用和生長發育[5]。因此,探究可提高櫻桃蘿卜耐熱性的生物刺激素并研究其抗熱機理,對實現櫻桃蘿卜的優質生產與均衡供應、緩解夏季果蔬供應不足具有重要的現實意義[6]。前人研究表明,小肽螯合鐵是一種來源于玉米蛋白的新型生物刺激素,有效成分為仿生內源肽及有機螯合鐵,其雙分子螯合結構與作物體內酶的活性中心鐵的存在形式類似。有研究表明,玉米肽具有良好的清除羥基自由基和DPPH自由基的能力及還原能力[7-8],可顯著提高作物養分吸收和利用效率,提高作物抗氧化能力[9]。而蕓苔素內酯是一種常見的含催化平衡成分和氨基酸類化合物的生物刺激素,具有提高作物抗逆性、改良農產品品質和增加產量等作用,已獲得市場的認可并廣泛應用[10]。徐靜等[11]研究表明,噴施30~60 mg/kg的6.003%氨基·蕓苔素內酯水劑可顯著提高芒果產量及品質,應用前景廣闊。目前關于小肽螯合鐵對高溫脅迫下作物耐熱性調控方面的研究尚未見報道。因此本研究以蕓苔素內酯為陽性對照,研究高溫脅迫下小肽螯合鐵對櫻桃蘿卜內源激素水平、抗氧化系統及光合系統的影響,探討小肽螯合鐵對櫻桃蘿卜抗氧化系統影響的生理機制及其對內源激素的調控作用,為緩解櫻桃蘿卜高溫脅迫提供理論依據。
1 材料與方法
1.1 試驗材料
試驗在山東省泰安市山東農業大學土肥資源高效利用國家工程實驗室進行。
供試櫻桃蘿卜種子來源于荷蘭BEJO公司,生長期45天。
供試生物刺激素為蕓苔素內酯粉劑,干物質量為0.136%,來源于上海拜諾國際生物科技進出口公司;小肽螯合鐵 (結構式如圖1) 來源于山西北正農生物工程有限公司,制備過程為將玉米蛋白通過酶解獲取含甘氨酸和蛋氨酸的肽片段,滅活除雜后過分子篩獲取含1000~1100分子量的肽類物質,經其與鐵元素螯合濃縮后制得水劑成品,干物質量為80 g/L。
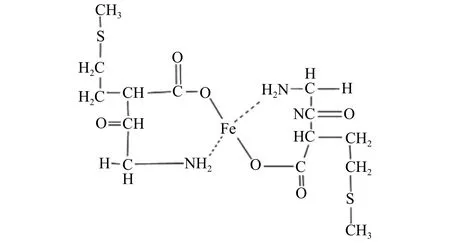
圖 1 小肽螯合鐵結構式Fig. 1 Structural formula of peptide-chelated iron
供試基質為蔬菜育苗基質,來源于丹麥品氏托普公司,基本理化性質為有機質434 g/kg、NO3--N 296 mg/kg、NH4+-N 84.1 mg/kg、有效磷99.2 mg/kg、速效鉀1169 mg/kg、pH值5.94(水基質比5∶1)、電導率 2180 μS/cm(水基質比 5∶1)。
供試育苗采用梯形方盒,上口邊長10 cm、高8 cm、底面邊長7 cm。自配營養液為氮、磷、鉀比例為20∶10∶20的育苗專用肥1000倍的水溶液[12]。
1.2 試驗設計
試驗共設6個處理 (表1),3次重復,置于光照培養箱 (寧波江南儀器廠) 內,晝夜溫度設置為(30 ±1)℃,常溫對照溫度為(25 ± 1)℃,相對濕度通過加濕器維持在50%~60%,光照強度約15000 lx,每天設定光照16 h[12]。
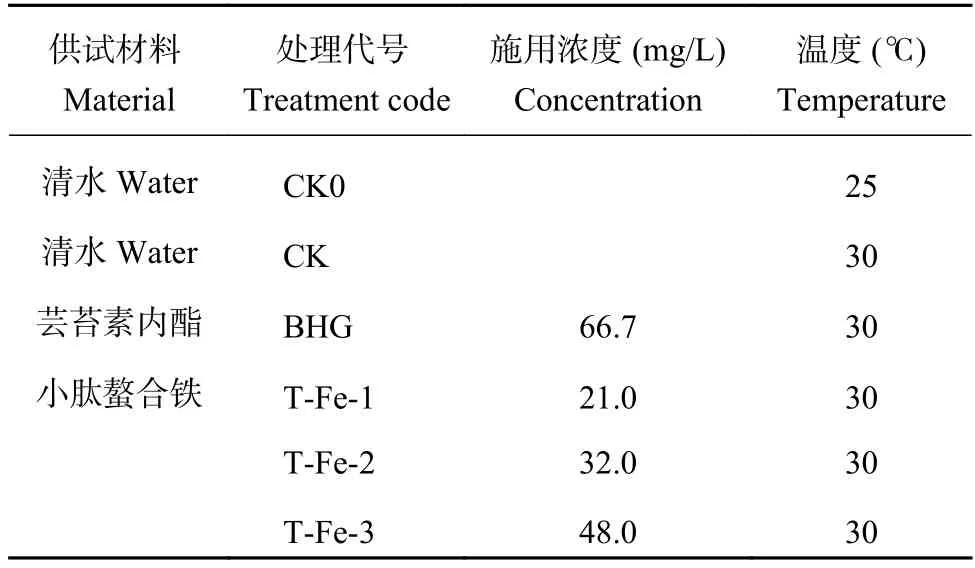
表1 試驗處理Table 1 Treatments for experiment
試驗于2018年3月20日至4月17日進行。浸種時選取大小一致、籽粒飽滿的蘿卜種子,分別置于不同濃度的兩種供試生物刺激素稀釋液中 (表1),于25℃恒溫培養5小時。浸種結束后于30℃恒溫培養箱進行培養,并設置25℃常溫空白對照。分別播種2粒櫻桃蘿卜種子于裝滿供試基質的育苗方盒中,埋深0.3 cm,播種后第5天 (2018年3月25日)間苗至一株,配置營養液與5倍濃度的生物刺激素混合液,播種后于第8天 (2018年3月28日) 和16天 (2018年4月5日),分別用混合液50 mL進行灌根,培養28天后測定各項指標。
1.3 樣品分析方法
分別測定每個處理3株生物量、根長度;生長素 (IAA)、脫落酸 (ABA) 的測定采用HPLC測定內源激素法[13];脯氨酸 (Pro) 含量、抗壞血酸過氧化物酶 (APX) 活性、谷胱甘肽還原酶 (GR) 活性、過氧化氫 (H2O2)、超氧陰離子 ()、Rubisco酶活性的測定采用酶聯免疫法,參照靳德明等的報道[14]。超氧化物歧化酶 (SOD) 活性、過氧化氫酶 (CAT) 活性、丙二醛 (MDA) 含量的測定用李琲琲等[15]的研究方法;葉片凈光合速率 (Pn)、氣孔導度 (Gs)、胞間CO2濃度(Ci) 和蒸騰速率 (Tr) 采用LI-6400便攜式光合測定儀測定。
1.4 數據分析方法
分別采用Office 2016和SAS 8.2軟件進行ANOVA方差分析及Duncan差異顯著性檢驗完成;用Excel 2016軟件進行作圖。
2 結果與分析
2.1 小肽螯合鐵對櫻桃蘿卜生物量及根長的影響
生物量和根長分別反映了櫻桃蘿卜的營養生長狀況及其從基質中吸收養分的能力。與CK0(清水25℃) 處理相比,T-Fe-1(21.0 mg/L小肽螯合鐵) 處理的整株生物量差異不顯著,T-Fe-2、T-Fe-3(32.0、48.0 mg/L小肽螯合鐵) 處理顯著降低;T-Fe-1與CK(清水30℃) 處理相比,整株生物量顯著提高26.0%,與BHG(66.7 mg/L蕓苔素內酯) 處理相比差異不顯著,而T-Fe-2處理的整株生物量與T-Fe-1和T-Fe-3處理相比顯著降低 (表2)。
與CK0處理相比,小肽螯合鐵處理顯著提高櫻桃蘿卜的根長;與CK處理相比,T-Fe-1處理根長顯著提高35.8%,T-Fe-1處理與BHG處理差異不顯著 (表2)。高溫下T-Fe-1處理的整株生物量和根長與CK相比均顯著提高,說明其添加對高溫下櫻桃蘿卜生長有一定的促進作用。
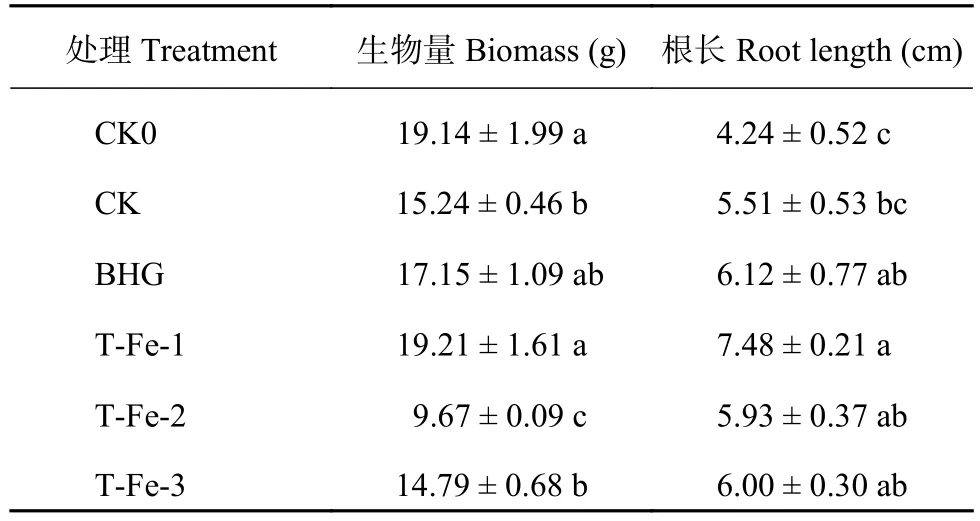
表2 不同處理的櫻桃蘿卜整株生物量及根長Table 2 Biomass and root length of cherry radish under different treatments
2.2 小肽螯合鐵對櫻桃蘿卜葉片內源激素及脯氨酸含量的影響
2.2.1 小肽螯合鐵對櫻桃蘿卜葉片內源激素含量的影響 圖2顯示,內源激素ABA和IAA在植物適應逆境的生理過程中起重要作用。與CK0處理相比,T-Fe-1、T-Fe-2處理的IAA含量均顯著提高。而TFe-1、T-Fe-2、T-Fe-3處理較CK處理分別顯著提高了144%、65.9%、26.1%,說明小肽螯合鐵對櫻桃蘿卜的生長發育起到較好的促進效果。與CK0處理相比,T-Fe處理的櫻桃蘿卜體內ABA含量差異不顯著;與CK處理相比,T-Fe處理的ABA含量顯著降低。
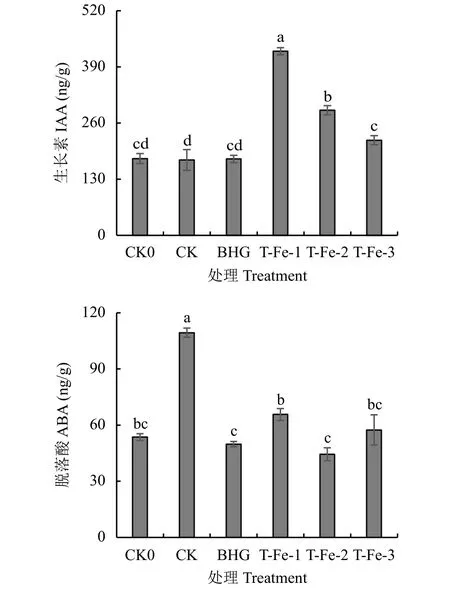
圖 2 不同處理櫻桃蘿卜葉片生長素和脫落酸含量Fig. 2 IAA and ABA content in cherry radish leaves in each treatment
2.2.2 小肽螯合鐵對櫻桃蘿卜葉片脯氨酸含量的影響正常生長條件下作物滲透調節物質Pro含量通常較低,而在脅迫條件下表現為上升。T-Fe處理的櫻桃蘿卜葉片Pro含量與CK0、CK相比均顯著上升,與CK處理相比,T-Fe-1處理的Pro含量顯著提高229.9%(圖3),說明高溫脅迫下添加適宜濃度的小肽螯合鐵對櫻桃蘿卜滲透調節系統均具有顯著的改善效果且效果優于蕓苔素內酯處理。
2.3 小肽螯合鐵對櫻桃蘿卜葉片抗氧化系統及膜脂過氧化程度的影響
2.3.1 小肽螯合鐵對櫻桃蘿卜葉片抗氧化酶活性的影響 表3表明,與CK處理相比,T-Fe-1處理的SOD活性無顯著差異,但隨著小肽螯合鐵濃度增加,對SOD活性的促進作用逐漸加強。T-Fe-1處理CAT活性與CK差異不顯著,但較BHG處理顯著提高262%。高溫處理下APX活性相比CK0均顯著提高 (表3),與CK處理相比,T-Fe-1處理的APX活性顯著提高21.7%。與CK0、CK相比,高濃度的TFe處理的GR活性顯著提高 (表3)。說明高溫脅迫
下,添加小肽螯合鐵處理可顯著提高APX、GR活性進而提高櫻桃蘿卜的抗逆性。
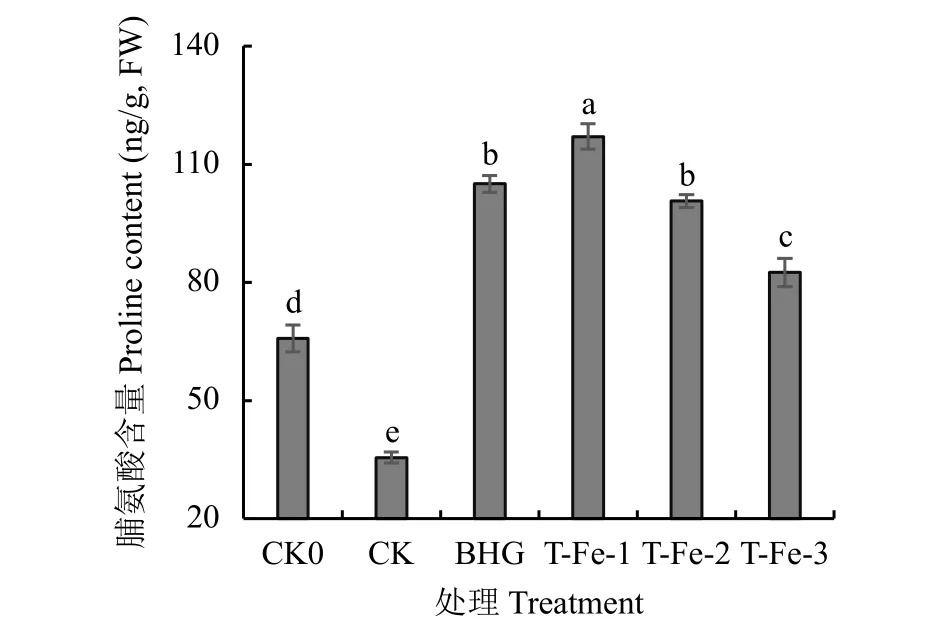
圖 3 不同處理櫻桃蘿卜葉片脯氨酸含量Fig. 3 Proline content in cherry radish leavesin each treatment
2.3.2 小肽螯合鐵對櫻桃蘿卜葉片膜脂過氧化程度的影響 高溫脅迫下,細胞膜上不飽和脂肪酸在活性氧自由基如和H2O2的毒害作用下易發生過氧化,生成MDA,與酶蛋白結合并發生鏈式反應,使酶蛋白失活,此過程稱為膜脂過氧化[16],而MDA是膜脂過氧化的標志性物質,其含量反映細胞膜的膜脂過氧化程度以及細胞膜受損程度。與CK相比,TFe-1處理的MDA、H2O2含量和活性無顯著變化,而T-Fe-2處理的含量均顯著降低 (表4),說明適宜濃度的T-Fe處理對櫻桃蘿卜細胞膜有保護作用。
2.4 小肽螯合鐵對櫻桃蘿卜葉片光合特性及其調節系統的影響
2.4.1 小肽螯合鐵對櫻桃蘿卜葉片光合特性的影響
表5表明,高溫脅迫嚴重影響了櫻桃蘿卜葉片的凈光合速率,與CK0處理相比,T-Fe-3處理櫻桃蘿卜的凈光合速率 (Pn) 無顯著差異,其它各處理均顯著降低。T-Fe-3處理櫻桃蘿卜葉片的凈光合速率較CK處理顯著提高31.5%。說明在本試驗條件下,48.0 mg/L小肽螯合鐵有利于提高櫻桃蘿卜的耐高溫脅迫能力,進而保證了植株葉片的凈光合速率保持在正常水平。
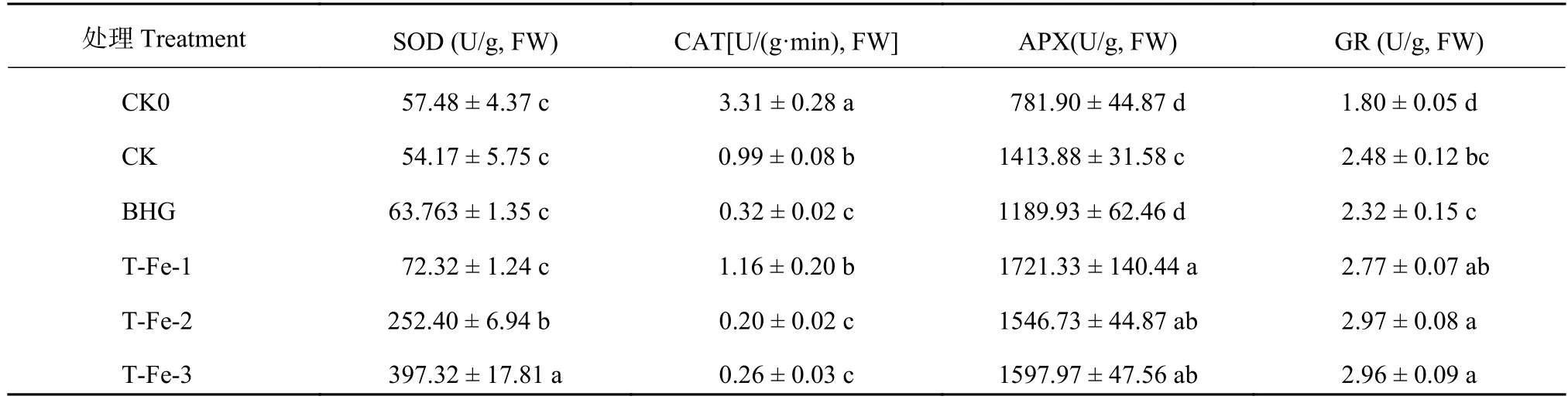
表3 不同處理的櫻桃蘿卜葉片抗氧化酶活性Table 3 Antioxidase activity of cherry radish leaves with different treatments
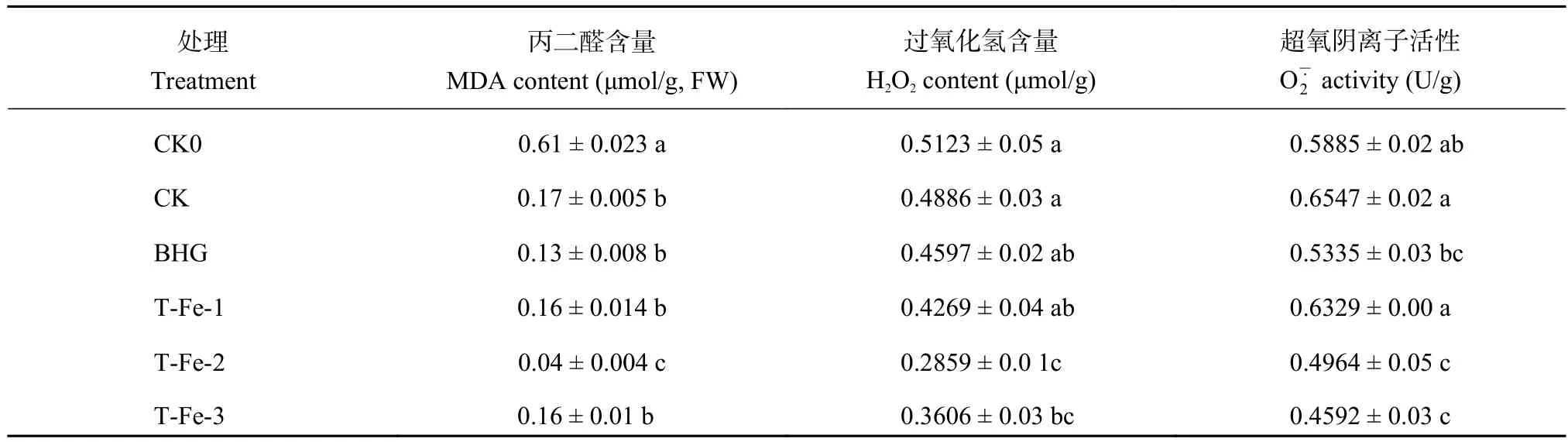
表4 不同處理的櫻桃蘿卜葉片丙二醛、過氧化氫含量和超氧陰離子活性Table 4 MDA, H2O2 content and O2- activity of cherry radish leaves with different treatments
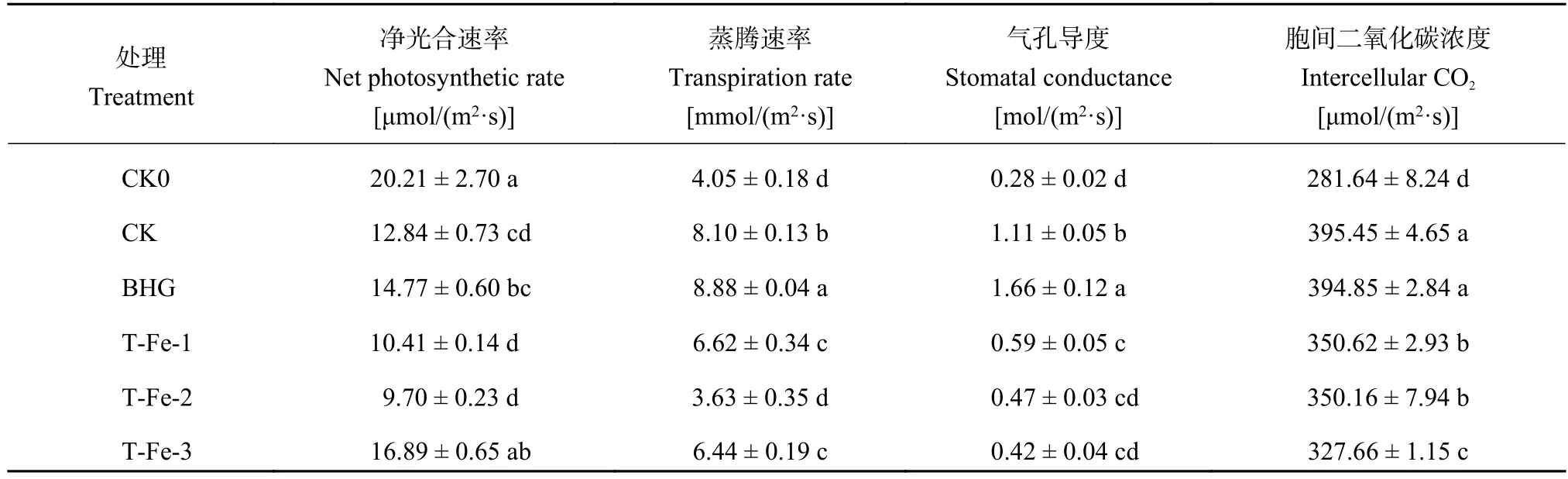
表5 不同處理的櫻桃蘿卜葉片光合特性Table 5 Photosynthetic characteristic of cherry radish leaves with different treatments
蒸騰速率 (Tr) 在一定程度上能夠反映作物調節水分運動的水平以及適應逆境的能力[17]。與CK0相比,高溫脅迫下添加各生物刺激素處理的櫻桃蘿卜Tr顯著提高;與CK相比,小肽螯合鐵處理的櫻桃蘿卜Tr顯著降低18.3%~55.2%(表5),而蕓苔素內酯處理顯著提高,添加小肽螯合鐵可降低櫻桃蘿卜Tr,從而避免大量水分散失來抵御高溫脅迫。與CK0處理相比,高溫脅迫下,CK、BHG及T-Fe-1處理的氣孔導度 (Gs) 與胞間二氧化碳濃度 (Ci) 均提高;與CK相比,T-Fe處理的Gs與Ci均顯著降低。其中T-Fe-1處理的Gs和Ci分別顯著降低46.8%和11.3%。說明高溫脅迫下,葉片可通過迫使部分氣孔關閉進而減少胞間二氧化碳濃度及減少水分散失。2.4.2 小肽螯合鐵對櫻桃蘿卜葉片1, 5-二磷酸核酮糖羧化酶 (Rubisco) 的影響 葉片Rubisco酶在光合碳同化過程中起關鍵作用,參與光合作用和光呼吸作用過程并調節兩者之間的關系,其活性可以反映光合作用暗反應的效率[18]。與CK處理相比,小肽螯合鐵處理差異均不顯著 (圖4),但T-Fe-3處理與BHG處理相比顯著降低,這可能是由于高濃度的小肽螯合鐵降低了櫻桃蘿卜胞間二氧化碳濃度,使Rubisco酶與二氧化碳的親和力降低進而影響酶活性導致的。
3 討論
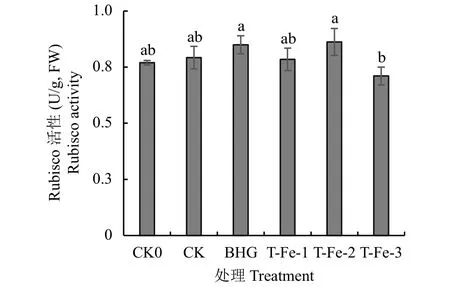
圖 4 不同處理櫻桃蘿卜葉片的Rubisco酶活性Fig. 4 Rubisco activity of cherry radish leaves with different treatments
生長素IAA是作物體內一種重要的內源激素,參與生長發育的各個過程,通過測定生長素的含量可推測作物生長態勢[19]。脫落酸是一種典型的脅迫激素,前人研究認為高溫脅迫時作物體內脫落酸積累有利于誘導作物耐熱性的提高[20]。王日明等[21]研究表明,高溫脅迫下維持較高的IAA含量有助于降低自由水的含量,延緩葉片的枯黃衰老和維持細胞膜的穩定性,從而提高作物耐高溫能力。本試驗結果表明,高溫下添加小肽螯合鐵處理的IAA含量與30℃對照相比顯著增加,這與王日明等的研究結果一致,說明添加小肽螯合鐵能提高櫻桃蘿卜在高溫脅迫下的長勢。而本試驗中小肽螯合鐵處理的ABA含量與30℃對照相比顯著降低,可能是由于高溫脅迫造成作物體內ABA累積,因此30℃對照處理的ABA含量較高,但作物體內ABA過多會抑制細胞分裂與伸長,通過生物量分析,T-Fe處理的ABA 含量對細胞生長發育無抑制作用,反而抵御高溫脅迫對櫻桃蘿卜的傷害,是較適合高溫下作物生長的含量水平。
高溫脅迫誘導植物活性氧爆發導致生物膜降解,影響植物正常生長發育,作物可通過抗壞血酸—谷胱甘肽循環及谷胱甘肽過氧化物酶循環來緩解脅迫。其中,SOD、CAT和 APX是植物體內酶促防御系統的重要保護酶,它們能有效分解體內活性氧自由基,從而使生物體免受傷害[22]。本試驗結果表明,T-Fe-1處理的GR、APX活性與30℃對照相比均顯著提高。GR可直接促進還原型輔酶II及谷胱甘肽的生成,從而促進谷胱甘肽過氧化物酶循環,緩解了H2O2的毒害;APX活性的提高可直接促進單脫氫抗壞血酸 (MDHA) 氧化生成抗壞血酸 (AsA),能夠維持AsA-GSH循環快速有效進行,保證了GSH和AsA的再生,從而有效抑制H2O2的增加[15],抵御高溫脅迫。MDA、、H2O2作為衡量膜脂過氧化程度的關鍵指標,其含量的減少也證實了抗氧化系統在清除活性氧抵御高溫脅迫過程中的關鍵作用。
光合作用是一切生物直接或間接的能量來源,是植物進行物質轉換和能量代謝的關鍵[23],有研究表明多數植物在短時間高溫脅迫下,光合作用就會受到明顯抑制[24]。本試驗結果表明,在高溫脅迫下,與非脅迫CK0處理相比,各處理Pn均顯著降低,表明高溫對櫻桃蘿卜的光合作用產生了明顯的抑制;TFe-1處理的Pn與CK處理差異不顯著,Gs和Ci顯著降低,T-Fe-3處理的Pn較CK處理顯著提高,而T-Fe-1處理的整株生物量和根長顯著高于T-Fe-3處理。童淑媛等[25]研究表明,保持綠色雖然可提高光的俘獲,但它并不總是與干物質和谷物產量增加有關。Thomas等[26]描述了4種類型的保持綠色,其中有一種定義為裝飾性的持綠,其具有葉片保持色素沉淀的特點,但光合作用下降。因此,造成T-Fe-1處理Pn降低而整株生物量提高的原因可能是光的俘獲低而利用率高。與CK0處理相比,高溫脅迫下Tr升高可降低葉片溫度,而T-Fe-1處理較CK處理Tr降低又可減少葉片失水,維持櫻桃蘿卜在高溫下正常生長。Pro作為滲透調節系統的重要物質,能夠通過促進蛋白質的水合作用使蛋白質膠體親水面積增大,降低細胞水勢來提高作物的適應性與抗逆性[27],對氧化還原電勢的變動起到緩沖作用[4]。本研究中,高溫脅迫下Gs的降低導致了呼吸作用減弱,進而Ci降低引起細胞內滲透調節系統的變化,為維持細胞體系的平衡,滲透調節物質Pro迅速合成,保障了細胞生命活動正常進行,這可能是高溫脅迫下Rubisco酶活性無顯著變化的原因。
本試驗條件下,小肽螯合鐵可提高內源激素含量和活性氧清除系統相關酶活性,及提高作物對高溫脅迫的適應性,降低作物損傷,抗逆促生效果顯著,具有較強的應用價值,與傳統肥料配合可制備抗逆增值型肥料,市場潛力大。然而受到短期培養試驗局限性的影響,仍需針對不同地區及作物特性通過田間定位試驗驗證其效果,同時其抗逆生理生化及分子機制仍需深入探究。
4 結論
30℃條件下,以濃度為21.0 mg/L的小肽螯合鐵處理可顯著提高櫻桃蘿卜整株生物量26.0%,顯著提高生長素含量144%;可分別顯著提高抗壞血酸過氧化物酶、谷胱甘肽還原酶活性21.7%、11.7%,并增強櫻桃蘿卜抗氧化系統活性,抵御高溫脅迫。超過該濃度,則櫻桃蘿卜生長受到抑制。
