ε-聚-L-賴氨酸及其抑菌機理研究進展
2019-09-19 09:00:52李曉玥
作物研究 2019年6期
李曉玥,劉 潭,楊 博
(湖南農業大學生物科學技術學院,長沙410128)
隨著人們對食品安全性的認識和要求日益提高,開發抗菌性強、水溶性好、安全無毒的天然食品防腐劑已成為全世界食品領域研究熱點。ε-聚-L-賴氨酸(ε-Poly-L-lysine,ε-PL)是一種天然生物防腐劑,具有抑菌譜廣、水溶性好、安全性高、熱穩定性好等特點,其優良的防腐性能符合人們對于新型食品防腐劑的要求[1]。ε-PL在日本、韓國等國家已作為食品防腐劑廣泛使用[2]。2004年,其被美國食品和藥品管理局(FDA)批準為安全食品保鮮劑(GRAS)[3]。我國也在2014年正式批準其作為食品添加劑新品種[4]。近年來,ε-PL在食品等領域的應用與發展,激發了研究者對其抑菌作用機理的探尋。本文在介紹ε-PL特性、菌種選育、發酵生產和應用的基礎上,對其抑菌作用機理進行了綜述。
1 ε-PL的特性
ε-PL通常由25~35個L-賴氨酸通過ε-氨基和α-羧基脫水縮合形成酰胺鍵聚合而成,是一種同型單體多肽聚合物。ε-PL的結構如圖1。ε-PL的水溶性和熱穩定性良好,其水溶液在120℃高壓滅菌處理20 min不會發生分解作用[5]。ε-PL分子量一般為2.5~4.5 kDa,為白色或淡黃色粉末,吸濕性較強,略帶苦味,微溶于乙醇,不溶于乙酸乙酯、乙醚等有機溶劑[6~8]。由于ε-PL是混合物,所以沒有固定的熔點,250℃開始軟化分解[9]。紅外光譜分析表明,ε-PL在1680~1640 cm-1和1580~1520 cm-1有強吸收峰[10]。
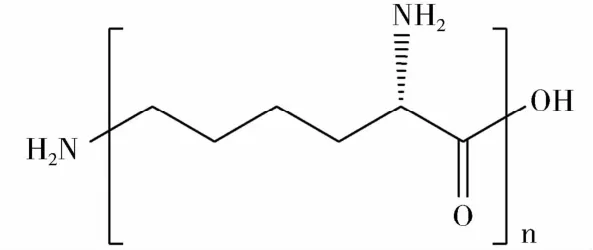
圖1 ε-PL的結構
2 ε-PL產生菌的選育
ε-PL的等電點約為9.0,當環境溶液pH<9.0時,其呈酸性帶正電荷,這為ε-PL菌株的篩選提供了便利[11]。在篩菌時,菌株分泌的帶正電荷的ε-PL可與帶相同電荷的瓊脂或堿性染料分子(如亞甲基藍等)產生靜電斥力,從而在菌株周圍形成肉眼可見的透明圈。同時,屬于生物堿的ε-PL還可與Dragendorff或Wagner試劑發生顯色反應,因此可簡便快速地從環境中篩選出ε-PL產生菌[12]。近年來,國內外學者進行了大量的工作以獲取菌種種質資源,分離得到10余種ε-PL產生菌,經形態學、理化性質、16SrDNA測序并進行系統進化樹分析等鑒定工作,確定這些來源菌株屬于鏈霉菌屬、北里孢菌屬、麥角菌屬等幾類微生物[13]。現今ε-PL的發酵生產通常采用鏈霉菌屬中的白色鏈霉菌(Streptomyces albulus)。
由于從自然界中篩選的ε-PL產生菌的生長速率和ε-PL產量較低,難以滿足生產的需求,故研究者們對ε-PL產生菌進行了大量的育種工作,以提高野生菌的ε-PL產量。鑒于ε-PL產生菌的遺傳背景較復雜且尚未揭示清楚,目前關于ε-PL產生菌的育種工作多采用誘變育種的方式進行。Hiraki等[14]利用亞硝基胍(NTG)誘變,并在培養基中加入S-(2-氨基乙基)-L-半胱氨酸(AEC)和甘氨酸進行脅迫,在經過長達20次的誘變后,獲得了一株ε-PL產量達2.11 g/L的突變株。陳瑋瑋等[15]使用硫酸二乙酯(DES)對北里孢菌PL6-3進行化學誘變,并在培養基中加入AEC,獲得了一株突變株MY5-36,經搖瓶發酵后其ε-PL產量達到1.17 g/L,是出發菌株的3倍,且經5 L發酵罐分批發酵后的ε-PL產量高達7.72 g/L,較出發菌株提高了7倍以上。李南臻等[16]利用60Co和常壓室溫等離子體(ARTP)誘變法對白色鏈霉菌H菌株進行復合誘變,最終選育出一株高產突變株H1-2,經10 L發酵罐發酵后的ε-PL產量高達24.50 g/L,較出發菌株提高了88.5%。此外,核糖體工程、基因工程等新興技術的發展,加快了育種周期。吳光耀等[17]利用核糖體工程育種方法,成功地將ε-PL的產量提高至3.70 g/L。Li等[18]發現,通過基因組重排技術可以提高菌體對葡萄糖的利用率,最終使ε-PL的產量提高了1.88倍。
3 ε-PL的發酵生產
目前,日本等國家通過微生物發酵法已經實現ε-PL的工業化生產,而我國對ε-PL的研究基本還處于實驗階段,僅有浙江新銀象等少數幾家公司進行了商業化應用。關于ε-PL的發酵生產,可通過優化菌種培養基和培養條件的方式提高ε-PL的產量。Shima等[19]利用野生菌S.albulus,探究了不同碳氮源對其代謝產ε-PL的影響,發現葡萄糖或甘油可以作為碳源被利用,而氮源可以是硫酸銨或者是酵母膏。Shih等[20]使用響應面法對S.albulus菌種的培養基成分進行了優化,結果表明對該菌株產ε-PL影響最大的3個因素分別為酵母膏、硫酸銨以及葡萄糖,經優化后,ε-PL產量從0.75 g/L增長到了8.13 g/L,產量提高了將近10.84倍。另外,培養過程中的pH對ε-PL產量也有一定影響。當菌體的生物量積累到一定程度時,培養基的pH值降低到4.2左右對于ε-PL的積累是至關重要的。Kahar等[21]利用S.albulus410菌株建立了pH兩階段調控的發酵工藝,先將pH值控制在5.0以上以積累菌體量,再將pH值控制在有利于菌體代謝產生ε-PL的4.0環境,這使得ε-PL產量大大提高,從原始的5.72 g/L提高到了48.35 g/L。
4 ε-PL的應用
ε-PL作為一種性能優良的天然防腐劑,目前主要作為食品添加劑廣泛應用于米飯、面食、魚類制品等傳統食品的防腐保鮮[22]。在日本,利用ε-PL作為食品防腐劑的商業化應用發展迅速,市場規模早已超過數十億日元。ε-PL作為新型食品防腐劑,不僅可以單獨使用,還可與其他食品添加劑混合使用以提高防腐能力。徐紅華等[23]研究發現,ε-PL和甘氨酸復配能夠延長牛奶的貨架期,保存11 d后仍無變質跡象。ε-PL還可作為藥物及DNA的載體,廣泛應用于醫學和制藥領域。氨甲蝶呤是目前治療白血病和惡性腫瘤最為常用的藥物,它與作為載體的ε-PL相結合,能使靶細胞對它吸收率大大提高[24]。另外,ε-PL為多陽離子聚合物,還能與帶電質粒經靜電相互作用形成一種內含可避免核酸酶降解的基因復合物,從而提升基因DNA藥物的轉移效率[25]。除此之外,ε-PL還可作為吸水材料,與丙烯乙二醇酯結合形成具有高吸水性的水凝膠,該水凝膠對吸水量的反應靈敏度較高,且具有一定的抑菌活性,可廣泛應用于衛生巾及嬰兒紙尿褲等系列產品中[26]。
ε-PL廣泛應用的同時,其安全性也受到考量。Neda等[27]通過小鼠慢性和亞急性灌胃實驗及對ε-PL的毒理學進行監測,發現ε-PL在各種實驗條件下均無任何毒性,且對神經、免疫、生殖系統以及胚胎發育都不會產生毒害。Hiraki等[28]通過放射法研究發現,ε-PL在胃腸道內幾乎不被吸收,7日內檢測到放射性標記的ε-PL經排泄途徑緩慢排出,在機體內不存在累積作用。因此ε-PL作為一種新型的生物防腐劑是安全可靠的。
5 ε-PL的抑菌機理
5.1 ε-PL的抑菌作用
ε-PL具有廣譜抗菌性,且經加熱后也能發揮較好的抑菌效果,并在與其他防腐劑或試劑(如乳酸鏈球菌素、醋酸等)復合使用時可在一定程度上提高抑菌活性。Shima[29]于1984年首次提出了ε-PL的抑菌性,發現ε-PL對革蘭氏陽性細菌、革蘭氏陰性細菌和真菌的生長繁殖均有著顯著的抑制作用。研究表明,ε-PL對能夠引起食物腐敗的多數菌種均有一定的抑制作用,如酵母菌屬中的尖銳假絲酵母、紅發夫酵母,革蘭氏陽性菌中的凝結芽孢桿菌、枯草芽孢桿菌,革蘭氏陰性菌中的產氣節桿菌、大腸桿菌等,故ε-PL的抑菌范圍較廣[30]。ε-PL經80、100、121℃處理一定時間后,其抑菌效果仍基本不變,具有較好的熱穩定性[31]。另外,ε-PL還可與其他添加劑復合使用以增強抑菌效果。據報道,ε-PL與0.1%強醋酸的復合使用后的抑菌效果較單獨使用時明顯增強,其與0.1%甘氨酸的復合抑菌效果次之[31]。
5.2 ε-PL的抑菌機理
目前關于ε-PL抑菌作用機制的研究主要集中在釀酒酵母、大腸桿菌、枯草芽孢桿菌及李斯特菌等菌種,這些菌種分別代表真菌、革蘭氏陰性菌和革蘭氏陽性菌。根據菌種特性的不同,ε-PL的抑菌作用效果及機制有著較明顯的差異。
5.2.1 ε-PL對釀酒酵母的作用機制
ε-PL對真菌的抑菌機理研究,主要以釀酒酵母(Saccharomyces cerevisiae)作為對象進行探究。薄濤等[32]研究發現,ε-PL對釀酒酵母的抑菌作用強弱依賴于ε-PL所添加的濃度。添加低于200μg/mL的ε-PL對釀酒酵母生長有一定的抑制作用,添加200~500μg/mL的ε-PL對釀酒酵母生長有明顯抑制作用,而添加500μg/mL以上的ε-PL則對釀酒酵母生長致死率高達100%。這種與作用濃度相關的現象,可能是ε-PL存在一定的作用閾值濃度。此外,研究發現ε-PL可提高酵母細胞膜的通透性和疏水性。同時,二價陽離子可對ε-PL的抑菌活性產生一定影響。當二價陽離子(如Mg2+、Ca2+等)濃度達到一定值時,可對ε-PL抑菌活性產生削弱作用。利用掃描電子顯微鏡(SEM)對不同濃度ε-PL處理后的細胞表面進行觀察后發現,當用500μg/mL的ε-PL處理釀酒酵母后,其細胞表面會出現孔洞、微膠粒或發生磷脂雙分子層的彎曲等現象。
另外,ε-PL對釀酒酵母的抑制作用還可能與細胞內活性氧(ROS)水平上調引起的細胞凋亡相關。Tan等[33]使用二氫睪酮羅丹明123(DHR 123)測定了經ε-PL處理后的釀酒酵母細胞內的ROS水平,并利用4,6-聯脒-2-苯基吲哚(DAPI)對細胞核DNA分子進行染色,以考察核DNA的變化。結果表明,ε-PL處理細胞12 h后,細胞內的ROS水平顯著上調,同時觀察到DNA的片段化,細胞發生死亡。推測該現象與細胞凋亡現象類似,但仍有待進一步驗證。Liu等[34]對ε-PL和乳酸鏈球菌素(Nisin)組合使用后的抑菌效果及協同抗菌機制進行了探究,發現Nisin能迅速破壞細胞壁和細胞膜,有利于ε-PL隨即進入細胞,與核DNA相互作用以阻止DNA復制。這項研究為ε-PL抑菌機制中有關ε-PL引起ROS水平上調、作用于核DNA進而誘導細胞凋亡的闡明奠定了基礎。Ye等[45]研究也發現,ε-PL作為一種多肽類抑菌劑,會在大腸桿菌中引起ROS脅迫,并且DNA損傷應答(SOS應答)調節基因的表達會受到ε-PL的影響。
近年來,代謝組學技術的蓬勃發展也推動了ε-PL抑菌機制的闡明。代謝組學旨在系統地分析生物中整個小分子代謝物的組成和動態響應,進而揭示生命的表型變化。Bo等[35]通過GC-MS等技術分析檢測了經ε-PL處理后的釀酒酵母細胞內代謝物的變化情況。結果表明,ε-PL處理釀酒酵母后,能引起細胞內多種代謝物(主要包括碳水化合物、醇類、脂類和氨基酸)的代謝水平發生變化。ε-PL會引起細胞內葡萄糖的積累,這表明其可能抑制了糖酵解過程。同時,它會引起糖酵解途徑的中間代謝物(如3-P-甘油酸、丙酮酸等)的濃度降低,這意味著ε-PL可能會對糖酵解途徑產生抑制。另外,它還能使三羧酸循環中間體(如谷氨酸、尿素、鳥氨酸等)濃度增加,從而使琥珀酸鹽濃度降低,這表明α-酮戊二酸主要轉化為谷氨酸,進而可能導致ε-PL對三羧酸循環的抑制。以上研究表明,ε-PL可能通過破壞膜功能,間接地抑制中心碳代謝,從而使菌種對數期延長,抑制菌種的生長,或作用于與代謝密切相關的細胞膜,使磷脂層瓦解,從而使菌種死亡。
5.2.2 ε-PL對大腸桿菌的作用機制
ε-PL對革蘭氏陰性細菌的抑菌機理研究,主要以大腸桿菌(Escherichia coli)作為對象進行探究。ε-PL對大腸桿菌的抑菌作用強弱同樣依賴于ε-PL所添加的濃度。付萍[36]的研究表明,50μg/mL以下的ε-PL處理大腸桿菌后可使其對數期延長,50~150μg/mL的ε-PL處理大腸桿菌后可使其生長受到明顯抑制,而150μg/mL以上的ε-PL處理大腸桿菌后可使其菌液近乎澄清。這同樣表明ε-PL發揮抑菌作用需要一定的閾值濃度。由于大腸桿菌作為一種革蘭氏陰性菌,具有內外膜結構,所以,研究者考察了不同濃度的ε-PL對大腸桿菌內外膜的作用[36~38]。結果發現,高濃度ε-PL對細胞外膜的破壞作用較強,作用40 min后,由疏水探針NPN(NPhenyl-1-naphthylamine)標記的細胞外膜的熒光強度變化趨于平緩。推測帶正電荷的ε-PL可能與大腸桿菌表面呈負電荷的脂多糖通過靜電作用結合,從而使大腸桿菌外膜通透性增加。同時,ε-PL處理菌體后經不同時間段檢測,發現細胞內膜的β-半乳糖苷酶含量均無明顯變化,而用不同濃度的ε-PL處理大腸桿菌后,其內膜的β-半乳糖苷酶含量會發生明顯變化。這表明ε-PL對大腸桿菌的作用是瞬時的,并會造成菌體內膜的通透性發生改變。利用掃描電子顯微鏡(SEM)對ε-PL處理后的大腸桿菌表面的形態變化進行觀察,發現經不同濃度的ε-PL處理后,菌體細胞表面會呈現出明顯的褶皺、凸起和凹陷,同時菌體間會發生聚集粘連。這一現象與抗菌肽抑菌機理中的氈毯模型極為相似。
Hyldgaard等也對ε-PL作用于大腸桿菌的抑菌機制進行了研究[38]。他們測定了大腸桿菌細胞膜表面的Zeta電位,并同樣研究了二價陽離子對抗菌活性的影響,同時通過原子力顯微鏡(AFM)觀測了ε-PL對細胞形態的影響。發現在對細胞膜進行熒光標記后,有熒光小囊泡的形成,這進一步驗證了ε-PL的抑菌機理可能與抗菌肽抑菌機理中的氈毯模型類似。同時,Shima[39]和Vaara[40]發現,陽離子多肽ε-PL通過離子吸附與帶負電的細胞表面相互作用,可干擾并破壞LPS層,導致外膜透化。Hyldgaard等[38]進一步考察了ε-PL對更高耐受性的大腸桿菌ΔrscC突變體的影響,發現莢膜外多糖莢膜異多糖酸的過量表達,有助于調節菌體生物膜的三維結構,并起到一定的屏障作用。去除脂多糖后,發現ε-PL作用于膜磷脂。
5.2.3 ε-PL對枯草芽孢桿菌及李斯特菌的作用機制
ε-PL對革蘭氏陽性細菌的抑菌機理研究,主要以枯草芽孢桿菌(Bacillus subtilis)及李斯特菌(Listeria monocytogenes)作為對象進行探究。Liu等[34]考察了ε-PL和Nisin對枯草芽孢桿菌細胞壁的影響,發現ε-PL單獨作用時僅使細胞輕微受損,Nisin單獨處理枯草芽孢桿菌后,細胞表面粗糙,細胞壁明顯受損。而當兩種物質復合作用時,可使細胞表層結構明顯受損。同時細菌近似空心狀態,細胞內含物逸出。由于Nisin主要被認為作用于細胞表面,故推測ε-PL可能作用于微生物內部結構[41]。
Hyldgaard等[38]初步探究了ε-PL對李斯特菌的抑菌機理。發現ε-PL對菌體形態的影響較小,推測可能是肽聚糖對細胞結構的維持作用所引起。李斯特菌的細胞膜含有賴氨酸衍生的磷脂酰甘油和賴氨酰心磷脂[42~44]。賴氨酸心磷脂是兩性離子[43],沒有凈負電荷,而磷脂酰甘油能進一步減少細胞膜的負電荷,從而減少細胞膜對陽離子肽即ε-PL的親和力[42]。因此ε-PL對李斯特菌具有較高的MIC,進而經ε-PL處理后,菌體細胞膜的通透性較低。綜上所述,關于ε-PL對革蘭氏陽性細菌的抑菌機理研究報道較少,其作用機制有待進一步揭示。
5.2.4 ε-PL對不同菌種的抑菌作用機制比較
從上述ε-PL對不同菌種的作用機制來看,ε-PL對菌種產生抑制作用需要一定的閾值濃度,并可能因細胞壁的結構不同而存在差異。在真菌釀酒酵母的研究中,500μg/mL的ε-PL可使菌種致死。而在革蘭氏陰性細菌大腸桿菌的研究中,150μg/mL的ε-PL即能使菌體聚合沉降,菌液變澄清。有研究推測這可能是由于革蘭氏陰性菌外膜主要由脂多糖和磷脂構成,其肽聚糖層相對較薄且稀疏,故其機械強度偏低。酵母菌其細胞壁最外層為甘露聚糖蛋白,內層葡聚糖起主要細胞機械強度的作用,因其交聯程度低,故機械強度相對較低,但高于革蘭氏陰性菌。而革蘭氏陽性菌細胞壁富含交聯的肽聚糖層,具有較高的機械強度。所以,在前述的革蘭氏陽性菌經ε-PL處理后,其細胞表面變化可能相對穩定[36]。
另外,從前述研究中還發現,ε-PL可能影響菌體細胞膜的通透性和疏水性。在釀酒酵母的研究中,經碘化丙啶染色和熒光強度檢測,發現500μg/mL的ε-PL對菌體細胞膜產生了損傷,增加了細胞膜的通透性。而在大腸桿菌的研究中,ε-PL影響了細胞內外膜的通透性。有研究推測,酵母菌作為一種真菌,其細胞壁最外層為甘露聚糖蛋白,帶有負電荷。同樣,革蘭氏陰性細菌的細胞壁富含脂多糖,會使菌體細胞表面帶有大量的負電荷。因此,帶較多負電荷的酵母菌和革蘭氏陰性菌的細胞表面與帶正電荷的ε-PL作用面積會較大。而革蘭氏陽性細菌細胞壁結構主要由肽聚糖組成,細胞表面所帶的負電荷通常較為稀疏,因此ε-PL可能只會對革蘭氏陽性細菌的細胞壁造成局部破壞[36]。
5.2.5 ε-PL可能的抑菌機制模型
(1)ε-PL抑菌機制與氈毯模型。Hyldgaard等[38]研究了ε-PL的抑菌機制,從多方面驗證并提出了ε-PL可能作用于膜磷脂進而產生抑制作用的機制,并認為ε-PL與大腸桿菌相互作用的模型很可能為氈毯模型(圖2)。氈毯模型是抗菌肽抑菌機制中的主要模型之一。在氈毯模型中,抗菌肽通過靜電吸附作用,與細胞膜上富含陰離子的磷脂分子頭部相結合,類似地毯狀地覆蓋于細胞膜磷脂雙分子層的表面。當抗菌肽的濃度達到閾值后,會在細胞膜上形成瞬時孔洞,使更多未結合的抗菌肽進入細胞內部。此時,磷脂雙分子層會發生彎曲及損傷,隨之細胞膜逐漸裂解并與抗菌肽結合形成微膠粒。而在前述的ε-PL對釀酒酵母的抑菌機制研究中,500 μg/mL的ε-PL作用于釀酒酵母時對其是完全抑制的,顯微觀察發現釀酒酵母經一定濃度的ε-PL處理后表面有明顯的凹陷,并會形成微膠粒(圖2)。這些現象均從一定程度上印證了ε-PL抑菌機制與氈毯模型的相似性。

圖2 氈毯模型
(2)ε-PL抑菌機制與ROS誘導細胞凋亡模型。據報道,ε-PL抑菌機制可能與ROS誘導的細胞凋亡有關。Ye等[45]研究發現,ε-PL會在大腸桿菌中引起ROS脅迫,且DNA損傷應答(SOS應答)調節基因的表達會受到ε-PL的影響。近年來,有較多研究報道了抗菌肽通過ROS誘導細胞凋亡的模型,如有關PAF26、coprisin、melittin等的研究[46~48]。在前述的Tan等[33]報道的關于ε-PL對釀酒酵母細胞內ROS積累和DNA片段化的研究中,也進一步闡述了該機制的可能性(圖3)。當ε-PL處理細胞12 h后,細胞內的ROS水平顯著上調,同時觀察到DNA的片段化,從而引起細胞死亡。同時,在與其他抑菌劑組合使用后的抑菌研究中,還發現ε-PL可與核DNA相互作用的現象。因此,ε-PL可能也會通過調控ROS水平間接影響DNA,從而使細胞凋亡。

圖3 ROS誘導細胞凋亡模型
6 ε-PL研究展望
ε-PL作為一種抗菌譜廣、安全無毒的天然食品防腐劑,具有廣闊的商業化前景。但目前ε-PL的生物合成機理和抑菌機制尚未揭示清楚,阻礙了其商業化應用。因此,研究ε-PL的微觀合成機理及抑菌機理具有重要的科學意義。今后在探究上述機制的基礎上,可從菌種種質資源改良、多組學分析、代謝調控及發酵優化等方面對ε-PL產生菌進行研究,以有效提高ε-PL的產量,并加強ε-PL的應用研究,最終實現ε-PL的產業化應用。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
四川勞動保障(2021年9期)2022-01-18 05:11:08
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年21期)2018-11-09 01:23:06
汽車工程學報(2017年2期)2017-07-05 08:13:02
中國衛生(2016年9期)2016-11-12 13:28:08
中國衛生(2015年9期)2015-11-10 03:11:12
