底泥中三氯生殘留對輪葉黑藻的生態毒性效應
2019-10-08 07:16:54張立娜王芷茵
農業環境科學學報 2019年9期
張立娜,王芷茵,安 婧
(1.中國科學院沈陽應用生態研究所污染生態與環境工程重點實驗室,沈陽 110016;2.中國科學院大學,北京 100049;3.東北大學資源與土木工程學院,沈陽 110016)
三氯生(2,4,4'-三氯-2'-羥基二苯醚,triclosan,TCS)是一種使用廣泛的廣譜殺菌劑。由于高效的殺菌性、不刺激皮膚、不會引起過敏及與日用化工原料有良好的配伍等特性,被廣泛用于牙膏、香皂、除臭劑、漱口水等家庭日常用品及個人衛生用品中[1]。工業的發展及個人護理用品的廣泛使用,使TCS的消耗量逐年上升,近30年來排放到環境中的TCS也不斷增加。據統計,全球TCS年消耗量在7.5×105t以上[2]。隨著人們對TCS等環境新型污染物的日益關注以及化學分析檢測技術的飛速發展,TCS在世界多地的環境樣品中被檢出。近年來,我國多地的水體樣品中也檢出了TCS殘留,且檢出率和檢出濃度顯著高于其他國家。Sun等[3]在中國九龍江水中檢測到TCS的殘留濃度為 96.5 ng·L-1;Oliveira等[4]研究表明日本地表水中TCS殘留濃度約為11 ng·L-1。由于TCS具有較好的穩定性和低揮發性,其辛醇比為4.8,因此,TCS在水體沉積物中吸附系數也較大,導致進入水體中的TCS可以進入到水體沉積物中并持續存在。Zhang等[5]發現,長江水體中TCS含量高達1.89 ng·L-1,底泥中含量達1.2 ng·g-1;Zhao等[6]發現,遼河、海河、黃河、珠江和東江等水體中TCS的質量濃度分別為1.39~2.6、0.156~39.1、0.43~3.72、1.89~5.2、11.7 ng·L-1,而底泥中質量濃度分別為 0.41~0.6、3、0.05~2.05、1.2~5.78、0.002~14.8 ng·g-1。而對于污染段來說,水體中TCS的質量濃度高達478 ng·L-1,底泥中質量濃度高達2723 ng·g-1。由于TCS具有類持久性、生物富集性、熱穩定和難揮發性等特點,其類持久性主要表現在能持久存在于環境中,具有很長的半衰期,導致其可以在水體和底泥中持久存在。而底泥中TCS殘留很容易被人類所忽視,其對水體動植物具有不容忽視的潛在生態風險。
底泥與水環境之間存在動態平衡,底泥在吸附的同時也會向水環境釋放TCS,從而對水生動植物產生生態毒性效應。韓琦等[7]研究表明河流對來自陸上的有機污染物主要通過固相吸附及沉降作用而在底泥中富集,并且底泥污染物會在一定水力條件和底棲生物的擾動作用下,通過水-底泥的交換反應在液相和固相間遷移。在TCS水體暴露實驗中,董玉瑛等[8]發現東北林蛙幼體蝌蚪暴露于TCS 1 h,15℃時LD50(半數致死量)為712.32μg·L-1;Hwang等[9]發現TCS對海膽胚胎的影響也很顯著,可以導致胚胎發育異常,使囊胚細胞分化緩慢。水生植物的研究主要集中在TCS對微藻類的急性毒性效應。陳東等[10]研究發現在毫克每升水平上TCS對淡水綠藻有明顯的毒性,可顯著抑制其生長、降低葉綠素含量以及抗氧化酶活性;李義剛等[11]研究發現TCS對羊角月牙藻的半數效應濃度(96 h-EC50)為0.112 mg·L-1。然而,目前水環境中TCS對大型藻類的毒性效應,尤其是TCS低濃度長期暴露的毒性效應研究相對匱乏。此外,針對殘留于底泥中的TCS對水生生物的生態毒性效應研究并不多見。
輪葉黑藻(Hydrilla verticillata)作為一種典型的沉水植物,在我國水環境中分布廣泛。輪葉黑藻能凈化水質并且吸收富集污染物質,且其整株浸于水中,根部插入底泥中,因而對水體污染物脅迫的響應較為敏感。李國新等[12]研究發現,輪葉黑藻葉片吸附Pb后,細胞內外離子的快速交換過程使細胞干癟,發生明顯的變形,細胞內膜系統的完整性遭到破壞;韓曉弟等[13]研究發現,在Pb2+脅迫下,輪葉黑藻葉片呈現不同程度的枯黃萎蔫現象,其程度隨處理濃度的增加而加重。目前關于TCS脅迫輪葉黑藻產生的毒性效應研究很少。基于此,本實驗以輪葉黑藻作為受試植物,研究了底泥環境低濃度TCS長期暴露下,輪葉黑藻對TCS的吸收以及其體內葉綠素、可溶性蛋白以及抗氧化酶系統對TCS脅迫的響應,以期為新型污染物的生態風險評估提供毒理學基礎數據和理論依據。
1 材料與方法
1.1 實驗儀器和供試材料
儀器:冷凍高速離心機(Centrifuge 5804 R),純水儀(DW 100);超聲波清洗器(KQ-500 DE),氮吹儀(DCY-24 Y),固相萃取裝置(SPE-12 A),烘箱(101-2 ABS),分析天平(JA 2003),紫外可見分光光度計(VARIAN Cary 50),液相色譜-質譜聯用儀(Maters 2695.ZQ 4000)。
供試材料:供試植物為輪葉黑藻,屬水鱉科,單子葉多年生沉水植物。輪葉黑藻芽孢購自遼寧省安新縣冀中水生植物種植公司,于實驗室培養兩周,選取10 cm左右長度的輪葉黑藻進行實驗。實驗用水為去離子水。營養液配制按照Hoagland′s培養液,輪葉黑藻采用10%Hoagland′s培養液。河流底泥采自遼寧省撫順市大伙房水庫(41.842°N,124.154°E),大伙房水庫為飲用水源地保護區,底泥中未檢測到TCS,含水率為33%。TCS購自阿拉丁生化科技股份有限公司(上海),純度≥96%,所用其他試劑均為分析純。將TCS溶于二甲基亞砜(DMSO)(DMSO=1 mL),配制母液(5 g·L-1)。
1.2 實驗設計
采用水培實驗,模擬河流環境,在無色玻璃水缸(長35 cm×寬20 cm×高25 cm)底部鋪設一層4 cm厚的底泥,每缸中放8 L用去離子水配制的10%Hoagland′s培養液。水缸上面放置常規架燈提供光照,同時進行曝氧,實驗裝置如圖1所示。選用株高10 cm左右,鮮質量約0.8 g的輪葉黑藻進行實驗。每個缸中定植20株,平均栽植在底層底泥上,栽植深度為1.5 cm左右。

圖1 實驗裝置簡圖Figure 1 Experimental device diagram
將所取底泥進行自然風干,并且去除里面的石頭、草枝等雜物。將TCS溶液均勻地拌入底泥樣品中,濃度分別為0.05、0.1 mg·kg-1和0.5 mg·kg-1,同時設置無TCS添加的空白對照組,平衡24 h,實驗設置三個平行。
將各實驗組放置于25℃恒溫植物培養室中,光暗時間比為12 h∶12 h,照度約為3600 lx,實驗進行28 d。分別在培養的第7、14、21 d和28 d采集植物樣品按照相關分析測試方法進行前處理后保存待測。
1.3 分析方法
1.3.1 TCS在葉片中的殘留
將采集的植物葉片樣品冷凍干燥24 h,稱取0.5 g,剪碎研磨,用10 mL乙腈提取,超聲30 min,在5000 r·min-1下離心10 min,取上清液轉移到40 mL棕色玻璃瓶中,重復以上步驟兩次。用10%H2SO4調節pH至2~3左右后,利用固相萃取法進行濃縮富集。最后用氮吹儀將濃縮液吹干,用1 mL乙腈定容并過0.45μm濾膜后,置于棕色小玻璃瓶中,保存在-20℃冰箱中待測。用液相色譜-質譜聯用儀測定樣品中TCS含量[14-17],測試條件為:流動相為乙腈和水,采用梯度洗脫方式:0~8 min,乙腈由20%勻速增加到85%;保持1 min;9~10 min乙腈由85%降回到20%。流速為1.0 mL·min-1;進樣體積為20μL;柱溫為30 ℃;在負離子模式下進行掃描;紫外檢測波長為282 nm。本實驗所取標樣濃度依次為 0.002、0.005、0.01、0.05、0.1、0.2、0.4μg·mL-1。標準曲線為y=-1 889.85+3.480 46x,R2=0.999 1。本實驗方法的回收率在75%~85%。
1.3.2 生理生化指標的測定
葉綠素采用分光光度法[18],新鮮樣品取0.2 g,加入少量石英砂和碳酸鈣粉及2~3 mL提取劑95%乙醇并剪碎研磨至勻漿,過濾定容。以665、649 nm下的吸光值計算葉綠素含量,單位為mg·g-1。可溶性蛋白質量分數采用考馬斯亮藍G-250染色法[19],配制蛋白質標準溶液和考馬斯亮藍G-250溶液,稱取0.2 g新鮮樣品研磨勻漿,蒸餾水定容至10 mL,離心上機。以540 nm下的吸光值計算可溶性蛋白含量,以鮮質量計,單位為mg·g-1。測定抗氧化酶之前處理樣品,稱取0.5 g樣品,用預冷后的蒸餾水研磨至勻漿,轉入15 mL離心管,蒸餾水反復沖洗研缽,合并清洗液并定容,于4 ℃下12 000 r·min-1離心10 min。POD采用愈創木酚法[18,20],以470 nm下的吸光值計算酶活性,以鮮重計,單位為U·g-1·min-1;CAT采用紫外吸收法,以240 nm下的吸光值計算酶活性[18,20],以鮮質量計,單位為U·g-1·min-1;SOD采用氮藍四唑法,以560 nm下的吸光值計算酶活性[18,20],以鮮質量計,單位為U·g-1。
1.4 數據分析
采用Microsoft Excel 2013軟件對數據進行處理,SPSS 17.0統計分析軟件對數據進行差異顯著性檢驗(LSD法,α=0.05),用英文小寫字母表示各處理組間的差異顯著性(P<0.05),用Origin 8.0進行繪圖。
2 結果與分析
2.1 輪葉黑藻葉片對底泥中TCS的吸收
TCS底泥暴露28 d內,輪葉黑藻葉片中TCS的殘留情況如表1所示。由表可見,TCS暴露初期(7 d),各處理組中輪葉黑藻葉片中TCS濃度大小依次為:TCS0.1>TCS0.5>TCS0.05。暴露 14 d 時,輪葉黑藻葉片中TCS含量與暴露7 d時相比略有下降,這可能與TCS在進入植物體內初期的遷移和降解速率有關。隨TCS暴露時間的延長,植株葉片中TCS含量逐漸升高,這可能與植株進入其生長旺盛期有關。0.5 mg·kg-1TCS暴露28 d時,輪葉黑藻葉片中TCS濃度最高,達到2.16 mg·kg-1。暴露濃度對輪葉黑藻吸收TCS的能力具有顯著影響,由表1可見,0.05、0.1 mg·kg-1TCS處理組中輪葉黑藻對TCS吸收速率在暴露21 d后逐漸減緩,而0.5 mg·kg-1TCS暴露28 d時輪葉黑藻對TCS的吸收速率仍可達62%。在整個暴露周期中,不同濃度TCS處理組中植物葉片TCS含量均呈先下降再回升的趨勢。但從整體看,與暴露初期植株體內TCS含量相比,TCS含量變化呈低濃度組趨于穩定、中濃度組降低、高濃度升高的變化規律。
2.2 TCS對輪葉黑藻葉綠素含量的影響
TCS底泥暴露對輪葉黑藻葉綠素含量的影響情況如圖2所示。由圖2A中可知,在28 d的暴露周期內,各處理組輪葉黑藻葉片中葉綠素含量與對照組相比顯著降低,且隨暴露時間延長,葉綠素含量受抑制的程度逐漸增加。0.5 mg·kg-1TCS暴露28 d時,植株葉片葉綠素含量的抑制率高達45.7%。
與葉片中葉綠素的變化情況相比,TCS對輪葉黑藻莖部葉綠素含量的抑制作用較弱。由圖2B中可知,暴露7 d和14 d時,隨著TCS暴露濃度的增加,植株莖部葉綠素含量逐漸降低。TCS暴露21 d后,莖部葉綠素含量有所回升,0.05~0.1 mg·kg-1TCS處理組中莖部葉綠素含量基本恢復至對照水平,但組間差異不顯著,而0.5 mg·kg-1TCS處理組中輪葉黑藻莖部葉綠素含量仍然受到顯著抑制。TCS暴露28 d后,莖部葉綠素含量抑制程度增加,組間差異顯著。
2.3 TCS對輪葉黑藻可溶性蛋白的影響
TCS對輪葉黑藻可溶性蛋白含量的影響情況如圖3所示。由圖3A中可知,暴露初期,TCS對輪葉黑藻葉片中可溶性蛋白含量具有促進作用,其中0.05 mg·kg-1TCS處理組輪葉黑藻葉片中可溶性蛋白含量升高最為顯著。隨暴露時間延長,植株葉片中可溶性蛋白含量逐漸降低。暴露14 d時,0.05~0.1 mg·kg-1TCS處理組葉片中可溶性蛋白含量雖有所降低,但仍呈激活效應,而0.5 mg·kg-1TCS處理組葉片中可溶性蛋白含量受到顯著抑制。暴露21 d后,各處理組輪葉黑藻葉片中可溶性蛋白含量均受到顯著抑制,且處理組間差異顯著。
與葉片中可溶性蛋白含量的變化情況相比,TCS對輪葉黑藻莖部可溶性蛋白含量的影響則呈現出明顯的抑制效應(圖3B)。TCS暴露7 d后,中高濃度處理組(0.1~0.5 mg·kg-1)輪葉黑藻莖部可溶性蛋白含量顯著低于對照組和低濃度處理組(0.05 mg·kg-1)。隨暴露時間延長,植株莖部可溶性蛋白含量有所回升,但仍呈抑制狀態。暴露21 d后,0.05 mg·kg-1和0.1 mg·kg-1TCS處理組組間差異不顯著,但0.5 mg·kg-1TCS處理組仍顯著低于對照組。

表1 輪葉黑藻葉片中TCS濃度(mg·kg-1)Table 1 TCS concentration in leaves of Hydrilla verticillate(mg·kg-1)

圖2 TCS對輪葉黑藻葉片(A)和莖部(B)葉綠素含量的影響Figure 2 Effects of TCS on chlorophyll content of Hydrilla verticillata(A:blade;B:stem)
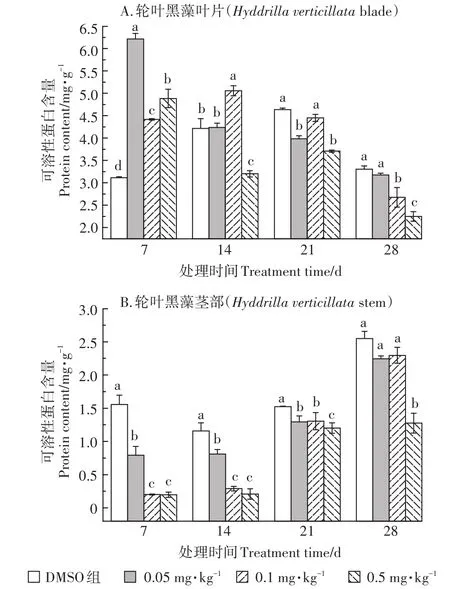
圖3 TCS對輪葉黑藻葉片(A)和莖部(B)可溶性蛋白的影響Figure 3 Effects of TCS on soluble protein of Hydrilla verticillata(A:blade;B:stem)
2.4 TCS暴露對輪葉黑藻酶系統的影響
TCS暴露對輪葉黑藻葉片及莖部抗氧化酶活性的影響情況如圖4和圖5所示。由圖4可知,暴露初期,三個處理組中葉片中CAT活性均呈激活效應。隨TCS暴露濃度增加,CAT活性逐漸降低,組間差異顯著。TCS暴露28 d時,各處理組中葉片CAT活性呈抑制狀態。與CAT相似,0.05~0.5 mg·kg-1TCS暴露7~14 d時,植株葉片中SOD活性呈現激活狀態,且隨暴露濃度和時間的增加,激活效應增強,組間差異顯著。0.05~0.1 mg·kg-1TCS暴露21 d時,處理組葉片中SOD活性呈激活狀態,且組間差異顯著,而0.5 mg·kg-1TCS處理組中SOD活性呈抑制狀態。暴露末期(28 d),三個處理組葉片中SOD活性有所回升,呈輕微激活狀態。與CAT和SOD活性變化趨勢不同,葉片中POD活性受TCS脅迫則在整個實驗周期內均呈現抑制狀態。暴露21 d時,三個處理組中葉片POD活性顯著降低,抑制率分別為84.9%、79.6%、87.1%。暴露28 d時,三個處理組中葉片POD活性有所回升,但仍處于抑制狀態。
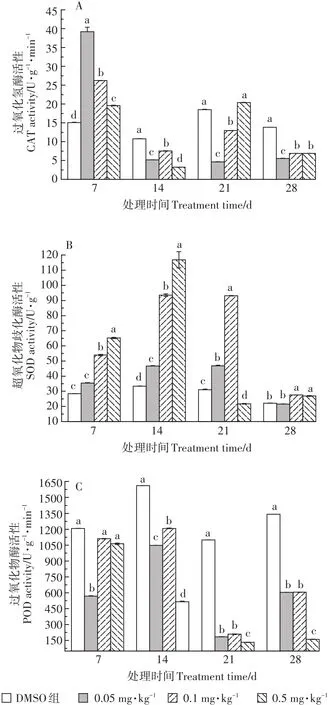
圖4 TCS對輪葉黑藻葉片酶系統的影響Figure 4 Effects of TCS on antioxidant enzyme system of Hydrilla verticillate blade
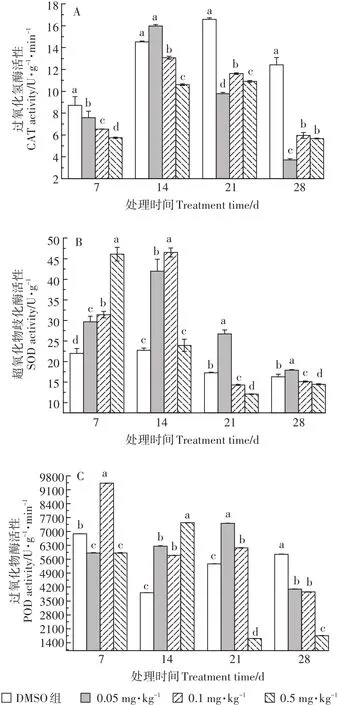
圖5 TCS對輪葉黑藻莖部酶系統的影響Figure 5 Effects of TCS on antioxidant enzyme system of Hydrilla verticillata stem
TCS脅迫下,植株莖部抗氧化酶活性的變化情況與葉片中有所不同。由圖5可知,暴露初期,三個處理組中莖部CAT活性均呈抑制狀態,且隨暴露濃度的增加,抑制程度增加,組間差異顯著。暴露14 d時,莖部CAT活性呈現低濃度(0.05 mg·kg-1TCS)被促進,而中高濃度(0.1~0.5 mg·kg-1TCS)被抑制的趨勢。隨著暴露時間延長,CAT活性逐漸降低。TCS暴露28 d時,三個處理組莖部中CAT活性達到了最低值,其抑制率分別為72%、55.8%、59.8%。與CAT不同,暴露7~14 d時,三個處理組中莖部SOD活性均呈激活狀態,且組間差異顯著。隨暴露時間延長,0.05 mg·kg-1TCS處理組中莖部SOD活性呈激活狀態,而0.1~0.5 mg·kg-1TCS處理組中莖部SOD活性被顯著抑制,且TCS濃度與莖部SOD活性抑制率之間具有一定的劑量-效應關系。POD與CAT和SOD活性變化趨勢不同,7~14 d時,隨暴露時間延長,誘導作用增加。TCS暴露21 d后,POD活性降低,而0.05~0.1 mg·kg-1TCS處理組中莖部POD活性仍呈誘導效應,但0.5 mg·kg-1TCS處理組中莖部POD被抑制。暴露末期,三個處理組中莖部POD活性均呈抑制狀態。
3 討論
TCS作為一種較為流行的廣譜抗菌劑被廣泛應用于各類洗護產品中,它會隨人類活動如洗漱、淋浴、衛生消毒等進入水體與底泥,最終對生態系統以及人類健康產生一定的危害[21]。本實驗結果顯示,長期暴露于TCS環境下的沉水植物——輪葉黑藻體內有TCS的殘留,而且植物體內TCS的含量呈現先降低后回升的趨勢。在整個暴露周期內,與暴露初期植株體內TCS含量相比,TCS含量變化呈低濃度組趨于穩定、中濃度組降低、高濃度升高的變化規律。張蕾[22]研究沉水植物對有機氯農藥的富集過程中也觀察到有機氯污染物在植物體內先降低后升高的變化趨勢。這可能是由于植株的富脂性表皮有利于吸附親脂性的有機物的緣故。在暴露初期,輪葉黑藻葉片中累積TCS的量較多,可能是由于輪葉黑藻初期接觸TCS毒性響應不明顯,而葉片表面能大量吸附TCS的緣故。有研究表明表面活性劑可將土壤/底泥相有機污染物解吸出來轉移到水相中[23]。但DMSO是否促進底泥中TCS的釋放仍需進一步研究。隨著暴露時間的延長,葉片中TCS含量降低,這與輪葉黑藻對進入其體內的TCS具有一定的遷移和降解性有一定的關系。張蕾[22]研究發現,有機氯農藥在輪葉黑藻體內存在吸附-轉運-降解的過程。隨后,可能是由于輪葉黑藻葉片對水體中TCS的吸收作用大于其在葉片中的降解過程,使TCS在葉片中含量又出現升高現象。本實驗中,TCS暴露28 d后,0.5 mg·kg-1TCS處理組中輪葉黑藻葉片中TCS濃度和累積速率最大。暴露前期,0.5 mg·kg-1的TCS顯著抑制了植株的生長,致使0.5 mg·kg-1TCS處理組中葉片對TCS的累積效應低于低濃度組。暴露后期,輪葉黑藻適應TCS環境脅迫,低濃度處理組中葉片TCS濃度已接近飽和狀態,而高濃度處理組中葉片后期持續累積TCS。這與Ahmed等[24]的研究結果相似,暴露一定時間后,隨四環素和磺胺類抗生素暴露濃度增加在生菜、西紅柿以及黃瓜體內累積量也隨之增加。
葉綠素是植物進行光合作用的主要色素,其含量的高低反映植物的生長能力和代謝能力。在本實驗中,TCS暴露7~28 d時,各處理組中葉片葉綠素含量顯著降低,與葉片中葉綠素含量影響情況類似,7~14 d時,輪葉黑藻各處理組莖部中葉綠素含量與對照組相比呈現抑制效應。這可能是TCS暴露干擾了植株光合作用等關鍵代謝過程,從而影響它的生長[25]。TCS暴露也可能破壞細胞結構,使葉綠素的含量降低,也可能是TCS破壞了細胞膜系統脂質過氧化而導致葉綠素細胞器膜的受損,從而影響了葉綠素的合成[26]。雖然輪葉黑藻葉片中殘留濃度在7~14 d呈下降趨勢,但在這個時期植株生長仍然受到脅迫,且TCS在輪葉黑藻葉片中或者底泥微生物作用下可能降解代謝為更高毒性的甲基三氯生或者二噁英等代謝產物以致脅迫效應更大。有報道稱TCS在其特定菌作用下可以生成甲基三氯生(MTCS)[27-28]。董玉瑛等[8]研究比較蝌蚪在避光和紫外下TCS的毒性作用,測得避光下 LD50值為 564.61 μg·L-1,紫外線照射下LD50值為 486.01 μg·L-1,后者毒性有較明顯增加,這說明代謝產物毒性可能大于TCS母體本身。此外,實驗發現TCS暴露7~14 d時,莖部葉綠素含量的降幅與TCS暴露濃度呈明顯的劑量-效應關系。這與陳東等[10]研究結果類似,TCS對蛋白核小球藻的葉綠素含量的抑制程度與TCS暴露濃度呈正相關。TCS暴露對莖部葉綠素含量的變化較葉片影響較小,可能是由于輪葉黑藻的莖部沒有形成層,只有初生結構,大部分TCS通過維管束結構傳輸到葉片中去的緣故。
可溶性蛋白作為重要的滲透調節物質和營養物質,能夠提高細胞的保水能力。同時,可溶性蛋白也被認為能反映植株抗逆性的狀態,它的含量的變化意味著植株處于較嚴峻的環境,但是環境嚴峻程度的持續增高,可能使可溶性蛋白含量產生不符合其生理規律的變化。在本實驗中,暴露初期,處理組葉片中可溶性蛋白含量顯著高于對照組,這可能是環境處于逆境時植株的抗逆性反饋所造成的。這與陳東等[10]的研究結果類似,TCS處理組使蛋白核小球藻可溶性蛋白含量增加。隨著暴露時間延長,TCS處理組葉片中可溶性蛋白含量呈現抑制狀態,且高濃度TCS處理組對可溶性蛋白的抑制程度顯著大于低濃度TCS處理組,這可能是由于TCS的脅迫使輪葉黑藻葉片中R基因的激活受到影響,進而影響可溶性蛋白的合成,或者是由于高濃度TCS使合成的可溶性蛋白分解[10]。輪葉黑藻葉片中可溶性蛋白在暴露后期呈顯著抑制狀態,與葉片中TCS殘留濃度在后期持續增加有關。有研究表明,當植物受有機物、重金屬以及寒旱等脅迫時,植物體內的SP含量會下降[29]。與葉片中可溶性蛋白含量的變化情況相比,在整個暴露周期莖部可溶性蛋白始終處于抑制狀態,這可能是由于植株的莖部直接接觸底泥,同時TCS破壞莖部薄壁細胞,莖部的生長發育受到抑制。
在植物正常生長的情況下,生物體內活性氧與抗氧化系統之間具有動態平衡系統,但是植株生長的環境有污染脅迫時,會產生過多的活性氧,破壞了細胞內氧化與抗氧化平衡,損傷生理生化功能,導致一系列損傷。因此,在很多研究中,SOD、CAT和POD被用作污染物毒性的指標[30-32]。在本實驗中,暴露前期(7~14 d),TCS處理組中輪葉黑藻各部位SOD活性均呈激活效應,但后期出現一定的抑制作用。SOD作為植株體內重要的氧自由基消除劑,它被認為是抗氧化防御系統的第一道防線,能夠把O-2·轉化成為 O2和H2O2,抑制羥基自由基等的形成,從而終止自由基連鎖反應,來保護植物的氧化損傷[33]。輪葉黑藻被TCS污染脅迫后,可能導致體內產生大量的O-2·,致使SOD活性升高來維持平衡。同時根據植物殘留來看后期TCS濃度增加,輪葉黑藻在高濃度TCS脅迫下,藻細胞結構遭到破壞,超過藻自身調節范圍,從而導致SOD合成障礙,活性下降[25]。與SOD不同,暴露初期,三個處理組中葉片CAT活性均呈誘導效應,隨暴露時間延長,CAT活性呈抑制狀態。而在整個暴露周期內,輪葉黑藻莖部CAT活性均呈抑制狀態。葉片中POD活性受TCS脅迫在整個暴露周期內均呈抑制狀態,不同濃度TCS處理組中頸部POD活性隨暴露時間延長呈現先增加后降低的趨勢。TCS暴露28 d時,三個處理組中葉片POD活性均呈抑制效應。POD和CAT被認為是抗氧化系統的第二道防線,通過減少H2O2來提高細胞對氧化損傷的抵抗力。它們可以協同作用使SOD產生的H2O2轉化成H2O和O2[34]。由于OD酶活在暴露初期有一定的促進現象,本實驗中,POD和CAT在暴露初期為了維持抗氧化防御系統的平衡酶活性相對較高,但后期兩者的酶活性也逐漸降低,整個抗氧化防御系統被破壞。
4 結論
(1)輪葉黑藻對底泥暴露的TCS具有吸收作用,植株葉片中TCS的殘留濃度受暴露時間和暴露濃度的影響顯著。
(2)TCS底泥暴露可顯著抑制輪葉黑藻葉片中葉綠素含量,莖部中可溶性蛋白含量也被顯著抑制,而輪葉黑藻葉片中可溶性蛋白含量呈先促進后抑制的趨勢。葉綠素和可溶性蛋白均為典型的生理生化指標,說明TCS對輪葉黑藻有一定的生態毒性效應。
(3)TCS底泥暴露對輪葉黑藻體內CAT、SOD和POD活性具有顯著影響,嚴重破壞了植株體內抗氧化防御系統的動態平衡,影響植株的正常生長發育。
