融合黃瓜光質需求的設施光環境智能調控模型
2019-10-10 02:50:16荊昊男李遠方張仲雄張海輝
農業機械學報 2019年9期
關鍵詞:模型
胡 瑾 荊昊男 高 攀 李遠方 張仲雄,3 張海輝,3
(1.西北農林科技大學機械與電子工程學院, 陜西楊凌 712100;2.農業農村部農業物聯網重點實驗室, 陜西楊凌 712100;3.陜西省農業信息感知與智能服務重點實驗室, 陜西楊凌 712100)
0 引言
黃瓜是一種重要的設施農業作物,光環境是影響黃瓜生長發育的重要因素,其不僅與光照強度有關[1-6],也與光質比(紅光占總光強的比例)密切相關。研究證明,合理調節光質比能顯著提升作物品質[7-8]。KHATTAK等[9]研究表明金魚草植物高度、節間長度和葉面積受光質影響較大。MIAO等[10]證明藍光比紅光更有利于促進黃瓜葉片光合電子傳遞。王君等[11]證明采用不同紅藍配比光處理的生菜其光能利用效率存在明顯差異。劉彤等[12]研究發現在不同條件下,植物光合作用所需的紅藍光強度比例也存在不同。但是目前大多數光環境調控的模型和系統研究只涉及到固定的光質比調控[13-15],近期也出現了考慮環境對光質影響的研究,其一定程度上提高了調控效果[16],但由于設置的試驗條件不足,以及算法限制等問題,并沒有進一步進行光質動態連續調控模型和方法的研究,造成現有設施光環境調控模型通常只考慮了固定光質比下的光強精準調控,尚未引入環境動態變化下的融合光質比和光強的協同調控方法,影響光環境調控效果。所以,建立融合作物光質需求的設施光環境智能調控模型,是設施農業環境調控急需解決的問題之一。
本文擬構建融合黃瓜光質需求的設施光環境智能調控模型。首先基于試驗數據,利用回歸型支持向量機(Support vector regression algorithm, SVR)算法建立融合黃瓜光質需求的光合速率預測模型,然后利用粒子群算法尋找特定溫度下最大光合速率點對應的光照強度和光質比,最后基于尋找的數據建立光合速率調控目標值模型。
1 材料與方法
1.1 試驗材料
試驗于2019年4月在西北農林科技大學北校區農業物聯網重點實驗室進行,供試的黃瓜品種為“金胚98-1F1”,試驗期間,對試驗材料在培養箱進行正常的培育,不噴灑任何農藥和激素。本試驗主要設計了不同光質比、光強和溫度耦合的嵌套試驗,選取長勢優良的黃瓜植株60株,采用LI-6800型便攜式光合速率測試儀測定樣本凈光合速率。為了避免因天氣狀況不同和植株“午休”對試驗結果造成影響,選擇較為晴朗的天氣,于08:30—11:30,14:30—17:30期間進行試驗。利用光合速率測試儀自帶的多個子模塊設置不同的環境條件進行試驗,其中,CO2濃度控制在400 μmol/mol,控溫模塊設定18、21、24、27、30、33℃共6個溫度梯度,LED光質比設定為0.1、0.3、0.4、0.5、0.6、0.7、0.9共7個梯度,LED光源設定為0、30、75、150、300、500、650、800、1 000、1 200、1 400、1 600、1 800 μmol/(m2·s)共13個光照強度梯度,獲取以溫度、光質比、光照強度為輸入,黃瓜凈光合速率為輸出的546組樣本數據。
1.2 融合光質需求的光合速率預測模型建立
智能算法可有效提升模型的預測效率[17-19], 相關研究發現BP神經網絡和隨機森林算法的運算效率和預測精度一般不及支持向量機算法[20],因此本文采用回歸型支持向量機進行模型構建。該模型以溫度、光質比、光照強度為輸入,黃瓜凈光合速率為輸出,選用高斯核函數進行數據與特征空間之間的映射,最后在特征空間構建最優回歸平面,完成模型建立,其流程圖如圖1所示。
(1)由于不同樣本數據之間的差距比較大,量綱也有所不同,故先對全體樣本數據進行歸一化處理,將數據統一到同一個區間[0,1]之內,所用的歸一化計算式為
y′=(x-xmin)/(xmax-xmin)
(1)
式中y′——歸一化之后的數據
xmin——同一量綱數據中的最小值
xmax——同一量綱數據中的最大值
(2)從歸一化后的546組數據中,隨機選取437組數據(占樣本總數據的80%)作為訓練集數據構建SVR模型,其余的109組數據(占樣本總數據的20%)作為測試集數據以驗證模型的泛化能力。
(3)本研究樣本數據之間存在明顯的非線性關系。對于非線性支持向量機問題來說,首先需選取合適的核函數,將數據映射到特征空間,然后在特征空間里利用線性支持向量機方法尋找最優回歸平面。本文采用高斯核函數進行數據的映射變換,并采用網格迭代尋優方法進行懲罰因子c和核函數參數g優化。其中,c控制對超出誤差樣本的懲罰程度,g決定了數據映射到新的特征空間后的分布,與函數的寬度參數有關。本文經過迭代尋優得到最優的c為17,g為5。其中高斯核函數的表達式為
(2)
式中δ——函數寬度參數,控制函數的徑向作用范圍
x——訓練集函數的自變量
xi——核函數中心
映射之后在特征空間內利用拉格朗日函數求解回歸平面,所得最優回歸平面為
(3)
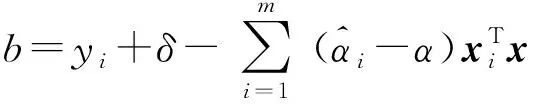
(4)
(4)以測試集實測凈光合速率為實測值,模型計算值為預測值,進行光合速率模型泛化能力和相關性比較。
1.3 基于粒子群算法的最優光照強度、光質比獲取

圖2 粒子群算法尋優流程圖Fig.2 Flowchart of particle swarm optimization algorithm
為了尋找特定溫度條件下的光飽和點和最佳光質比,本文采用粒子群算法進行尋優。粒子群算法是一種基于迭代的優化算法,具有操作方法簡單,尋優速度快的特點[21]。該算法基于群體中個體位置信息和食物位置信息連續發送的機制,利用信息共享完成個體移動方向和移動速度調整,從而實現群體不斷地從雜亂無序的運動變為有序的活動,完成從隨機解出發尋找最優解的過程。基于上述思想,本文采用光合速率預測模型實例化的方法建立不同溫度下的尋優適應度函數,利用粒子群算法獲取不同溫度下凈光合速率最大點,以及對應的光照強度和光質比,其流程圖如圖2所示,尋優具體過程包括以下步驟:
(1)進行特定溫度下的光合速率預測模型實例化,并以此構建尋優目標值函數。
(2)確定模型參數,其中種群規模(即算法所需粒子的數量)為60,算法最大迭代次數為300,粒子的位置范圍為[0,1],速度范圍為[-0.005,0.005],粒子的個體學習因子為0.5,社會學習因子為1.5,慣性權重為0.5。
(3)初始化每個粒子的位置和速度,并以每一點對應的光合速率預測值作為適應度函數進行計算。
(4)通過比較每個粒子的適應度尋找初始化后群體中的最優位置及其適應度值,在此基礎上進行迭代尋優,使每個個體都向最優解的方向運動,直到尋找到最優解。
(5)當滿足尋優結束條件,即運行次數達到該算法設置的最大迭代次數后自動停止,并記錄和保存所尋找到的最優解結果。
1.4 光合速率調控目標值模型建立
粒子群尋優算法尋找了特定溫度下光合速率最優點對應的光質比和光照強度,基于此尋優結果,利用偏最小二乘回歸法構建黃瓜的光合速率調控目標值模型,主要包括以下步驟:
(1)基于粒子群算法尋找到的特定溫度下最大光合速率對應的光質比和光照強度,分別計算對應的紅、藍光的光照強度,計算式為
Pr=PL
(5)
Pb=P(1-L)
(6)
式中Pr——最優點紅光光照強度
Pb——最優點藍光光照強度
L——最優光質比
P——最優總光照強度
以溫度為輸入,對應的紅光和藍光的光強為輸出,采用最高階次為三次的偏最小二乘回歸法,進行模型擬合,得到的調控模型表達式為
y=at3+bt2+ct+d
(7)
式中a、b、c、d——調控曲線系數
t——環境溫度
y——紅光或藍光需求量
2 試驗結果與分析
2.1 試驗結果
分析了溫度、光照強度、光質比與凈光合速率的關系,得到部分試驗曲線如圖3所示。由圖3可知,在相同溫度下,隨著光照強度的增大,黃瓜凈光合速率總體趨勢增大,當光照強度較低時,黃瓜的凈光合速率隨光照強度增長而快速上升,到達一定值后增長速度變緩,甚至會出現下降。而且光質比對黃瓜凈光合速率也有顯著影響,在不同溫度下其最佳光質比不一致,整體來說,隨著光質比的增加,光合速率呈先增后減的趨勢。因此,建立光質比動態調控可以在一定程度上優化調控效果。
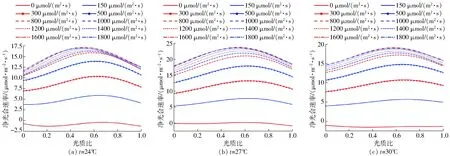
圖3 不同溫度下黃瓜凈光合速率與光照強度、光質比的關系Fig.3 Relationships between net photosynthetic rate,light intensity and light quality of cucumbers
2.2 光合速率預測模型驗證
為了驗證SVR光合速率預測模型的性能,同時選取BP神經網絡算法、隨機森林算法(Random forest, RF)建模進行對比,3種模型的評價指標對比如表1所示。
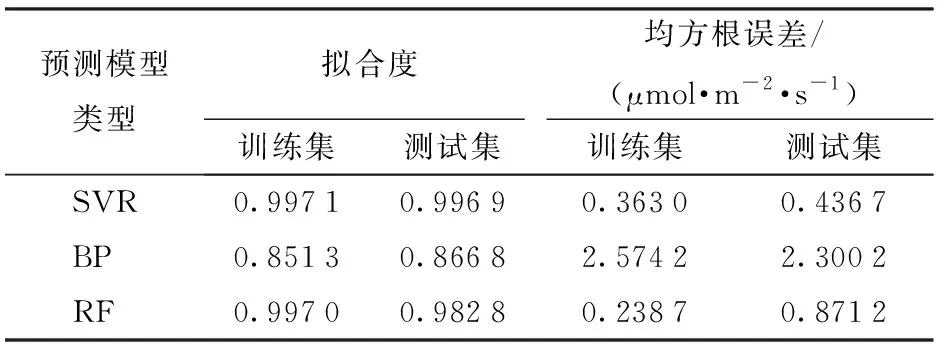
表1 3種模型的擬合度和均方根誤差對比Tab.1 Comparisons of fitness and root mean square errors of three models
由表1可知,利用SVR算法建立的模型訓練集數據和測試集數據的擬合度分別為0.997 1與0.996 9,均高于BP神經網絡算法和RF算法建立的模型;進一步對比均方根誤差可知,隨機森林算法在模型建立過程中出現過擬合,雖然訓練集的均方根誤差最小,但測試集的均方根誤差較大。綜合考慮,選用SVR算法能更好地反映數據之間的關系。
為了觀察SVR預測模型的泛化能力,選用測試集的109組樣本數據對模型的泛化能力進行驗證。實測值和計算值的相關性如圖4所示。結果表明,測試集擬合度為0.996 9,實測值和計算值兩組數據之間具有較高的相關性。
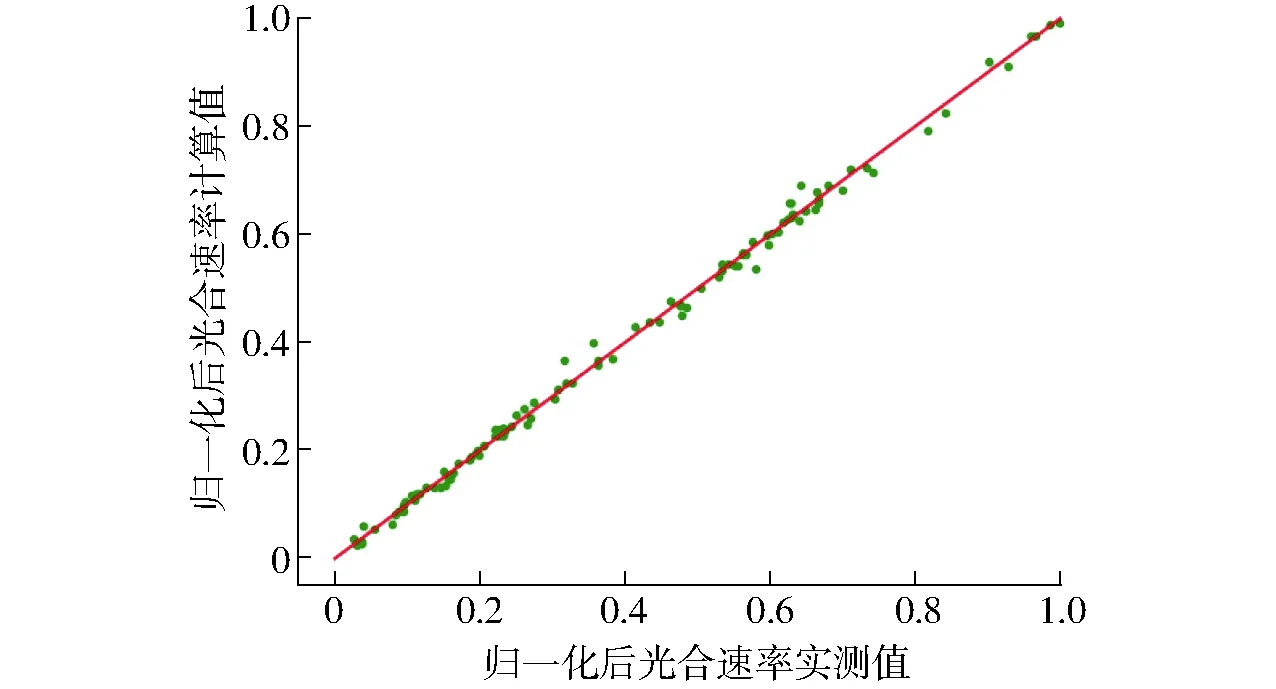
圖4 光合速率預測模型相關性分析Fig.4 Correlation analysis of photosynthetic rate prediction model
2.3 粒子群尋優結果
利用粒子群算法完成了特定溫度下凈光合速率最大點的尋優,其中18、21、24、27、30、33℃條件下的尋優迭代過程如圖5所示。由圖5可知,粒子群算法尋優過程中無振蕩,很快便可以收斂到最優點,證明該方法對于此類樣本尋優適合。得到最優點數據如表2所示。
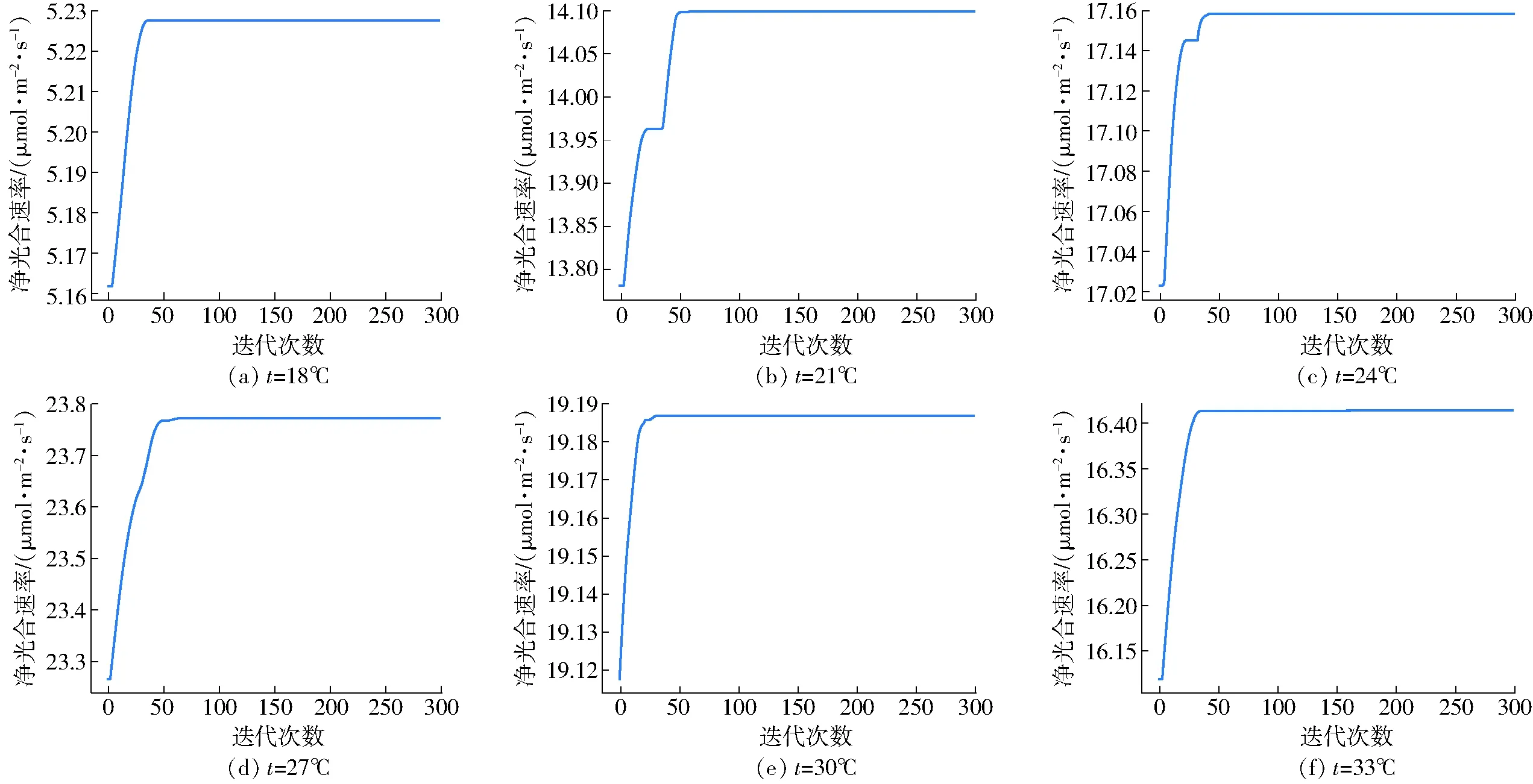
圖5 粒子群算法尋優迭代過程Fig.5 Processes of particle swarm optimization
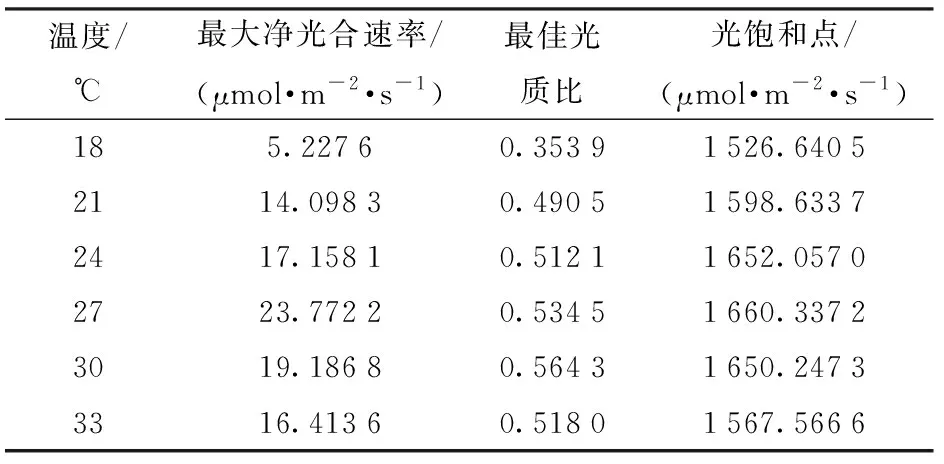
表2 光強與光質比尋優結果Tab.2 Values of optimal points for light intensity and light quality
由表2可知,在不同溫度條件下,最大光合速率、光飽和點以及最佳光質比都有所不同。隨著溫度的升高,3種變量的變化趨勢都是先增后減,其中最大光合速率、光飽和點大約在27℃附近達到最大值,而最佳光質比在30℃附近達到最大值。進一步分析發現,溫度在21~33℃范圍時,最佳光質比約為0.5,但低溫會使最佳光質比較大程度地偏離0.5,18℃時尋找到的最佳光質比只有0.353 9,說明在較低的溫度下藍光可能更有利于提升作物光合能力。
2.4 調控模型驗證
基于光飽和點和最佳光質比尋優結果,利用式(5)、(6)分別計算特定溫度下紅、藍光的需求量,然后利用偏最小二乘回歸法進行擬合,得到紅、藍光的調控曲線如圖6所示,得到的擬合曲線方程分別為
y=-0.138 321 296t3+6.996 413 49t2-
60.851 210 7t+176.653 100
(8)
y=0.071 543 518 5t3-3.925 050 00t2+
41.036 025 0t+1 101.351 70
(9)
紅、藍光調控模型均方根誤差分別為14.635 1、10.192 0 μmol/(m2·s)。
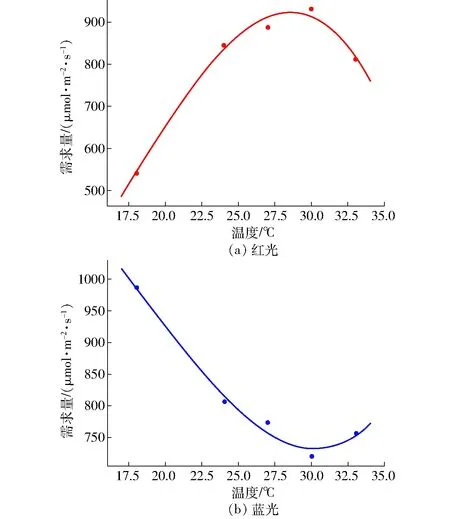
圖6 調控曲線Fig.6 Intelligent regulation model curves
由圖6可知,在紅、藍光的調控模型中,紅光需求量隨溫度的升高而先增后減,大約在28℃附近達到最大值;而藍光需求量隨溫度的升高而先減后增,大約在30℃附近達到最小值。這是因為植物光合作用的主要能量來源于紅光,在28℃附近,黃瓜的光合作用最強,此時所需的紅光光照強度也最多。而刺激植物氣孔張開是藍光的一個重要功能,在低溫和高溫條件下作物會通過關閉氣孔來調節光合作用,所以隨著溫度的升高,所需的藍光光照強度先減小后增大,且在30℃附近,藍光光照強度達到最小,此種變化趨勢可能還與酶的活性等生理參數密切相關[22]。
為了進一步驗證所得調控模型的準確性,本文進行了計算值與實測值相關性分析。通過對實際測量的9個溫度下光響應曲面光合速率最大值尋優得到光飽和點和最佳光質比,并利用式(5)、(6)得到對應溫度下紅、藍光實測值,同時利用式(8)、(9)獲取對應溫度下紅、藍光預測值,從而進行實測值與計算值的相關性分析,結果如圖7所示。
由圖7可知,通過相關性擬合得到的擬合度分別為0.834 5和0.853 5,紅光調控模型相關直線的斜率為1.001 3,截距為1.612 2;藍光調控模型相關直線的斜率為0.985 7,截距為9.526 0,紅、藍光調控模型的均方根誤差分別為15.087 8、10.138 3 μmol/(m2·s),說明該調控模型計算值與實測值之間有較高的相關性,可滿足實際紅、藍光高效調控需求。
2.5 模型調控效果分析
溫度、光照強度、光質比都是影響黃瓜光合速率的重要因素,三者對黃瓜的光合作用存在明顯的互作作用。為了驗證耦合條件下,光質比對光合速率的影響,對比不同文獻中常見的光質比與本模型得到的最佳光質比在相應光飽和點下的凈光合速率,結果如表3所示。
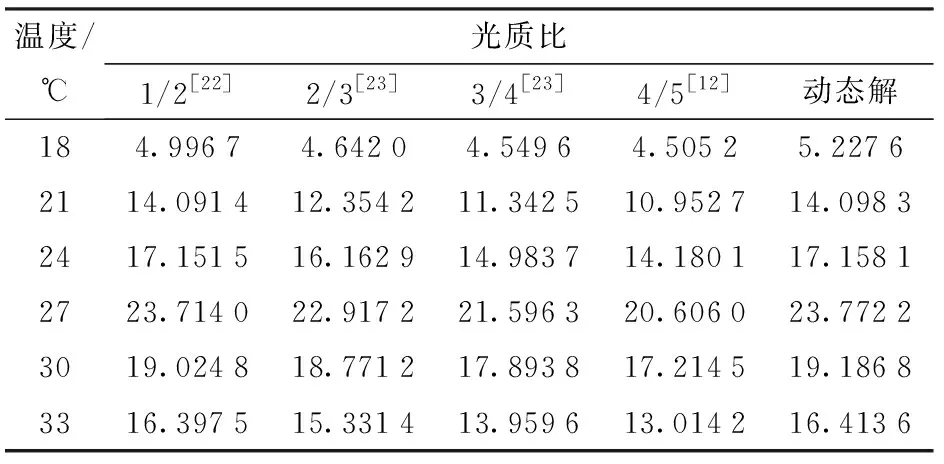
表3 不同光質比對應的凈光合速率Tab.3 Net photosynthetic rates at different light qualities μmol/(m2·s)
由表3可知,在利用粒子群尋優找到的最佳光質比下,黃瓜的凈光合速率高于其他幾種常見的光質配比。在這幾種常用的光質比條件中,光質比為0.5時對應的凈光合速率最高,當光質比超過0.5時,凈光合速率隨光質比的增大而減小。且由表2可知,不同溫度條件下,適合黃瓜生長的最佳光質比不同,最佳光質比隨溫度的升高而先增大后減小。所以相比于0.5這一最優固定光質比,利用粒子群算法動態尋優得到的光質比在相同總光強條件下,其調控效果更優。
3 討論
為了進一步分析不同光質比造成凈光合速率變化的原因,對比了不同溫度下光質比對氣孔導度的影響,如圖8所示。由圖8可知,光照強度、光質比和溫度對氣孔導度有顯著影響,相同光質比條件下,氣孔導度隨溫度變化趨勢與光合速率變化趨勢保持一致,大約在27℃附近達到最大值。從圖8可知,在較低溫度(24℃)及較高溫度(33℃)時,光質比的變化對氣孔導度有顯著影響,增加藍光光強可以顯著地影響氣孔導度變化。27~30℃附近,在最適光質比范圍內,增加藍光光強對氣孔導度的影響很小,可能是由于在此溫度范圍內,光質比不再是影響其變化的主導因素,此時光合速率的下降可能與酶的活性等生理參數更相關。同時,在相同溫度和光照條件下,氣孔導度隨光質比的增加而先增后減[24],且在光質比約為0.5處達到最大值,該變化趨勢與凈光合速率的變化趨勢一致。從而表明最佳光質比的動態變化很有可能是藍光的氣孔導度效應引起的。
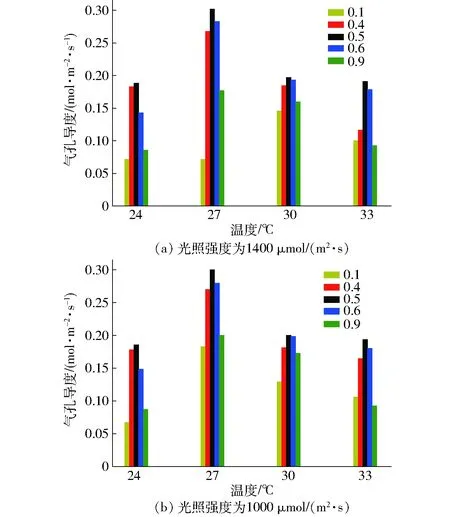
圖8 不同溫度下光質比對氣孔導度的影響Fig.8 Effect of light quality on stomatal conductance at different temperatures
同時,光質比還會通過影響色素活性等生理參數進而影響其光合作用。相關研究發現紅藍光通過光敏色素和隱花色素共同調節光合作用。儲鐘稀等[25]研究了光質對黃瓜葉片光合作用特性的影響,發現紅光處理的葉片具有較高的光系統Ⅱ(PSⅡ)活性和較低的光系統Ⅰ(PSⅠ)活性,藍光處理的葉片具有較低的PSⅡ活性和較高的PSⅠ活性。而PSⅠ和PSⅡ是光合作用重要的兩個過程,所以紅藍光比例過高或過低均會對植物的光合作用造成不利影響。王曉艷等[22]研究了不同比例的紅藍光對弱光下黃瓜葉綠體超微結構和相關光合特性的影響。結果顯示,光質比為0.5處理組的比葉質量、凈光合速率、氣孔導度和光合性能指數均最高,分析得到光質比為0.5最有利于黃瓜幼苗葉綠體的發育和高光合速率的實現。其結果與表2中黃瓜的凈光合速率隨光質比的增大而先增后減,且在光質比約為0.5時達到最大值一致。
4 結論
(1)以溫度、光質比、光照強度為輸入,黃瓜凈光合速率為輸出,建立基于回歸型支持向量機算法的融合黃瓜光質需求設施光環境智能調控模型。驗證結果表明,該模型的擬合度為0.996 9,其均方根誤差為0.436 7 μmol/(m2·s),明顯優于BP算法和隨機森林算法構建的模型,為設施光環境調控提供了重要依據。
(2)基于粒子群算法實現了不同溫度下最佳光質比和光飽和點獲取,其結果表明不同溫度下最適合黃瓜生長的光質比存在差異,最佳光質比隨溫度的升高呈現先增后減的趨勢,大約在30℃附近達到最大值。
(3)建立了紅、藍光目標值調控模型,其均方根誤差分別為15.087 8、10.138 3 μmol/(m2·s),其調控效果明顯優于傳統固定的光質比,可用于指導實際應用中的光環境調控。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
