不同形態氮肥對紫花苜蓿生長、硝酸鹽轉運蛋白基因MtNRT1.3表達及氮吸收的影響
2019-11-07 11:54:36王玉強劉文濤趙國琦孫盛楠
草地學報 2019年5期
關鍵詞:植物
王玉強,沈 宇,錢 進,劉文濤,趙國琦,孫盛楠
(揚州大學動物科學與技術學院,江蘇 揚州 225009)
氮素是植物生長必不可少的大量營養元素,也是植物體內重要有機化合物的結構性元素,如蛋白質、核酸、葉綠素、酶和激素等;同時又是作物高產、品質優良的重要影響因素[1]。土壤中主要存在2種形態氮源,銨態氮和硝態氮,植物主要通過根系吸收土壤中的銨態氮和硝態氮,而不同形態的氮素會影響植物的氮素同化、光合作用以及植物體內陰陽離子的平衡,從而對植物生長產生不同的效應[2-3]。
銨態氮和硝態氮是植物吸收的主要氮素形式,吸收量占吸收陰、陽離子的70%左右[4],銨態氮促進植物吸收陰離子,消耗有機酸;而硝態氮促進植物吸收陽離子,促進有機陰離子的合成。不同植物對不同形態氮素的吸收、轉化利用能力也不同,比如旱地植物一般表現為“喜硝性”,而水生植物或強酸性土壤上生長的植物,則一般表現為“喜銨性”,這是植物為適應土壤環境而長期進化的結果。有研究[5]表明,水稻田施用銨態氮,可以明顯提高植株的株高、地上干物質量,從而促進水稻的生長。而有研究[6]發現,不同形態氮肥對小麥的產量、增產量和增產率的影響中,硝態氮肥最高,硝態氮、銨態氮組合次之,銨態氮最低。
大量研究都著眼于植物對銨態氮與硝態氮的吸收量、吸收速率和光合特性上,忽視了土壤這個具有強大緩沖能力的載體。植物的生長和營養的吸收利用都離不開土壤,土壤的理化性質可以直接影響植物對銨態氮和硝態氮的吸收。有研究[7]發現,在土壤高pH的條件下,不論來自有機質的礦化還是直接施入土壤的銨態氮,會迅速通過硝化作用轉化為硝態氮,土壤中銨態氮濃度低至只有0.77 mmol·dm-3,而硝態氮濃度能高達 6 mmol·dm-3。
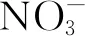
目前研究最多的NRT1轉運體是擬南芥AtNRT1.1蛋白。硝酸鹽可以誘導其表達[24]。AtNRT1.1在功能上與硝酸鹽對根生長的刺激作用有關[25,26]。AtNRT1.1已被證明在硝酸鹽感應系統中發揮了作用,該系統通過促進根尖中生長素的流動來刺激根系生長[27,28]。迄今為止,豆科植物中的硝酸鹽轉運體很少受到關注,可能是因為這些物種能夠通過共生結瘤的生物固氮來適應氮饑餓[29]。盡管硝酸鹽在豆科植物中也起著重要的營養和信號作用,但在這些物種中只有少數假定的硝酸鹽轉運體被克隆出來[30],到目前為止,它們都沒有功能特征。事實上,在氮限制條件下,豆科植物能夠吸收礦物質氮,尤其是土壤中硝態氮,以滿足其營養需求,然后才能分化出功能共生結瘤。此外,有研究[31]表明,硝酸鹽通過局部和系統性調節途徑,抑制根結瘤和固氮活性。在給植物接種根瘤菌的試驗中,氮輸入被證明能夠調節百脈根(Lotusjaponicus)中根瘤的發生[32]。硝酸鹽轉運蛋白基因MtNRT1.3主要在紫花苜蓿根中表達,編碼雙親和硝酸鹽轉運體,類似于擬南芥中的AtNRT1.1[33]。有研究發現蒺藜苜蓿中的MtNRT1.3與描述的NRT1家族成員不同,根中MtNRT1.3的表達受氮饑餓刺激,并在含有NO3-的培養基中受到抑制[34]。
紫花苜蓿是現代畜牧業生產中重要的蛋白飼草資源,本研究以紫花苜蓿為研究對象,探究紫花苜蓿對不同形態氮素的吸收、轉化和土壤微環境對外源氮素添加的響應,以及氮素、植物和土壤三者之間的相互關系。探討不同形態氮肥對土壤氮含量及植物根系養分吸收的影響,了解土壤氮素與植物供求關系,揭示不同形態氮素對紫花苜蓿生理特征、氮含量的影響,通過平衡銨態氮和硝態氮含量,合理調控氮肥種類和養分供求以滿足作物要求,提高資源利用效率和作物生產力。
1 材料與方法
1.1 試驗材料
供試紫花苜蓿品種為WL233(購于北京正道種業有限公司),供試肥料包括:分析純硝酸鈉、氯化銨和硫酸鉀;化學純化過磷酸鈣,有效磷(以P2O5計)為14%~16%。
盆栽土壤取自揚州大學草學智能溫室試驗地0~30 cm的表土,有機質19.08 g·kg-1,全氮0.85 g·kg-1,pH值8.32,銨態氮4.83 mg·kg-1,硝態氮12.48 mg·kg-1。
1.2 試驗設計
試驗采用盆栽法,于2019年4月在揚州大學草學智能溫室內進行。試驗選擇容器為18 cm×17 cm×20 cm的塑料花盆,每盆裝土3.5 kg,根據土壤基礎肥力測定,每盆按照1 kg土施加K2O 200 mg、P2O5300 mg作為基肥。每盆均勻點播10粒種子,待幼苗長出第3片真葉時進行間苗,每盆留8株長勢一致的健康幼苗。
試驗設計采用單因素完全隨機設計,共設4個處理,分別為以不施氮處理為對照組(Con)、銨態氮(NH4Cl-T1)、硝態氮(NaNO3-T2)和混合氮(銨態氮、硝態氮1︰1混合-T3),每個處理重復4次,共16盆。各形態氮肥施加總量為250 mg·kg-1(每1 kg土),以Fahraeus無氮營養液[35]結合NH4Cl和NaNO3配置所需氮素濃度,3葉期時開始澆灌,每次澆灌營養液200 mL,每周澆一次營養液,并進行日常澆水、除雜防護管理,共計5周。現蕾期取樣測定各個指標。
1.3 測定指標和方法
1.3.1土壤pH及土壤氮含量的測定 用常規農化分析方法測定pH值(Sartorious PB-10)[36]。采用試劑盒(購于蘇州科銘生物科技有限公司)測定土壤銨態氮和硝態氮含量。
1.3.2紫花苜蓿生長性狀及植株銨態氮和硝態氮含量測定 株高:測量植株的自然狀態下的垂直高度;根長:將植株根系整個挖出,去除土壤(保證根的完整),用直尺測量主根的根長;所有處理地上部和地下部(蒸餾水洗凈,吸水紙吸干)稱取鮮重,留一小部分用于測定植株銨態氮、硝態氮含量和根中MtNRT1.3 基因的表達(—80℃保存),其余部分于烘箱105℃殺青30 min后,在75℃烘干至恒重,稱取干質量即為生物量;
植物銨態氮和硝態氮含量:采用試劑盒測定(購于蘇州科銘生物科技有限公司)。
1.3.3紫花苜蓿根RNA的提取和cDNA的合成 稱取1~2 g紫花苜蓿根,用磷酸緩沖鹽溶液(Phosphate buffer saline,PBS)洗凈,吸水紙吸干,放入玻璃研磨器中,接著加入1 mL Trizol(細胞和組織總RNA提取試劑)研磨至勻漿,裝于1.5 mL指形管中,加入0.2倍體積的氯仿,離心20 min,取上清,加入等體積的冰異丙醇,—20℃冰箱過夜沉淀。
將之前過夜沉淀的混合液4℃,12 000 rpm離心20 min;棄去上清,加入750 μL的75%乙醇(焦碳酸二乙酯無酶水配制),繼續4℃,12 000 rpm離心20 min;重復上一步驟,在超凈工作臺中將指形管內水分晾干,根據得到的RNA團塊大小加入適量的DEPC水,65℃水浴10 min,然后放回冰上,測量RNA濃度。取1 μLRNA溶液,在260 nm和280 nm測定吸光度值,并計算A260/A280,檢測RNA的純度。
用Thermo Script反轉錄酶(Invitrogen)和OligodT18 Primer對2 μg的RNA樣品進行反轉錄,反應溫度為42℃,合成cDNA。
1.3.4qPCR法測定紫花苜蓿MtNRT1.3基因的表達 引物及內參基因見表1,使用SYBR Premix Ex Taq (Perfect Real Time)Kit(TaKaRa Biotechnology (Dalian)Co.,Ltd.)試劑盒進行qPCR定量測定。反應體系為20 μL:SYBR Green Super Mix (2×)10 μL,稀釋過的cDNA 2 μL,正反引物各 0.8 μL,用超純水補齊。計算采用2-ΔΔCt法計算基因的相對表達量。

表1 實時熒光定量PCR引物信息
1.4 數據處理
采用Microsoft Excel 2015 和SPSS 19.0 進行數據處理及方差分析,Sigmaplot 10.0進行作圖。
2 結果與分析
2.1 不同形態氮肥對土壤pH值和銨態氮和硝態氮含量的影響
由圖1A可得,施有硝態氮肥的T2和T3處理組土壤pH值顯著低于T1和對照組(P<0.05),單施銨態氮T1處理組與對照組相比差異不顯著。土壤中銨態氮和硝態氮含量直接關系到植物從土壤可以獲得的氮含量。從圖1B可以看出,不同處理對土壤氮含量有顯著影響,T2處理土壤中銨態氮含量顯著高于其他各組,T3處理組顯著低于其他各處理組(P<0.05),T1處理與對照組相比無顯著差異(P>0.05)。由圖1C可以看出,T2處理土壤中硝態氮含量也顯著高于其他各處理組,與圖1B土壤中銨態氮含量不同的是T3處理硝態氮含量顯著高于T1和對照組,土壤中硝態氮含量依次為T2>T3>T1>Con,且各組間差異顯著(P<0.05)。

圖1 不同施肥處理對土壤pH及土壤銨態氮和硝態氮含量的影響
2.2 不同形態氮肥對紫花苜蓿株高、根長及生物量的影響
由圖2B可以看出,不同形態氮肥處理下,紫花苜蓿根長差異不顯著(P>0.05),但是株高有顯著差異。只施硝態氮肥的T2處理下紫花苜蓿株高顯著高于對照組和混合氮T3處理組,其他各組間差異不顯著(圖2A)。

圖2 不同施肥處理對紫花苜蓿株高及根長的影響
施加氮肥能夠有效提高植物的生物量,由圖3A可知,各施肥處理組地上生物量顯著高于對照組,且單施銨態氮和硝態氮地上生物量顯著高于混合氮T3處理組(P<0.05);單施銨態氮肥T1處理組地下生物量顯著高于對照組和混合氮T3處理組(P<0.05),但與單施硝態氮肥組T2處理組相比差異不顯著(P>0.05)(圖3B);從圖3C根冠比可以看出,單施硝態氮肥組根冠比顯著低于對照組(P<0.05),值得注意的是,對照組根冠比的比值大于其他各處理組,T1處理根冠比比值大于T3處理組,T3處理大于T2處理(P>0.05)。
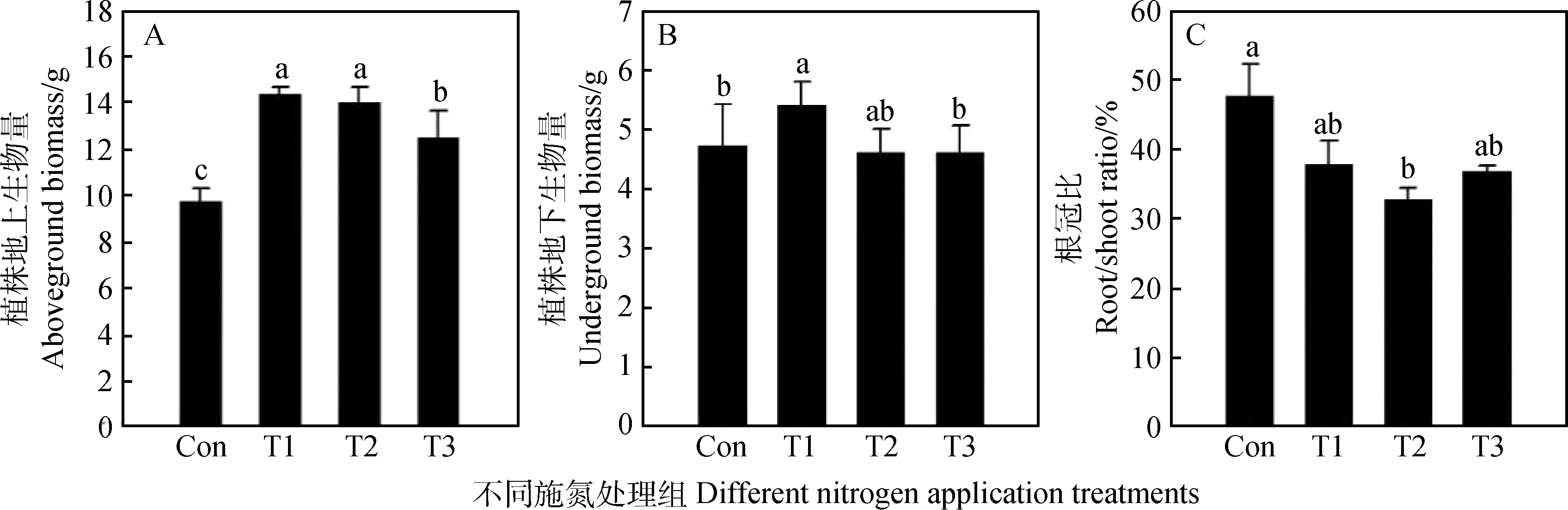
圖3 不同施肥處理對紫花苜蓿地上、地下生物量及根冠比的影響
2.3 不同形態氮肥對紫花苜蓿植株銨態氮和硝態氮含量的影響
不同形態氮肥處理對紫花苜蓿植株銨態氮和硝態氮含量有顯著影響。由圖4A可知,混合氮T3處理下植物銨態氮含量顯著高于其他各處理組,且各組間差異顯著(P<0.05),植物銨態氮含量在不同處理間依次升高(Con

圖4 不同施肥處理對紫花苜蓿地上部銨態氮和硝態氮含量的影響
2.4 不同形態氮肥對硝酸鹽轉運蛋白基因MtNRT1.3表達的影響
硝酸鹽轉運蛋白基因MtNRT1.3主要在紫花苜蓿根中表達,圖5可以看出,Con組MtNRT1.3基因的表達量顯著低于其他處理組;單施硝態氮肥的T2處理該基因的表達量最高,MtNRT1.3基因的表達量顯著高于T1和T3處理組(P<0.05);T1和T3處理組間無顯著差異(P>0.05)。
2.5 紫花苜蓿生理特征和MtNRT1.3與土壤氮含量的關系
由表2可以看出,土壤銨態氮含量與基因MtNRT1.3、株高和土壤pH呈顯著正相關關系(P<0.01);土壤硝態氮與葉銨態氮、地上生物量和基因MtNRT1.3呈顯著正相關關系(P<0.01),但與根冠比呈顯著負相關關系(P<0.01)。

圖5 不同施肥處理對紫花苜蓿硝酸鹽轉運蛋白基因MtNRT1.3表達量的影響
3 討論
3.1 添加不同形態氮素對土壤理化性質的影響
土壤具有良好的緩沖能力,一般都能維持穩定的土壤微環境,但是有研究表明,不同施肥處理會打破土壤穩定的pH環境[37]。植物可以直接利用土壤中的銨態氮和硝態氮,根系吸收陰陽離子是平衡的,根系吸收銨態氮時就會向土壤中釋放H+使土壤pH值下降;吸收硝態氮時,向土壤釋放OH-致使土壤pH值升高。本試驗銨態氮處理和混合氮處理降低了土壤pH值,硝態氮處理數值上提高了土壤pH值,但是與對照相比差異不顯著,這說明施加銨態氮處理植物吸收銨態氮較多,從而降低了土壤pH[38,39]。圖1B T1處理組銨態氮含量與對照組無顯著差異,說明單施銨態氮肥對土壤銨態氮含量影響較小,這與王西娜等[40]的研究一致,這也說明銨態氮更容易被植物吸收利用;T3處理土壤中銨態氮和硝態氮含量顯著低于T1、T2處理,說明混合施加硝態氮和銨態氮更有利植物根系對土壤氮素的吸收利用[41]。圖1C T1處理的硝態氮含量顯著高于對照組,說明施入土壤的銨態氮很可能被轉化為硝態氮,從而導致土壤硝態含量升高。施入土壤中的銨根離子為陽離子,容易與土壤膠粒吸附結合,從而被硝化微生物硝化為硝酸根或亞硝酸根離子存在于土壤中[42]。

表2 紫花苜蓿生理特征及MtNRT1.3與土壤氮含量的關系
注:表中數值為Pearson相關系數,**表示相關性顯著,P<0.01
Note:The values are Pearson correlation coefficients in the table. ** Significant correlation,P< 0.01
3.2 紫花苜蓿的生長及對不同形態氮素的吸收利用
植物的生長離不開氮素,施加氮肥能夠提高植物產量。本試驗如圖2A所示單施硝態氮肥提高了紫花苜蓿的株高,說明硝態氮與銨態氮相比更有利促進紫花苜蓿的高度生長。由圖3B,3C可知施氮處理明顯提高了紫花苜蓿地上、地下生物量,這結果與Hojjati等[43]研究結果一致。單施銨態氮肥使得植株地上、下生物量最高,說明銨態氮更容易被紫花苜蓿吸收利用。根系是植物吸收利用養分的關鍵部位,根冠比的大小反映了植物干物質積累方向及地上和地下的相互關系。有研究[44]表明,養分供應不足時,根系的側根數量和根毛會增加來尋找更多的養分供應植物生長,光合產物會優先配送到根部,根冠比增大,本試驗的結果與其一致,說明施加氮肥能夠降低植物的根冠比,有利于植物地上物質的積累。本試驗還發現,單施硝態氮肥處理根冠比最低,結合圖3B T2處理與T1處理地下生物量數據得出硝態氮可能抑制了紫花苜蓿根系的生長,光合產物優先向地上部分運輸。
測定紫花苜蓿中銨態氮和硝態氮含量時,本試驗發現(圖4A,4B)混合氮處理組的植株的銨態氮和硝態氮含量均較高,且兩者含量都呈現一種趨勢,Con<處理T1<處理T2<處理T3處理。本研究結果表明施肥能夠增加植株內2種氮素的含量,單一施銨態氮和硝態氮并不會引起植物內單一氮素含量增加,可能是因為2種氮素的含量在植物體內是相對平衡的,銨態氮和硝態氮之間存在相互轉化的過程。有研究[45]表明,植物液泡中累積的硝酸鹽可以作為內源養分,在外源氮素不足時會被植物體吸收轉化利用。由圖4可以得出,2種氮素配合施加更有利于氮素在體內的積累,且硝態氮處理比銨態氮處理更有利氮素在植物體內積累,施用硝態氮肥不僅能使紫花苜蓿吸收更多的硝態氮,還能較快地將其轉化為銨態氮積累在體內。本研究結果顯示,紫花苜蓿宜混合施用銨態氮和硝態氮肥且以硝態氮比例較大為佳。
3.3 不同形態氮素對紫花苜蓿硝酸鹽轉運蛋白基因MtNRT1.3的影響
不同形態氮素對紫花苜蓿根的刺激反應也不同,植物為了更好地吸收利用氮素資源,形成了特定的硝酸鹽吸收同化途徑。由圖5可以看出,不同施肥處理上調了硝酸鹽轉運蛋白基因MtNRT1.3的表達,其中單施硝態氮肥T2處理該基因的表達量最高,說明硝酸鹽誘導了MtNRT1.3的表達,該基因編碼的硝酸鹽轉運蛋白參與了紫花苜蓿根對硝酸鹽的吸收轉運。同時,在T2處理中,紫花苜蓿葉片中的硝態氮含量最高,說明該基因表達量的增加與植株對氮的吸收密切相關。此外,T1處理下MtNRT1.3的表達量也顯著高于對照組,可能是由于部分銨態氮轉化成硝態氮,從而促進了該基因的表達。在研究豆科模式植物蒺藜苜蓿NRT1.3基因的試驗中,發現MtNRT1.3參與了氮素限制反應,提高了植株在氮素限制條件下獲得NO3-的能力[34]。Anthoni Pellizzaro等[46]發現,MtNRT1.3受脫落酸的刺激,并參與對蒺藜苜蓿主根生長的調節。上述試驗雖然都發現MtNRT1.3與硝酸鹽轉運有關,但是其結果與本試驗結果不同,本試驗發現在高濃度外源氮素條件下,施加不同形態氮肥使該基因的表達量顯著增加;與已有結論[34,46]相似的是,硝態氮肥可能會抑制植物地下部分的生長,這可能與氮肥來源、植物生長環境、土壤氮含量等因素有關。
3.4 紫花苜蓿生理特征、MtNRT1.3表達與土壤氮含量的關系
植物和土壤兩者在自然環境中存在著特定的有機聯系,當外界環境發生變化時,植物與土壤之間會產生不同響應。如本試驗表2所示,紫花苜蓿MtNRT1.3基因的表達與土壤中銨態氮、硝態氮呈正相關性,這說明當土壤中銨態氮和硝態氮誘導了紫花苜蓿MtNRT1.3基因的表達,有研究[28-29]證實MtNRT1.3類似于擬南芥的AtNRT1.1,AtNRT1.1已被證明參與雙親和性硝酸鹽轉運系統,在硝酸鹽感應系統中發揮了作用,該系統通過促進根尖中生長素的流動來刺激根系的生長[27,28],但是本試驗發現土壤中銨態氮含量卻與其具有正相關性,可能是因為土壤中過多的銨態氮在土壤硝化細菌的作用下轉化為硝態氮,從而影響MtNRT1.3的表達。所以,當紫花苜蓿根中MtNRT1.3基因的表達量增加時,植物可以通過根系更有效地分配和利用土壤中的氮素,從而提高植株株高,增加地上生物量。通過相關分析還發現,土壤硝態氮的含量與植物根冠比呈負相關性,與地上生物量呈正相關性,說明硝態氮肥更有利于紫花苜蓿把光合產物優先向地上組織中運送。
4 結論
施氮處理提高了紫花苜蓿植株中的氮含量,施加不同形態氮肥均提高了紫花苜蓿根中MtNRT1.3基因的表達量,且該基因的表達量與土壤中銨態氮和硝態氮含量呈正相關性;相比于銨態氮肥,施加硝態氮肥不僅增加了植株硝態氮含量,而且提高了植株銨態氮含量;相比于單施硝態氮和銨態氮肥,混合氮肥對提高植物氮含量效果最好;施加硝態氮肥更有利于紫花苜蓿地上部分生物量的累積;因此,對紫花苜蓿施加混合態氮肥,尤其是增加硝態氮肥的比例更有利于植株的生長和對氮素的吸收。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
