麋鹿的分類地位與遺傳多樣性研究概述
2019-11-19 08:26:14張樹苗白加德李夷平梁兵寬李俊芳李俊生
野生動物學報 2019年4期
關鍵詞:物種
張樹苗 白加德 李夷平 陳 頎 梁兵寬 李俊芳 李俊生 常 江*
(1.北京麋鹿生態實驗中心,北京,100076;2.國家林業和草原局調查規劃設計院,北京,100714;3.國家環境保護區域生態過程與功能評估重點實驗室,中國環境科學研究院,北京,100012)
麋鹿(Elaphurusdavidianus)是世界自然保護聯盟(IUCN)紅皮書野外滅絕物種,是我國I級重點保護野生動物,是國內外物種遷地保護的經典案例,是世界物種重引進項目的成功范例。野生麋鹿種群在經歷商周的鼎盛時期之后走向衰落,至十九世紀末,麋鹿在我國本土滅絕[1]。1893—1895年,英國第十一世貝福特公爵收集了18只散落在歐洲各地的麋鹿,集中飼養在其烏邦寺莊園里,這些麋鹿成了全球現存麋鹿的祖先“奠基者”[2]。第一次世界大戰后,烏邦寺的麋鹿種群日漸擴大。為挽救瀕危物種,保護生態平衡,實現麋鹿回歸故鄉,中國于1985—1987年共從英國引入麋鹿77只,分別飼養于北京南海子麋鹿苑和江蘇大豐麋鹿國家級自然保護區,建立了麋鹿半散放種群,拉開了麋鹿在我國遷地保護的序幕[3]。經過30多年的努力,目前,中國麋鹿種群數量已超過7 000只,麋鹿棲息地較為穩定,暫未發現有明顯的生存力和繁殖力下降的情況,是中國乃至全球珍稀物種保護領域中的成功范例。但是,麋鹿在近代經歷了4次遺傳瓶頸事件,使其群體遺傳變異嚴重丟失。第一次是英國烏邦寺建群時,第二次是英國各動物園建群時,第三次是北京南海子麋鹿苑和江蘇大豐麋鹿國家級自然保護區引入建群時,第四次是從南海子和大豐引入到湖北天鵝洲建群時[4]。目前,基于分子生物學的研究顯示,麋鹿中性分子標記遺傳多樣性較低,且高度近親繁殖,這個遺傳困境備受學術界、野生動物保護工作者與管理部門的關注與重視。雖然我國的麋鹿種群發展穩定,且還會繼續增長,在保護生物學領域創造了物種重引進的奇跡。但不能忽視,麋鹿的近交系數極高,奠基者數量較少,仍然面臨嚴峻的遺傳困境,在種群管理方面需要避免高度近交和遺傳多樣性減少與喪失。Clutton-Brock等[5]指出大型草食動物的種群管理應考慮實施人工種群調控措施,Foose和 Frankham等[6-7]指出瀕危物種的保育要實施遺傳管理措施。本文介紹麋鹿分類地位和麋鹿遺傳多樣性的有關研究,為中國麋鹿遺傳資源保護及種群的健康可持續發展和管理提供參考。
1 麋鹿分類地位
麋鹿是中國特有的動物種類,麋鹿屬曾被認為有5種,其中雙叉麋鹿(E.bifurcatus)、晉南麋鹿(E.chinanensis)、藍田麋鹿(E.lantianensis)和臺灣麋鹿(E.formosanus)4種已滅絕,現僅存達氏麋鹿(E.davidianus),也即大衛鹿[1,8-9]。
脊索動物門Chordata
脊椎動物亞門Vertebrata
哺乳綱Mammalia
偶蹄目Artiodactyla
反芻亞目Ruminantia
鹿上科Cervoidea
鹿科Cervidae
鹿亞科Cervinae
麋鹿屬
Elaphurus
達氏種
E
.
davidianus
雙叉種
E
.
bifurcatus
晉南種
E
.
chinanensis
藍田種
E
.
lantianensis
臺灣種
E
.
formosanus
圖1麋鹿的分類地位(引自白加德,2014)[8]
Fig.1 The classification of Père David’s deer(Cited from Bai Jiade,2014)[8]
達氏種是1866年由愛德華(Milne-Edwards A.)根據法國傳教士大衛從北京南苑帶到法國巴黎博物館的標本研究建立的。其屬名Elaphurus為長尾之意,指出麋鹿尾長這一鮮明特征。種名“davidianus”是紀念第一個發現者大衛的,所以西方人習慣地稱麋鹿為“大衛鹿”(David’s deer)。在中國,很少這樣稱呼。麋鹿是中國學術界沿用甲骨文的稱謂,老百姓稱麋鹿為“四不像”,近三四十年來,麋鹿一詞已被普及[9]。
雙叉種(E.bifurcatus)是1930年,德日進和皮韋托[10]在研究中國河北泥河灣拉方期哺乳動物群時,發現一類鹿角化石,分叉性質與現生麋鹿相類似,建立的一個新種。
1933年,Sowerby[11]根據河南安陽殷墟的發現,在真鹿屬(Cervus)的名下記述了一個新種,稱為真鹿屬梅氏種(Cervus(Rucervus)menziesianus),后被修改劃入麋鹿屬中,叫麋鹿屬梅氏種(E.menziesianus)[10],與現生種麋鹿相并列。1955年楊鐘健[12]、1956年裴文中[13]指出梅氏種是達氏種的同物異名,梅氏種不能成立。于是,根據國際動物命名法規優先權法則,梅氏種已經統一于1866年愛德華對現生種的命名。
晉南種(E.chinanensis),是1974年賈蘭坡根據山西早更新世的考古材料確定的古麋鹿新種[14]。
藍田種(E.lantianensis),是1975年計宏祥根據陜西早更新世的材料確定的另一古麋鹿新種[15]。
1978年,日本人大裕之等修定臺灣早更新世的狍屬臺灣種(Capreolusformosanus)為麋鹿屬臺灣種(E.formosanus)。
2018年,Dong等[16]對產自山西天鎮的鹿角標本進行了詳細研究,發現了麋鹿的兩個種,其中一個是雙叉麋鹿,另外一個由于形態的不同,建立了麋鹿種之下的一個亞種——原達氏鹿Elaphurusdavidianuspredavidianus。他們認為鹿科麋鹿屬歷史曾共有4個種8個亞種,其中達氏麋鹿有4個亞種,雙叉種有2個亞種,晉南種有2個亞種,豐富了我們對麋鹿屬物種多樣性的認識。
關于現存達氏麋鹿的分類地位,學術界存在不同意見。盛和林[17]和Simpson[18]認為,麋鹿屬于鹿科(Cervidae)鹿亞科(Cervinae)麋鹿屬(Elaphurus),與Grubb,Geist和白加德等認為麋鹿是獨立的屬觀點相一致[8,19-20]。

反芻亞目Ruminantia 鼷鹿上科Tragulodea 鼷鹿科Tragulidae 鹿上科Cervoidea 麝科Moschidae 麝屬Moschus 鹿科Cervidae 獐亞科Hydropotinae 獐屬Hydropotes 鹿亞科Cervinae 麂族Muntiacini 毛冠鹿屬Elaphodus 麂屬Muntiacus 鹿族Cervini 斑鹿屬Axis 鹿屬Cervus 麋鹿屬Elaphurus反芻亞目Ruminantia 鼷鹿次目Tragulina 異鼷鹿科Hypertragulidae 鼷鹿科Tragulidae 有角次目Pecora 鹿科Cervidae 麝亞科Moschinae 麝屬Moschus 麂亞科Muntiacinae 麂屬Muntiacus 毛冠鹿屬Elaphodus 鹿亞科Cervinae 斑鹿屬Axis 黇鹿屬Dama 鹿屬Cervus 麋鹿屬Elaphurus
圖2反芻亞目下的分類系統(引自盛和林,1992;Simpson,1945)[17-18]
Fig.2 The classification of Ruminantia(Cited from Sheng Helin,1992;Simpson,1945)[17-18]
但有學者認為麋鹿應該并入鹿屬(Cervus)。Randi等[21]基于線粒體D-loop全序列分析,探討了鹿科、鹿亞科25個種和亞種的系統發生關系,首次提出將麋鹿并入鹿屬。Gilbert等[22]認為鹿亞科分為4個屬,鹿屬、斑鹿屬(Axis) 、黇鹿屬(Dama)和沼鹿屬(Rucervus),麋鹿歸入鹿屬,Cronin等[23-24]和Randi等[25]把鹿亞科分為 3 個屬,認為麋鹿應劃入鹿屬。劉向華等[26]根據線粒體細胞色素b基因(Cytb)序列及部分片段構建的系統發育樹探討鹿亞科屬間屬內進化關系,分析結果也認為麋鹿應并入鹿屬,涂劍鋒等[27]研究鹿亞科動物線粒體DNA控制區全序列,構建的系統發育樹結果表明,麋鹿屬的麋鹿、斑鹿屬的豚鹿(Axisporcinus)以及黇鹿屬的黇鹿(Damadama)與鹿屬的分化處于屬間差異,支持將其并入鹿屬,匡葉葉[28]在“建立AFLP分子標記檢測體系”的基礎上,通過對北京南海子245只麋鹿的AFLP檢測,基于遺傳相似系數矩陣,結果表明,麋鹿應屬于鹿屬、Emerson[29]也認為麋鹿與鹿屬進化關系較近,應并入鹿屬。
2 麋鹿與鹿亞科動物間的進化關系
物種間的進化關系反映了物種的系統演變歷程及親緣關系,對物種的保護遺傳學研究有重要的意義。鹿亞科是大、中型的鹿類,按照盛和林[17]對鹿亞科的分類方式,鹿亞科分成:黇鹿屬(Dama)、斑鹿屬(Axis)、麋鹿屬(Elaphurus)和鹿屬(Cervus)。中國的鹿亞科動物包括:鹿屬的梅花鹿(Cervusnippon)、馬鹿(C.elaphus)、坡鹿(C.eldii)、水鹿(C.unicolor)和白唇鹿(C.albirostris),斑鹿屬的豚鹿和麋鹿屬的麋鹿(Elaphurusdavidianus)7個物種,其中麋鹿與白唇鹿是我國的特有物種(表1)。古生物化石研究表明,自中新世早期鹿亞科從鹿科總進化枝分化出來后,麋鹿屬是最早從鹿亞科分離出來的進化枝[30],現存麋鹿是這一屬中高度特化的物種。
表1 中國鹿亞科動物起源、保護等級及國內分布

Tab.1 The origin,protection level and distribution of Cervinae animals in China
關于麋鹿與鹿亞科其他鹿的進化關系,學術界有多派觀點。較多數學者認為,相比鹿亞科其他屬,麋鹿屬與鹿屬有較近的進化關系。Grove等[31]對鹿科動物的共源性狀進行了研究,結果顯示麋鹿屬和鹿屬之間有較近的進化關系,這與對鹿科動物染色體核型研究的結果一致;國外學者利用分子生物學分析揭示,麋鹿更接近于鹿屬Cervus[32-34]。Emerson和Tate[29,32]對鹿亞科4個屬10個種和亞種的進化關系進行了蛋白電泳分析,發現麋鹿、馬鹿和梅花鹿的遺傳關系較近,結果亦支持這一觀點,并認為麋鹿和其他鹿類角的形態差異并非長時間進化分歧造成的。
部分學者認為麋鹿與馬鹿進化關系近。Maqbool[35]等研究了麋鹿和馬鹿雜交后代的遺傳連鎖圖譜以及其子代的數量性狀座位(QTL),從遺傳學角度反映二者具有較近的進化關系。
也有學者認為麋鹿與坡鹿、澤鹿(Rucervusduvaucelii)進化關系近。劉向華等[26]對鹿亞科 4 個屬的馬鹿東北亞種(C.elaphusxanthopygus)、甘肅亞種(C.elaphuskansuensis)、阿爾泰亞種(C.elaphusasiaticus)、西藏亞種(C.elaphuswallichii)、四川亞種(C.elaphusmacneilli)、天山亞種(C.elaphussongaricus)、四川梅花鹿(C.nipponsichuanicus)、白唇鹿、水鹿、坡鹿、豚鹿、印度斑鹿(Axisaxis)、麋鹿及鼷鹿科(Tragulidae)大鼷鹿(Tragulusnapu)共 15 個種和亞種的Cytb基因部分序列做了分析比較,分析結果認為麋鹿與坡鹿形成姊妹關系(PP=97;BP=89),具有較近的親緣關系;盛桂蓮等[36]對麋鹿、黇鹿、豚鹿、馬鹿、水鹿、坡鹿、白唇鹿、梅花鹿、大鼷鹿的Cytb部分同源序列進行的系統發育分析來看,坡鹿與麋鹿聚成姊妹群,反映了二者具有較近的進化關系,這與Emerson基于蛋白質的研究結論一致。孟浩等[37]對鹿亞科的麋鹿、馬鹿、坡鹿、澤鹿的mtDNA控制區序列進行比較,構建NJ分子系統發育樹,發現麋鹿與坡鹿、澤鹿同屬于一個分支,親緣關系最近;涂劍鋒等[27]對13種鹿亞科動物東北馬鹿、甘肅馬鹿、蒙古馬鹿(C.elaphusalashanicus)、阿爾泰馬鹿、天山馬鹿、四川馬鹿、塔里木馬鹿(C.elaphusyarkandensis)、歐洲馬鹿、東北梅花鹿(C.nipponhortulorum)、臺灣梅花鹿(C.nippontaiouanus)、越南梅花鹿(C.nipponpseudaxis)、海南水鹿(C.unicolorhainana)、印度水鹿(C.unicolorniger)、臺灣水鹿(C.unicolorswinhoii)、海南坡鹿(C.eldiihainanus)、泰國坡鹿(C.eldiisiamensis)、印度坡鹿(C.eldiieldii)、白唇鹿、麋鹿、豚鹿、黇鹿、毛冠鹿(Elaphoduscephalophus)的D-loop區全序列,并結合GenBank 檢索到的12種鹿亞科動物同源序列開展分析,構建的系統發育樹,表明麋鹿屬的麋鹿、斑鹿屬的豚鹿以及黇鹿屬的黇鹿與鹿屬的分化處于屬間差異,麋鹿與鹿屬的坡鹿互為姐妹枝,親緣關系最近。
此外,匡葉葉[28]以麋鹿、林麝(Moschusberezovskii)、黑麂(Muntiacuscrinifrons)和豚鹿為研究對象,通過AFLP分子標記,檢測遺傳多樣性的AFLP,基于遺傳相似系數矩陣,構建聚類分析,發現麋鹿與豚鹿、梅花鹿的親緣關系最近。
還有學者認為麋鹿與黇鹿進化關系近。李文斌等[38]利用15個微衛星標記對麋鹿、黇鹿、梅花鹿、赤麂(Muntiacusmuntjak)、馬鹿5種鹿類動物群體DNA多態性進行測定,結果發現黇鹿和麋鹿為一類或各自成為一類。
3 麋鹿的遺傳多樣性研究進展
遺傳多樣性是生物多樣性的核心問題,與生物多樣性的形成、消失和發展休戚相關[39]。遺傳多樣性是評價物種進化潛能與健康狀況,以及物種抵御環境變化、生存壓力能力強弱的一個重要指標[7,40-41]。遺傳多樣性的研究工作是生物多樣性就地保護的基礎,更是遷地保護計劃的關鍵與支撐[42-43]。
從保護遺傳學角度看來,物種的長期生存和發展依賴于其遺傳多樣性和進化潛力的保持,遺傳多樣性的降低及喪失勢必對物種的生存帶來不利影響,導致種群生存能力下降和種群退化,甚至滅絕。種內遺傳多樣性或變異性愈豐富,物種對環境變化的適應能力愈大,其進化的潛力也就愈大。一般來說,瀕危物種相比非瀕危的廣布種,具有較低的遺傳多樣性。生物種群遺傳變異水平和群體遺傳結構是其進化歷史、分布范圍、生活型、繁育方式、擴散機制等各種不同因素綜合作用的結果,與其適應性和進化潛力密切相關。特別對于小種群而言,物種遺傳多樣性主要受到遺傳漂變、有效種群大小、瓶頸效應和奠基者效應等因素影響。因此,最大限度地保護種群現有的遺傳多樣性就顯得尤為重要。
我國麋鹿遺傳多樣性的研究是隨著分子標記的發展而逐步深入的,但過去一致的結論是,麋鹿種源的遺傳多樣性極低[44]。烏邦寺莊園最初的18只麋鹿,性比記載為8雄10雌,至少2只雌性沒有參加繁殖[45-46]。根據于長青[44]的推斷,當時烏邦寺參與交配繁殖的雄性麋鹿應為2只左右,即現存麋鹿應是最初約 10 只麋鹿的后裔。也有學者認為,在英國烏邦寺出生的子一代麋鹿可能都是同一只雄性麋鹿的后代[44],它們的遺傳變異非常單一。1945年,烏邦寺的麋鹿開始向其他地區擴散,Foose等[6]認為此期間麋鹿的近交系數為0.16—0.26,若假定烏邦寺初始麋鹿的近交系數為0,Foose等推算1977年麋鹿的平均近交系數為0.116。麋鹿重引進中國后,于長青[44]對中國麋鹿種群進行有效種群數量分析,結果顯示我國麋鹿近交現象極其明顯,平均近交系數已高于0.2,這與Sternicki等[47]利用國際物種信息系統(International Species Information System,ISIS)對世界各地動物園登記的1947—2002年間出生的2 042頭麋鹿進行研究的結果相符,該研究顯示麋鹿近交系數0.242 2—0.281 2,高于兄妹交配的近交系數。麋鹿作為高度近交的種群,在經歷了十九世紀末極其嚴重的瓶頸效應以及其后烏邦寺莊園對外輸出的多次間歇性瓶頸效應之后,遺傳多樣性較低。同時,受到環境條件的制約,使麋鹿的生長受到一定程度的制約,最終導致遺傳質量較低[2]。關于中國的麋鹿遺傳多樣性,學者們進行了一系列的研究。蛋白質電泳分析表明,現在麋鹿種群沒有發現明顯多態性,遺傳多樣性嚴重降低,近乎純合種群。
到了二十一世紀初期,中國學者通過線粒體、基因片段開展遺傳多樣性研究。Zeng等[48]利用線粒體DNA控制區及微衛星位點對來自中國的3個已發展穩定的麋鹿種群:北京麋鹿苑、江蘇大豐麋鹿國家級保護區、湖北石首麋鹿國家級保護區研究發現,3個種群共享1個線粒體單倍型,目前中國麋鹿在母系遺傳上非常單一,沒有發生分化;84對跨物種微衛星引物只篩選出5個微衛星位點,且檢測結果得出的平均期望雜合度為0.46—0.54,表明我國麋鹿種群的遺傳多樣性比較貧乏。吳海龍等[49]篩選出8個微衛星位點,構建了麋鹿血液cDNA文庫,以用于麋鹿MHC基因及表達基因的研究。張林源等采用25個微衛星位點對中國麋鹿進行群體檢測研究,結果顯示北京麋鹿生態實驗中心的麋鹿種群及其浙江臨安的麋鹿亞種群的遺傳多樣性較低[50]。孟浩等[37]通過分析麋鹿線粒體DNA 控制區序列發現,在所測得的麋鹿樣本序列中,同源性達99.81%,單倍型多樣度(Hd)與核苷酸多樣度(π)都較低,也表明了麋鹿群體多態性不高。匡葉葉[28]通過研究麋鹿個體間遺傳距離,發現北京麋鹿苑麋鹿群體具有極高的遺傳異質性,種群遺傳多樣性極低,為一個較為純合的種群。
目前,關于麋鹿遺傳多樣性的研究已經進入到基因組水平。Zhang等[51]通過第二代高通量基因組測序技術,首次構建了麋鹿基因組草圖,經與馬、牛、羊對比,發現麋鹿具有物種特有的69個基因家族、167個基因。Zhu等[52]利用種群基因組測序和比較基因組分析,對麋鹿遺傳多樣性進行了研究,結果顯示,與瀕危的朱鹮(Nipponianippon,建群個體數僅為7只)[53]、大熊貓(Ailuropodamelanoleuca,較高的遺傳多樣性)[54]以及北極熊(Ursusmaritimus)的基因組[55]相比,朱鹮的近交程度最高,麋鹿次之。并認為麋鹿并沒有存在很高的近交程度[52](Froh值越大,表示近交的程度越大)(圖3),與多年來麋鹿群體的幼體存活率的結果比較一致,推測中國麋鹿種群的77頭奠基者可能比朱鹮的7個奠基者具有更多遺傳多樣性。比較基因組分析顯示,麋鹿群體盡管經歷過嚴重的種群瓶頸,但是其仍存留較高的遺傳多樣性,推測近親繁殖歷史可能有助于清除有害的隱性等位基因。同時,檢測到一批受到正選擇的基因,涉及對高鹽食物的潛在適應、血壓調節、心血管發育、膽固醇調節、血糖控制和甲狀腺激素合成,推測這些適應性遺傳特征有助于緩解與高鹽飲食相關的不利影響。
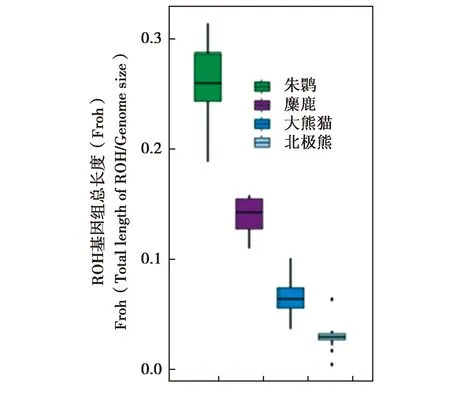
圖3a 麋鹿、朱鹮、大熊貓與北極熊種群ROH基因組長度比例Fig.3a Box plot of Froh for Père David’s deer,crested ibis,panda,and polar bear populations.Froh denotes the proportion of total ROH length
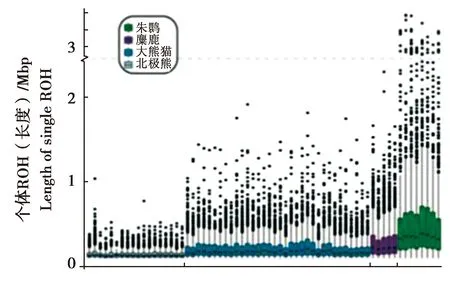
圖3b 麋鹿、朱鹮、大熊貓與北極熊個體ROH基因組長度比例(引自Zhu等,2018)[52]Fig.3b Box plot of length of ROH in each individual from Père David’s deer,crested ibis,panda,and polar bear
4 小結
麋鹿是中國重點保護的小種群代表動物之一,現存的麋鹿均為達氏種。2018年的研究結果,建立了麋鹿種之下的一個亞種——原達氏鹿[16],相比較與現生的麋鹿,新發現的這類麋鹿的特點是鹿角前枝比后枝更纖細短小,其前枝的分支點距離前枝與后枝的分支點較近。通過對麋鹿化石種分類的討論,將麋鹿屬分為4個種和8個亞種,對研究麋鹿屬在中國的演化提供了重要證據。
根據考古研究,鹿亞科在中新世早期從鹿科總進化枝分化出來,而鹿屬在上新世末期(Terminal Pliocene)至更新世初期起源并逐步分化。麋鹿出現于更新世早期(Early Pleistocene),與鹿屬其他物種的分歧時間大約在 270 萬年[26]。坡鹿是中國鹿屬中最原始的種,與鹿屬其他種的分歧時間在270萬—320萬年。水鹿與白唇鹿、梅花鹿和馬鹿的分歧時間在 200萬—240 萬年。梅花鹿與馬鹿的分歧時間在 140萬—160 萬年。大多數學者認為麋鹿與鹿屬的親緣關系近,尤其是與坡鹿的親緣關系最近。
麋鹿在經歷了十九世紀末極其嚴重的瓶頸效應以及其后烏邦寺莊園對外輸出的多次間歇性瓶頸效應之后,中性遺傳多樣性基本喪失,種群內個體之間的遺傳相似系數極高,理論上極度缺乏維持其可持續發展的生存潛力,麋鹿存在著近交衰退而走向滅絕的威脅。但是,從烏邦寺建群到現今遍布世界的多個小種群來看,由于中國政府的大力支持與保護,加上麋鹿對環境的潛在適應,麋鹿的數量一直在逐步增長,且未發現有明顯的生存力和繁殖力下降的情況,說明麋鹿群體復壯取得初步成功。
目前,嚴重的近交似乎對麋鹿的種群繁衍未產生明顯的不良影響。根據北京麋鹿苑的記載,近20年來幼體存活率一直維持在80%以上,成體體重以及雄性個體鹿角明顯增大,子代發生畸形和母獸難產現象出現的概率都非常低。理論上,近交系數每增加10%,種群繁殖能力將下降25%左右,一方面,近親繁殖似乎并沒有對麋鹿的繁殖力和生存力產生明顯的影響。有學者認為,現存麋鹿在建群初期,種群內部發生了積極的遺傳凈化作用,結合麋鹿種群迅速增長和繁殖過程中的有效競爭選擇機制等,導致種群內部的稀有有害等位基因被淘汰,從而使種群內優良基因得以保存,耐受近交效應的能力增強[4,52]。另一方面,遺傳多樣性水平的降低也可能使麋鹿抵抗隨機風險的能力隨之降低,易受到自然災害及疫病的嚴重威脅,湖北石首麋鹿保護區2008年受到雪災的影響,造成大批麋鹿死亡。2010年北京南海子麋鹿苑和湖北石首麋鹿國家級自然保護區由于受魏氏梭菌(Clostridiumwelchii)的影響,麋鹿種群猝死率分別為10.87%和23.35%[56]。麋鹿是高度特化的物種,如果不加以人為打擾,一旦面臨劇烈的條件改變,極可能由于缺乏自身的適應調節機制,遭受物種絕滅危險.這也是盡管全世界各地目前的麋鹿數量有了較大增長,但是并不足以將其從瀕危物種名單中刪除的原因[36]。
盡管麋鹿線粒體DNA多樣性研究顯示其具有極低的遺傳多樣性,但從其基因組水平多樣性顯示,中國麋鹿種群的77只奠基者比朱鹮的7只奠基者[57]具有更多遺傳多樣性[51-52]。考慮到朱鹮和麋鹿現存種群的復壯成果,建議進一步開展核基因水平和基因組水平多樣性方面的研究工作,以促進我國麋鹿種群的健康可持續繁衍。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
