基于高通量RNA轉錄組測序技術分析特應性皮炎患者皮損與非皮損組織差異表達基因
2019-11-28 02:11:36陳麗潔梁景耀張錫寶邵蕾潘清麗何素玲劉玉梅王建琴
中華皮膚科雜志 2019年10期
陳麗潔 梁景耀 張錫寶 邵蕾 潘清麗 何素玲 劉玉梅 王建琴
廣州醫科大學皮膚病研究所 廣州市皮膚病防治所 510095
特應性皮炎(atopic dermatitis,AD)是一種常見的慢性遺傳相關性炎癥性皮膚病,發病機制尚未完全明確。多認為AD 發病與遺傳易感性、環境因素以及免疫因素密切相關[1]。目前對AD分子表型的認識多基于基因芯片或者實時定量聚合酶鏈反應測定等方法,而基于新一代RNA測序(RNA-seq)的研究很少。自2001年他克莫司及吡美莫司外用制劑上市以來,暫未見其他可以廣泛使用的安全高效治療AD的藥物[2]。提高對AD分子表型的認識,有助于其新療法,特別是靶向治療的研發。RNA-seq能夠彌補表達譜芯片研究的不足,它不依賴于預設計,允許對整條基因組進行分析,更大范圍地準確檢測基因表達水平。轉錄組(transcriptome)指特定組織或細胞在某一功能狀態下所有轉錄本的總和,包括編碼RNA(mRNA)和非編碼RNA(ncRNA),它們能夠反映細胞的生長、發育、凋亡等一系列重要的生理過程。以Illumina 為基礎的轉錄組測序RNA-Seq,具有通量更高、運行時間更短、測序片段更長、分析成本更低等優點。本研究運用RNA-seq技術篩選AD 皮損及非皮損組織的差異表達基因,從功能基因組水平揭示AD 的發病機制,為AD 的分子診斷、藥物治療、預防和新型藥物研發提供新的契機。
對象與方法
一、對象
選取2016年7-10月于廣州市皮膚病防治所/廣州醫科大學皮膚病研究所門診就診的5 例漢族AD患者,男3例,女2例,年齡9 ~ 23歲。
納入標準:①符合Williams診斷標準;②SCORAD評分>25的中重度患者[3];③切取皮損組織前1周沒有局部外用藥物治療,2 周內未口服治療AD 的藥物。排除標準:患藥疹、高IgE綜合征、惡性嗜酸性粒細胞增多、皮膚感染、結締組織病及腫瘤性疾病等。選取腹部(2例)、背部(2例)或大腿處(1例)皮損及非皮損部位(在皮損邊緣取材,離皮損區約1.5 cm)皮膚組織,分別裝入有RNAlater 保存液(德國,Qiagen公司)的5 ml凍存管,-80 ℃冰箱保存,用于后續總RNA提取。
本研究經過廣州市皮膚病防治所/廣州醫科大學皮膚病研究所醫學倫理委員會批準(批件號:201611),取材前受試者或其監護人均簽署知情同意書。
二、方法
1.RNA 提取及轉錄組測序:按照Trizol(美國Invitrogen 公司)法提取總RNA,構建文庫,使用Agilent 2100 Bioanalyzer(RNA 6000 Nano Kit,美國Agilent 公司)檢測總 RNA 濃度、RIN 值、28S/18S 和片段大小。qRT-PCR 對文庫的有效濃度進行準確定量,以保證文庫質量。基于邊合成邊測序(sequencing by synthesis,SBS)技術,利用 BGISEQ-500(深圳華大基因)平臺進行高通量測序,得到100 bp的測序讀長。
2.測序數據過濾:將測序所得的原始數據(即raw reads)中低質量、接頭污染以及未知堿基N 含量過高的reads 過濾掉,得到有效reads(即clean reads),以保證結果的可靠性。
3.基因差異分析、功能注釋及生物學通路分析:使用DEseq2和PossionDis方法進行差異檢測分析,DEseq2方法基于負二項分布原理,本研究根據既往文獻[4-5]描述的方法進行差異表達基因檢測。對差異檢驗得到的P值做多重假設檢驗校正[6],通過控制錯誤發現率(false discovery rate,FDR),用于控制多重試驗中的Ⅰ型錯誤率[7]來決定P值的閾值。不同樣品之間表達水平變化倍數超過2 倍(|log2FC| ≥ 1)且FDR 校正P< 0.05 表示差異有統計學意義。根據差異基因檢測結果,使用R軟件中的pheatmap 函數進行層次聚類分析。利用基因本體 論(gene ontology,GO)數 據 庫(http://www.geneontology.org/)對差異表達基因進行功能注釋,采用 COG(cluster of orthologous groups of proteins)對差異表達基因進行分類統計,P≤0.01的功能視為顯著富集。利用KEGG(Kyoto Encyclopedia of Genes and Genomes)數據庫(http://www.genome.jp/kegg)對差異表達基因進行pathway 功能分析,P≤0.01的通路視為顯著富集。

表1 候選基因引物序列
4.qRT-PCR 驗證候選基因:為驗證AD 患者轉錄組測序結果的準確性,選取轉錄組測序結果中的5條基因,在NCBI上檢索目的基因CXCL1、KRT6A、IL36A、SERPINB4、PSAPL1 的 mRNA,用 Primer Premier5 設計引物,見表 1。另外選取 5 例 2018 年5-7月在廣州市皮膚病防治所/廣州醫科大學皮膚病研究所門診就診的中重度漢族AD 患者,排除和納入標準同前,男4例,女1例,年齡17 ~25歲。患者或其監護人簽署知情同意書后,每例取皮損、非皮損部位組織各1 份,分別提取總RNA,逆轉錄為cDNA,使用ChamQ SYBR qPCR Master Mix 試劑盒(南京諾唯贊生物科技有限公司),參照說明書進行qPCR 擴增,依據Real-time Quantitative PCR(美國ABI公司,StepOne Plus)系統說明書進行qRT-PCR。每份樣品重復檢測3次,采用2-△△Ct法進行相對表達量計算。
結 果
一、RNA-seq數據分析
通過RNA-seq分析,每份樣品平均獲得10.96 Gb數據,clean reads Q30 都在84.4%以上。共檢測到基因21 729 條,其中已知的基因為19 268 條,預測新基因為2 545 條。檢測出23 153 個新轉錄本,其中18 889 個屬于已知蛋白編碼基因的新的可變剪接亞型,2 545個屬于新的蛋白編碼基因的轉錄本,其余1 719個屬于長鏈非編碼RNA。
二、差異表達基因檢測
RNA-seq 分析顯示,AD 患者皮損與非皮損組織相比,78 條基因表達差異有統計學意義,其中67 條在皮損中高表達,11 條低表達,包括已知與AD 炎癥(CXCL1/2/8、IL6/IL1β、MMP1、SERPINB4、S100A2、GZMB、OASL、OSM)、屏障功能(KRT16、FABP5、CYP1A1)、角質形成細胞分化(IL-20)等相關的基因。見圖1、表2。在類聚分析熱圖中,皮損和非皮損表達的基因可以明顯的區分出來,見圖2,表明皮損和非皮損有各自特征的表達譜,有不同的病理生理學發病機制。
三、差異表達基因的GO分析
對這些差異表達基因進行GO 分析顯示,在生物過程分類中,數量最多的是參與細胞過程(53 條)的基因,第2、3 位分別是參與單組織過程(47條)和代謝過程(45條)的基因。在分子功能分類中,催化活性(46 個)和結合整合功能(27 個)的基因數量最多。在細胞組分分類中,細胞(43個)、細胞組分(43 個)、細胞膜(30 個)基因數位居前三。此外,34條基因參與刺激反應,27條參與多細胞生物過程,還有其他較多基因參與生物調控、定位和信號傳遞相關功能。見圖3。
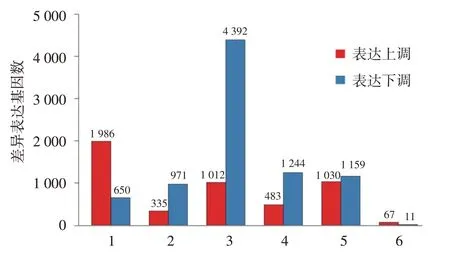
圖1 5 例特應性皮炎皮損與非皮損組織差異基因表達量比較 1 ~5:分別為病例1 ~5的皮損與非皮損比較;6:5例合計的皮損與非皮損比較

表2 特應性皮炎皮損與非皮損組織相比差異表達基因及差異倍數
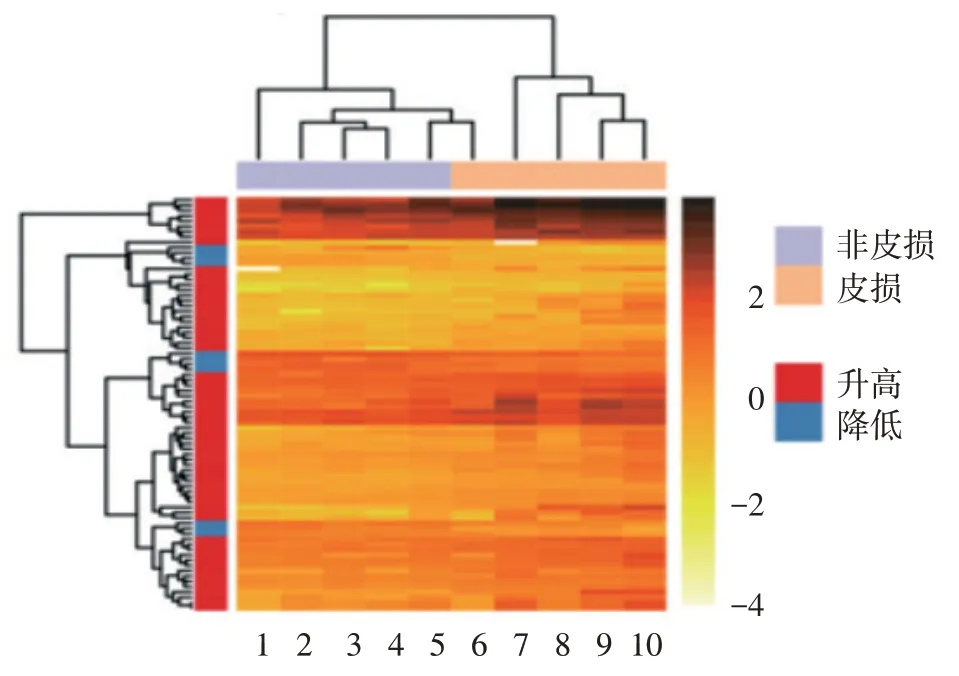
圖2 特應性皮炎皮損與非皮損基因表達聚類分析 1 ~5:分別為病例4、5、1、3、2 的非皮損;6 ~ 10:分別為病例1、2、4、5、3 的皮損
四、差異表達基因Pathway功能分析
78 條差異表達基因富集于132 條通路,其中13條通路顯著富集,包括白細胞介素17信號通路、NOD 樣受體信號通路、Toll 樣受體信號通路、腫瘤壞死因子信號通路、細胞因子受體、胞質DNA-傳感通路、沙門菌感染相互作用通路等。見圖4。
五、qRT-PCR驗證基因表達變化
結果顯示,CXCL1、KRT6A、IL36A、SERPINB4在皮損中的mRNA 表達明顯高于非皮損組織,而PSAPL1在皮損中的mRNA表達明顯低于非皮損組織,與轉錄組測序結果趨勢一致(圖5)。
討 論
目前AD轉錄組的研究主要利用傳統的基因芯片技術,只能檢測已知基因的轉錄本表達量,不能發現未知的新轉錄本,基因組覆蓋面較狹窄。RNASeq 測定差異基因的準確率顯著高于基因芯片[8]。了解患者皮損和非皮損差異表達基因對于探討AD的發病機制及尋找更精確的診斷方法及生物學標志物均具有重要意義。
我們首次運用新一代高通量測序技術對漢族中重度AD患者皮損和非皮損組織的轉錄組進行測序和功能分析。樣品比對顯示,基因組的平均比對率為 92.26%,clean reads Q20 都在95.34%以上,說明此次序列的質量滿足轉錄組分析的要求。共篩選得到21 729 條基因,其中19 268 條為已知基因,2 545條為預測的新基因。對皮損及非皮損組織進行差異表達基因分析,并對差異表達轉錄組基因進行功能分析,發現差異表達基因可能通過調節相關基因mRNA的表達進而影響AD炎癥的病理生理學改變。根據KEGG 數據庫分析發現,這些差異基因涉及132 個具體的代謝途徑分支,影響IL-17 信號通路、NOD樣受體信號通路、Toll樣受體信號通路、腫瘤壞死因子信號通路、細胞因子受體、胞質DNA-傳感通路等,為進一步大量挖掘AD 發病過程中的重要基因,開展AD 靶點治療及功能驗證等提供了基礎數據。
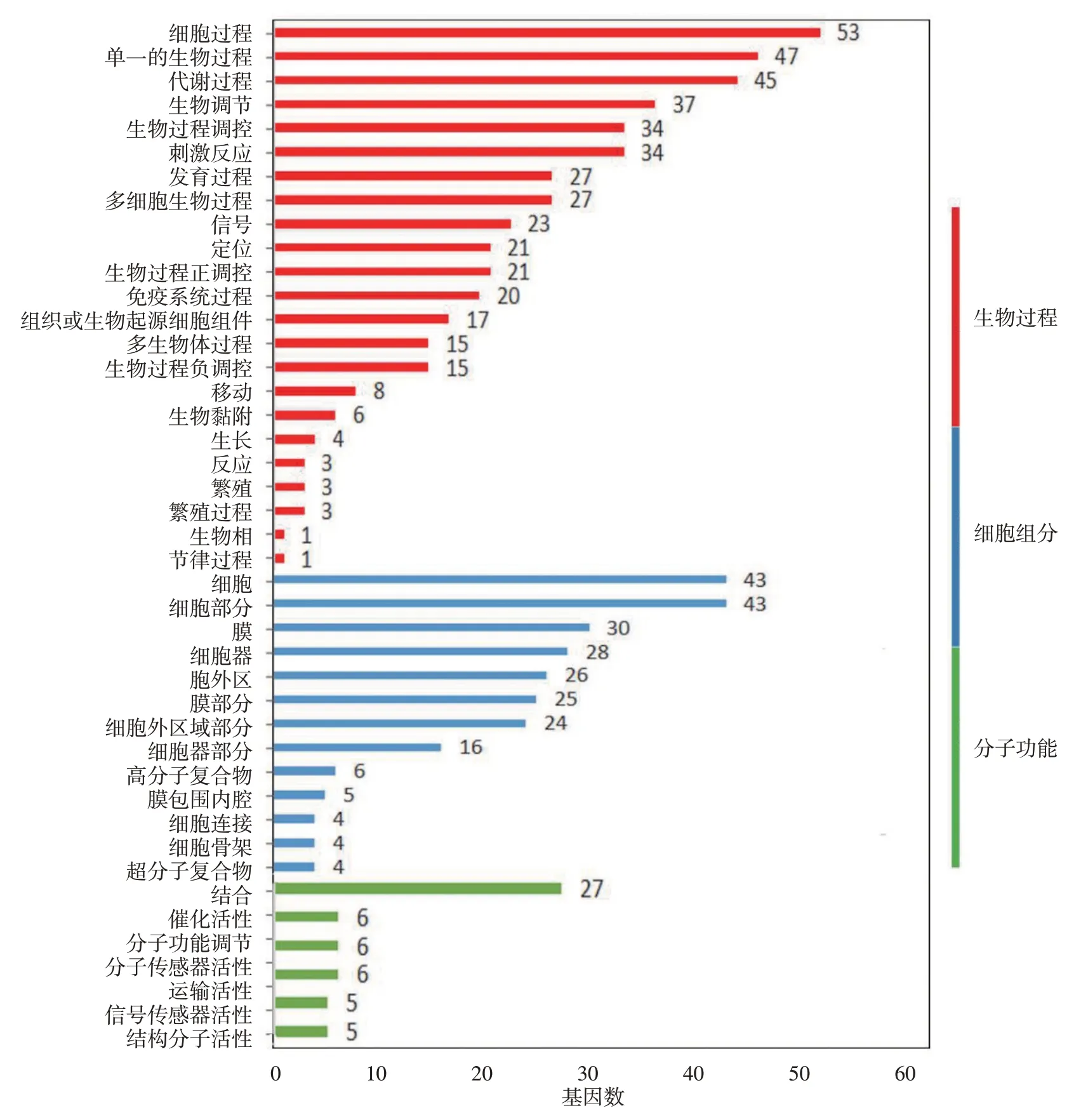
圖3 特應性皮炎皮損差異表達基因GO功能分類圖
本研究發現的78 條顯著差異表達基因,其中27 條目前已有報道和AD 相關,包括CXCL128、IL61β、MMP1、SERPINB4、S100A2 等。我們隨機選取5個候選基因用qRT-PCR進行驗證,結果與轉錄組測序趨勢一致,表明本研究結果可靠。在78條顯著差異表達基因中,上調倍數較高的6條基因與IL-17通路中中性粒細胞趨化、對外界病原的免疫、炎癥宿主的自身免疫防御調節相關。有研究顯示,IL-17 在亞裔AD 患者的發病機制中起非常關鍵的作用[9],本研究結果與上述研究相符。已發現銀屑病和AD 皮損中IL-17 過表達,而且有研究表明,IL-17C 作為炎癥的主要介質在這兩種疾病中具有致病性[10-11],因而提示抗IL-17 的生物制劑有可能用于治療亞裔AD。一項抗IL-17c 的單克隆抗體MOR106 的Ⅰ期臨床試驗結果令人鼓舞,研究組80%的AD 患者達到EASI50,相比之下,安慰劑組比例不到20%,而且MOR106還顯示出良好的安全性[12]。2018年Sanyal等[10]運用免疫組化、RNA-Seq等發現,非洲裔美國AD 患者具有Th2/Th22 偏向性,Th2 和Th22 標記物及IgE 與疾病嚴重程度顯著相關,但其Th1和Th17軸較歐洲裔美國AD患者衰減[13]。2015年一項運用RNA-seq對18例中重度美裔AD患者皮損和非皮損組織差異表達基因的研究發現,髓系細胞表達的觸發受體TREM-1(triggering receptor expressed on myeloid cells 1)通路和IL-36因子基因在患者皮損中高表達[14]。本研究中發現的差異表達基因與該研究有很多重合,但本研究顯示,漢族AD的差異表達基因具有以IL-17為主的模式特點。
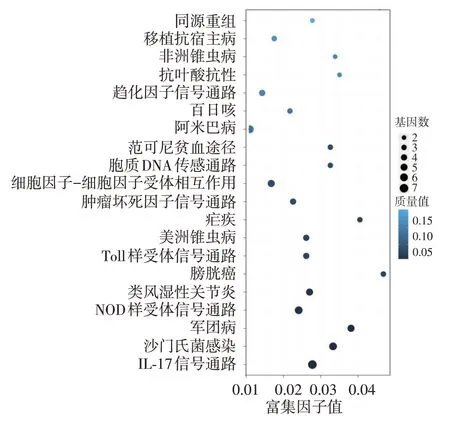
圖4 特應性皮炎皮損差異表達基因Pathway富集結果
本研究顯示,炎癥相關基因CXCL1、IL-1β、CXCL8、IL1RL1、OSM 等在皮損處顯著高表達。Jung 等[15]發現用異澤蘭黃素 eupatilin 治療 AD 小鼠,IL-1β 水平會相應降低。Khattri 等[16]用生物制劑ustekinumab 治療中重度成人AD,4 周后CXCL1水平降低。在哮喘患者痰細胞中CXCL8的表達水平和巨噬細胞的百分比呈負相關[17],在嚴重的哮喘患者中 OSM 主要來源是中性粒細胞[18]。IL-33 和IL1RL1 變異與巴西人群中哮喘和過敏性哮喘有關,而RA50、IL-33 和 IL1RL1 多態性與中國人群特應性哮喘有關[19]。
本研究顯示,屏障功能相關的SERPINB3/B4、KRT16 基因在漢族AD 患者皮損處顯著高表達。已經有研究證實,SERPINB3/B4和小鼠早期炎癥和屏障功能障礙相關,且用于構建小鼠AD 模型[20]。有學者匯總5 項獨立研究分析AD 差異表達基因[21],總共127個樣本,89條基因表達有顯著差異,功能注釋顯示這些基因參與免疫反應(如防御素、原蛋白)、角質形成細胞分化、表皮發育(如FLG、CORIN、AQP、LOR、KRT16)、炎 癥(如 IL-37、IL27RA、CCL18)和脂質代謝(如AKR1B10、FAD7、FAR2)。本研究發現,角質形成細胞分化相關基因IL-20 在皮損處顯著高表達。促炎細胞因子IL-20和IL-24 對FLG 表達有一定影響,并參與FLG 的終末分化[22]。
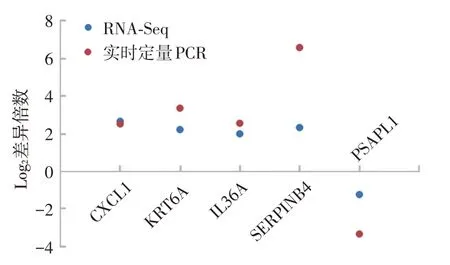
圖5 特應性皮炎(5例)皮損與非皮損組織差異表達基因的實時定量PCR 驗證 選取5 個轉錄組測序所得的差異表達基因CXCL1、KRT6A、IL36A、SERPINB4、PSAPL1 進行實時定量 PCR 驗證,結果與轉錄組測序顯示的表達趨勢一致
綜上,本研究通過新一代高通量轉錄組測序技術篩選出漢族中重度AD皮損和非皮損間差異表達基因,為進一步探索AD 的病理生理學機制和個性化治療提供了依據。
利益沖突所有作者均聲明不存在利益沖突
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
