遺傳因素與男性不育相關性研究進展
2019-12-26 01:29:06馬良宏
中國男科學雜志 2019年5期
關鍵詞:區域
馬 珂 田 稼 馬良宏
1.寧夏醫科大學臨床醫學院(寧夏銀川 750004);2.寧夏醫科大學總醫院寧夏人類精子庫;3.寧夏醫科大學總醫院泌尿外科;4.寧夏醫科大學生育力保持教育部重點實驗室
男性不育是由多種因素(男性精子形成、發育、成熟障礙或精子輸出受阻)導致的,育齡夫婦有正常性生活,且未采用任何避孕措施,婚后1年不能受孕的男性因素的總稱[1]。目前據WHO統計,世界約有15%育齡夫婦不能生育,男性因素至少占50%[2]。
一、目前男性不育致病因素研究進展
男性不育最常由精子發生障礙引起,臨床上稱為少精子癥或無精子癥。男性不育致病因素復雜(見圖1),目前已知的主要因素包括遺傳因素、抗精子抗體、感染、內分泌因素、疾病、鋅缺乏、環境、飲食和吸煙等[3]。但以上各種因素引起少弱精癥或無精子癥的機制尚未完全闡明。目前導致男性不育的機制主要有以下三種:(1)諸多研究表明這些因素與生精細胞的增殖與凋亡失衡有關。凋亡是細胞主動退化的一種機制,生精過程中細胞凋亡對維持正常的精子生成至關重要,該過程的失衡可能是導致少精子癥、非梗阻性無精子癥的重要機制之一;(2)一部分因素通過影響生精細胞或精子體內的能量代謝通路,使其體內能量代謝異常,進而引起弱精子癥而致男性不育;(3)一部分因素如睪丸炎癥等可以通過影響精子從形成到射出的輸出的通道阻塞而致梗阻性無精子癥導致男性不育。根據這些因素作用機制大致可分為精前性、精子性及精后性。除此之外男性不育患者中約有1/3的患者無明確病因,這部分患者稱為特發性男性不育癥者。
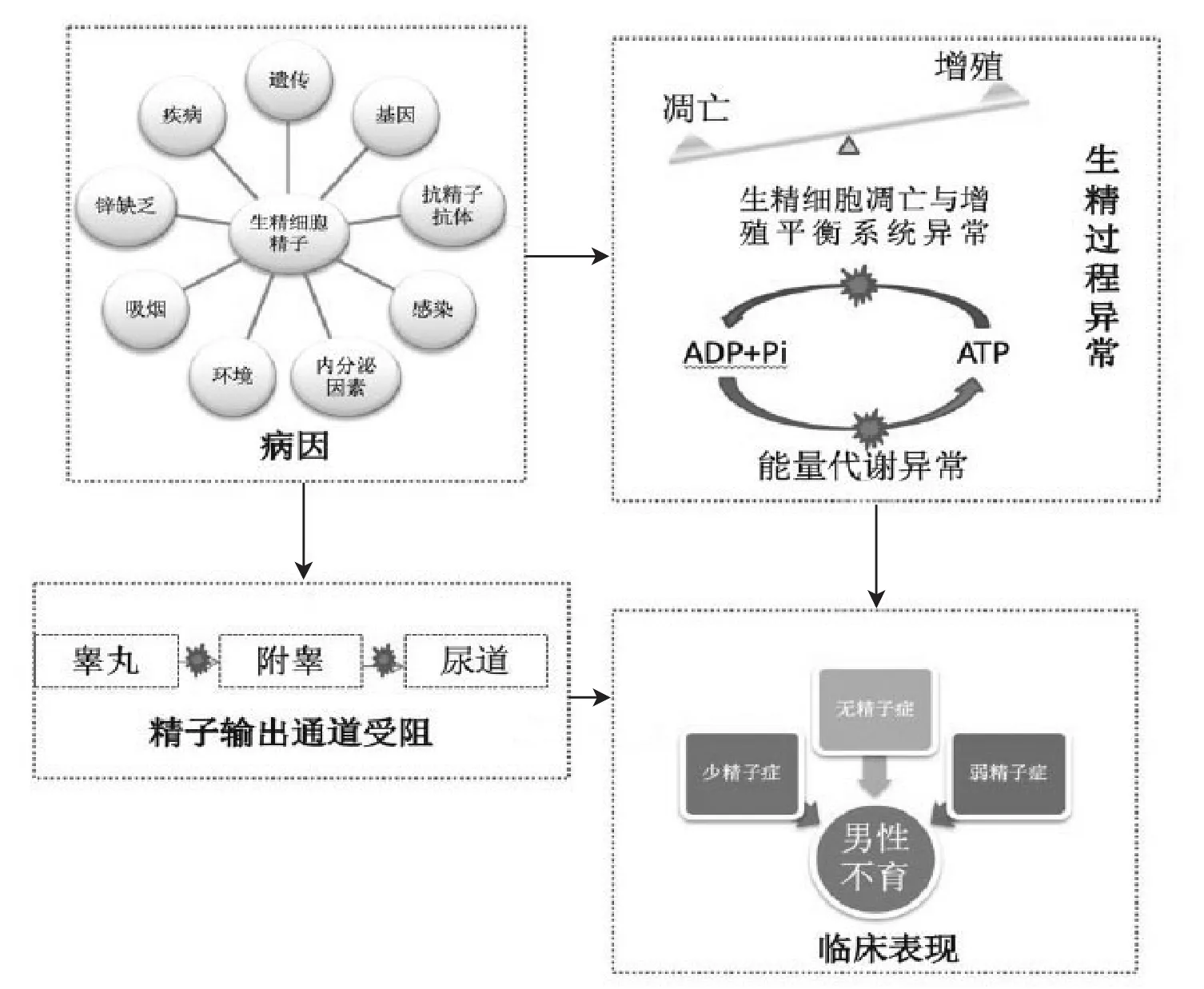
圖1 男性不育相關因素致病過程模式圖
二、遺傳因素與男性不育相關性研究進展
精子的發生是一個受到精細調節的眾多基因共同參與的復雜而有序的過程。研究表明,在少精子癥、弱精子癥及非梗阻性無精子癥發生的因素中,遺傳因素占到了大約30%,這其中包括基因突變和染色體異常[4]。無精子癥(或嚴重的少精子癥)最常見的遺傳因素是克氏綜合征(47,XXY)、染色體結構異常(例如易位)或Y染色體長臂上AZF區域的微缺失。染色體核型異常,與精子生成相關的基因缺失、突變或表達異常,都可能導致精子生成障礙,形成少精子癥、弱精子癥或無精子癥,最終導致男性不育。
(一)性染色體與男性不育
人類有23對染色體,正常男性性染色體核型為46,XY。目前研究發現人類23對染色體的結構、數量均與生殖發育密切相關。
X、Y染色體作為決定性別發育的重要染色體,染色體上有決定性腺發育成熟和生殖細胞形成的重要基因。X、Y染色體的數量、結構變化往往直接決定了個體性發育成熟以及生殖細胞、胚胎的發育與結局,因此目前X、Y染色體結構、數量變化及其基因仍是學術界研究男性不育的重點(圖2)。
人類Y染色體大約長60 Mb,有擬常染色體區(pseudoautosomal regions,PARs)和男性特異性區域(male-specific region of the Y chromosome,MSY)。擬常染色體區位于Y染色體兩端,在減數分裂過程中可與X染色體發生重組,男性特異性區域約占Y染色體總長的95%,由異染色質序列以及3類常染色質序列拼接而成[5]。
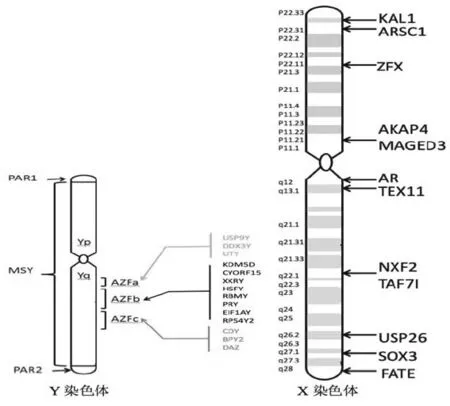
圖2 性染色體與男性不育基因
Li等[6]通過對我國東北4659位特發性不育患者的研究,發現其中412位患者存在染色體異常,其中性染色體異常314(6.74%)例;常染色體異常98(2.10%)例,染色體數目和結構異常分別占到259(62.86%)例和153(37.14%)例,其中常見的性染色體核型異常為:46,XX、47,XYY和45,X/46,XY。
1.Y染色體異常與男性不育 男性不育患者中,有10%~15%的非梗阻性無精子癥和5%~10%的嚴重少精子癥由Y染色體微缺失引起[7],Y染色體長臂(Yq11)常染色質區存在精子發生相關基因,稱為無精子因子(azoospermia factor,AZF)。該因子通常與少精子癥有關,尤其是無精子癥,是男性不育的第二大常見遺傳病因。AZF分為AZFa、AZFb和AZFc 3個區域,現已發現這3個區域內至少有15個與精子發生相關的基因。這些區域含有基因和轉錄單位,其中大多數呈現特定的睪丸表達,也是目前研究發現與嚴重少精子癥、非梗阻性無精子癥發生最常見的染色體微缺失區域[8-10]。
除了已報道的3個區域外,在AZFb和AZFc區之間重復區域也存在STS片段的丟失,有學者將這個區域命名為AZFd區,主要包括sY145位點,AZFd是否存在,目前仍有爭議,最新修訂的2014年版歐洲男科指南將sY145單獨列出[11]。AZF缺失的類型和臨床表型豐富多樣。中國人無精子癥和嚴重少精子癥患者Y染色體微缺失率約為6.59%,其中AZFc區和AZFb+c+sY145區缺失最為常見[12]。
Y染色體微缺失的相對高發生率與其結構相關。它富含高度重復的序列(包括直接重復序列,反向重復序列和回文序列)和高度多態性,這使得長臂特別容易受到通過具有升高序列同源性的區域之間的染色體內同源重組而出現的缺失[13]。
無精子缺失基因(DAZ),位于AZFc區域,是第一個被確定為導致人類無精子癥的基因,在睪丸特異性表達[14]。它編碼366個氨基酸的蛋白質,含有具有RNA結合功能的RNP/RRM結構域和7個串聯重復的14個殘基[15]。
RNA結合基序基因(RBMY),位于AZFb區域,編碼RNA結合蛋白(同DAZ),在胎兒、青少年和成人睪丸生殖細胞中特異性表達,但在體細胞如Sertoli細胞中不表達。RBMY缺失與生殖細胞分化有關,主要影響早期減數分裂[16]。
USP9Y基因,位于AZFa區域,該基因缺失會導致移碼和缺乏外顯子7,導致蛋白質水平降低約90%[17]。
最近發現了AZF區一些較小的缺失,例如AZFc區域的拷貝數變異(CNVs)(主要是gr/gr缺失[18]),可能與生精障礙有關,盡管這種情況也可能在正常精子男性中發現[19];b2/b3缺失與精子發生障礙也有關聯[20]。Y染色體微缺失還與試管嬰兒胚胎質量差、神經精神類疾病、腫瘤存在相關性[21,22]。
2.X染色體異常與男性不育的關系 到目前為止已經發現4種主要的X染色體畸變:(1)在克氏綜合征中發現的X染色體的非整倍性 (47,XXY或其他X染色體的鑲嵌性)。(2)涉及X染色體的易位,例如非綜合性46,XX性睪丸性發育障礙(XX-男性綜合征)或X-常染色體易位[23]。 (3)CNVs影響 X 染色體。 (4)點突變破壞X染色體基因[24]。
克氏綜合征:是男性不育最常見的核型異常。不育男性中的KS患病率非常高:嚴重少精子癥患者高達5%,無精子癥患者高達10%。在80%的病例中,KS的核型是47,XXY。剩余患者呈現更多的非整倍體,例如48,XXXY或49,XXXXY,其特征是更嚴重的臨床形式。過多的性染色體源于精子發生和卵子發生過程中,父母配子的錯誤分離,主要是不正確的減數分裂造成的[25]。
減數分裂在產生生物多樣性和支持物種生存方面發揮著作用,參與染色體交換和DNA重組的許多減數分裂蛋白在進化中高度保守。減數分裂的改變會破壞配子體發生,導致部分或完全的不育。位于X染色體上的TEX11部分缺失,可以導致減數分裂停滯,這意味著前體細胞不能進行適當的減數分裂。這是與X染色體上的基因有關的男性不育的第一個主要遺傳原因[26]。
雄激素受體(androgen receptor,AR)基因位于X染色體長臂上(Xq11-12)[27],由8個外顯子組成,編碼4個功能性區域:N-末端轉錄區 (外顯子1)、DNA結合區(外顯子2和3)、鉸鏈區、配體結合區(外顯子4-8),編碼919個氨基酸殘基的蛋白質[28]。突變會引起雄激素不敏感綜合征(AIS)并導致少精子癥或無精子癥[29]。AR基因外顯子1是目前研究的熱點,外顯子1突變會導致其轉錄激活效能降低,減弱AR對靶基因的表達調節,使精子發生過程改變[30]。約2%~3%的無精子或嚴重少精子患者是由AR基因突變引起[31]。
ANOS1(以前稱為KAL1)位于X染色體短臂上(Xq22,31),在促性腺激素釋放激素(GnRH)產生神經元的遷移中起重要作用,突變會引起Kallmann綜合征導致低促性腺性功能減退癥和精子發生的改變[32]。
RHOX包含3個基因:RHOXF1,RHOXF2和RHOXF2B,它們在人類卵母細胞和雄性生殖細胞中選擇性表達[33,34]。RHOXF2/2B突變顯著損害了調節HSP70家族轉錄因子和伴侶等下游基因的能力[35,36]。此外,USP26的變異也與男性不育相關,但有些結果相互矛盾,仍需進一步研究[37]。
(二)常染色體與男性不育
迄今為止,研究發現與少弱精子癥、無精子癥發生相關的基因不僅存在于性染色體上,在常染色體中也大量存在。
羅伯遜易位:發生于近端著絲粒染色體的一種易位形式。兩個近端著絲粒染色體于著絲粒或附近斷裂后,導致兩條中心染色體長臂融合 (D組染色體:13,14,15 和 G 組染色體:21,22eY)[38],形成一條由長臂構成的衍生染色體。通常為雙著絲粒,丟失了相應的短臂。在載體表型中無明顯變化,但可能導致睪丸體積和睪酮水平降低,影響精子發生或導致胎兒死亡[39]。
CFTR基因:位于 7號染色體上(7q31.1-31.2),純合子形式引起囊性纖維化,是高加索人群中最常見和最嚴重的常染色體隱性疾病之一。CFTR表達的突變會引起男性先天性雙側輸精管缺失(CBAVD),導致阻塞性無精子癥[40]。
INSL3-RXFP2基因:INSL3的編碼基因位于19號染色體上,由兩個帶內含子的外顯子組成。在患有隱睪癥的男性中,與這兩個基因突變相關的患病率約為4%~5%[41]。
SYCP3基因:位于12號染色體上(12q23),在睪丸中特異性表達。它編碼236個氨基酸的蛋白質,具有兩個卷曲螺旋結構域,是一種與生殖細胞減數分裂過程中涉及的突觸相關的DNA結合蛋白。突變引起減數分裂異常,造成卷曲螺旋結構域腺苷堿基的缺失,導致移碼突變并出現早期終止密碼子,導致結構域不完整。對突變進行功能分析,結果顯示變短的SYCP3幾乎沒有功能性蛋白質-蛋白質相互作用[42]。SYCP3突變通過男性減數分裂停滯和女性復發性妊娠引起無精子癥[43]。
此外,KLHL10突變會損害少精子癥患者的同源二聚化[44];AURKC的純合子突變產生大頭多倍體精子[45],均與男性不育相關。
(三)線粒體DNA異常與男性不育
精子活力是男性生育力的主要決定因素之一,精子中的線粒體是提供精子活力必需的細胞器。人類mtDNA僅由沒有內含子的外顯子組成。由于高轉換率,沒有校對或有效的DNA修復系統,缺乏組蛋白保護或DNA結合蛋白,mtDNA突變率比核DNA高10~100倍。線粒體DNA突變可能會減少ATP產生從而降低男性生育能力。線粒體DNA拷貝數也是線粒體遺傳變異和病理學的重要因素[46]。
研究表明,精液參數改變的患者mtDNA拷貝數顯著增加,mtDNA完整性顯著降低,活性氧(ROS)產生增加[47]。mtDNA的大規模缺失可能導致不完整的呼吸鏈的產生,這可能導致男性生育能力和精子活力的下降[48]。
最近有研究顯示mtDNA 4,977-bp缺失與精索靜脈曲張引起的不育相關,但有必要進行進一步研究[49]。
三、結語
男性不育的遺傳學研究是一個快速發展的領域。尋找與男性不育致病相關的染色體、基因變化越來越受到研究界的重視。過去幾年的遺傳學關聯研究,基因突變篩查,動物模型和基礎研究清楚地表明生精障礙和男性不育的高發病率。相信隨著研究的不斷深入,將會有更多的染色體、基因異常在男性不育致病方面的作用與機制被發現。這也將為進一步闡明男性不育癥發病的遺傳因素提供重要的理論依據,同時也為男性不育癥的基因靶向治療提供思路。
猜你喜歡
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
科學(2020年5期)2020-11-26 08:19:22
軟件(2020年3期)2020-04-20 01:45:18
商周刊(2018年15期)2018-07-27 01:41:20
敦煌學輯刊(2018年1期)2018-07-09 05:46:42
北京教育·普教版(2017年1期)2017-02-05 13:26:23
新疆農墾科技(2016年2期)2016-08-21 13:50:16
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
新疆財經大學學報(2015年3期)2015-12-10 03:49:15
