酸性礦山廢水對沉積物真核微生物群落的影響
2019-12-26 04:32:48張曉輝劉紅玲
中國環境科學 2019年12期
劉 帆,張曉輝,唐 宋,王 茂,劉紅玲*
酸性礦山廢水對沉積物真核微生物群落的影響
劉 帆1,張曉輝1,唐 宋2,王 茂3,劉紅玲1*
(1.南京大學環境學院,污染控制與資源化研究國家重點實驗室,江蘇南京 210023;2.中國疾病預防控制中心環境與健康相關產品安全所,北京 100021;3.中山大學公共衛生學院,廣東 廣州 510080)
對受酸性礦山廢水污染的橫石河上游到下游水體和沉積物中理化參數以及沉積物中真核微生物的多樣性進行系統調查,使用斯皮爾曼相關性分析和典范對應分析(CCA)甄別真核微生物群落的主要環境脅迫因子.結果表明:橫石河中真核微生物優勢種群為真菌門(Fungi)(4.51%~86.69%)、綠藻門(Chlorophyta)(0.61%~77.36%)和纖毛蟲門(Ciliophora)(0.81%~34.91%).沉積物中真核微生物群落Alpha多樣性隨酸性礦山廢水污染梯度降低而逐漸升高.與原核微生物群落不同,酸性礦山廢水環境沉積物中真核微生物群落結構的變化主要受硫酸根和電導率的影響.
酸性礦山廢水;真核微生物;硫酸根;電導率;下一代測序
酸性礦山廢水(AMD)是含硫的礦物在氧氣和水存在情況下,經風化、淋溶和微生物共同作用而形成[1].由于處理工藝缺失,處理費用昂貴,大部分酸性礦山廢水未經處理便直接排放到自然環境中[1].酸性礦山廢水具有pH值低,硫酸鹽、電導率和重金屬濃度高的特點[2], 對大多數生命來說都是難以生存的極端環境[3-5].然而,這種有極端環境脅迫的獨特生境卻孕育著大量嗜酸、耐酸、耐重金屬的微生物.由于環境脅迫較為明確,物種豐度相對較低,生物學與地球化學過程關聯緊密,AMD環境被認為是基于基因組定量研究微生物生態和群落功能的理想“龕位”[6-7].另外,由于AMD污染,環境中微生物豐度也較低,這種復雜性較低的系統為研究較小尺度內相關微生物基因異質性以及為推斷復雜微生物群落進化過程提供了一個理想的條件[7].
隨著第二代測序技術的發展,對這種酸性極端環境中微生物群落多樣性以及群落功能和進化的了解越來越清晰.以往的研究往往集中在原核微生物群落,以致對真核微生物群落的了解較為薄弱[4,8-10].然而,真核微生物群落在酸性礦山廢水的形成、沉積物生態系統重金屬耐受、物質循環和能量流動等方面的作用不容忽視[11].
大寶山礦區位于廣東省韶關市曲江區境內,自19世紀70年代以來,采礦和冶煉活動產生的大量礦渣和酸性礦山廢水排入到下游的橫石河中,而橫石河是當地居民農業灌溉的主要水源之一.因此,橫石河的嚴重污染不僅給當地生態環境造成了極大的風險,還威脅到當地居民的健康[12].本研究選取廣東省大寶山礦區污染的橫石河作為研究點位,從上游到下游采樣,覆蓋不同的礦山廢水污染梯度,從極度污染區域到輕微污染區域.系統調查不同污染梯度中真核微生物群落多樣性的差異,并結合化學分析方法和生物信息學方法對引起真核微生物群落結構和多樣性變化的主要脅迫因子進行甄別.本研究拓展了對酸性礦山廢水污染環境中真核微生物群落多樣性的了解,為酸性礦山廢水污染生物修復提供了理論參考.
1 材料和方法
1.1 樣品采集
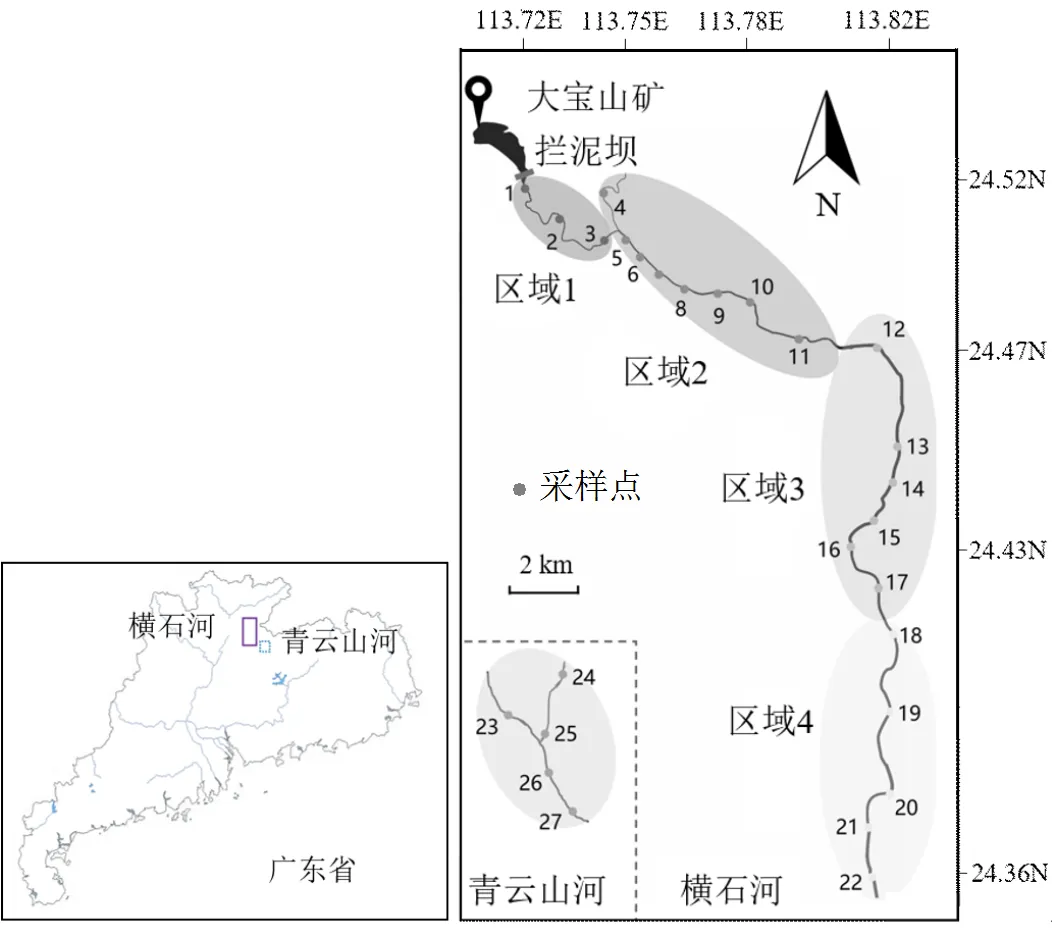
圖1 廣東省大寶山橫石河和對照河流采樣點位
選取大寶山礦區橫石河流域(約25km)作為研究區域.在2014年10月,沿橫石河從主污染源(大寶山礦區)到下游輕微污染區域設置采樣點,現場測定水樣pH值,采集500mL水樣4℃保存待分析.抓斗式采泥器(HYDRO-BIOS Apparatebau GmbH, Kiel- Holtenau, Germany)采集22個(1~22號)表面沉積物(河流底部0~5cm,約500g)樣品.根據距主污染源的距離和酸性礦山廢水污染梯度將橫石河分為4個污染區域(圖1):區域1-極端污染區域(1~3號樣品);區域2-重度污染區域(4~11號樣品);區域3-中度污染區域(12~17號樣品);區域4-輕度污染區域(18~22號樣品).此外,采集周邊未受采礦活動影響的河流中(青云山河)水體和沉積物樣品為無AMD污染對照(23~27號)樣品.一周內完成樣品凍干、磨細、分裝后保存于-80℃冰箱用于后續實驗.
1.2 理化參數測定
取0.5g干燥的沉積物于10mL 1mol/L的偏磷酸鈉((NaPO3)6)中分散24h后,使用Malvern Mastersizer 2000粒度儀(Malvern Instruments GmbH, Rigipsstr. Herrenberg, Germany)測定沉積物的粒度.剩余沉積物干樣經球磨儀磨碎過200目的尼龍篩,分別測定過氧化氫酶活力、pH值和電導率 (EC)、總有機碳(TOC)含量[13-15].取適量土壤于純水中震蕩提取,然后用Thermo ICS-5000離子色譜(Thermo Fisher Scientific Inc, Waltham, MA, USA)測定硫酸根(SO42-)含量.沉積物中重金屬含量篩查,將0.2g底泥樣品加入到12mL反王水(硝酸、鹽酸體積比為3:1)中,使用MILESTONE ETHOS UP (Milestone Office, Shelton, CT, USA)微波消解儀消解,趕酸稀釋后使用電感耦合等離子體發射光譜(ICP-OES) (PerkinElmer Office, Akron, OH, United States)篩查金屬含量(As、Ca、Cd、Co、Cr、Cu、K、Mg、Mn、Mo、Ni、Pb、Sr、V和Zn).然后對橫石河沉積物中顯著高于對照河流中的金屬(As、Cd、Cu、Pb和Zn)使用BCR連續提取法[16]進行重金屬4種形態分析,并測定水樣中這5種重金屬含量(ICP-MS (PerkinElmer Office, Akron, OH, United States)).
1.3 沉積物eDNA提取
使用土壤基因組DNA提取試劑盒,根據試劑盒建議的方法(基于MO BIO Powersoil DNA Isolation Kit)提取沉積物中的總DNA.使用Nanodrop (Nanodrop 2000, ThermoFisher, USA)檢測所提取DNA的濃度和純度,如果濃度小于20ng/μL,使用乙醇進行沉淀濃縮.
1.4 18S rDNA PCR擴增
用于18S rRNA基因V9區域的引物序列為: 1380F(5’-CCCTGCCHTTTGTACACAC-3’)和1510R (5’-CCTTCYGCAGGTTCACCTAC-3’)[17].上游引物5’端連接有Golay Barcode sequence用于區別樣品[18].PCR擴增條件為:退火溫度98℃反應30s,以下擴增步驟進行20個循環:變性(98℃,5s),退火(62℃, 30s),延伸(72℃,15s);循環結束后末次延伸條件為72℃反應7min.每次進行PCR反應時需設置對照組:陽性對照,陰性對照,無模板陰性對照.PCR反應結束后,1.5%瓊脂糖凝膠電泳檢驗PCR產物的大小,PCR產物經純化后以等物質的量混合組成測序文庫立即用于下游測序.
1.5 Ion Torrent測序和測序數據分析
測序工作以Ion Torrent Proton測序儀器相關試劑用戶手冊為依據.測序數據已上傳到NCBI Sequence Read Archive (SRA)數據庫(Accession SRP094407).生物信息學分析系統環境為Bio-Linux 8[19],測序數據處理的主要程序為QIIME[20]和USEARCH V9.0[21].通過RDP classifier[22]搜尋Protist Ribosomal Reference database (PR2)[23]進行比對注釋.所得高質量結果用于后續分析.
1.6 統計分析
所有統計分析主要是基于R語言(V3.6.1),對理化參數在橫石河不同區域以及橫石河和對照河流之間的差異都進行Kruskal-Wallis秩和檢驗,<0.05視為有顯著差異.使用斯皮爾曼相關性分析來研究Alpha多樣性與理化變量的相關性.使用典范對應分析(CCA)來確定重要的理化參數對微生物群落的影響,理化參數選擇依據CCA計算過程中VIF (variance inflation factors)值來確定.
2 結果
2.1 理化參數

表1 橫石河主要理化參數
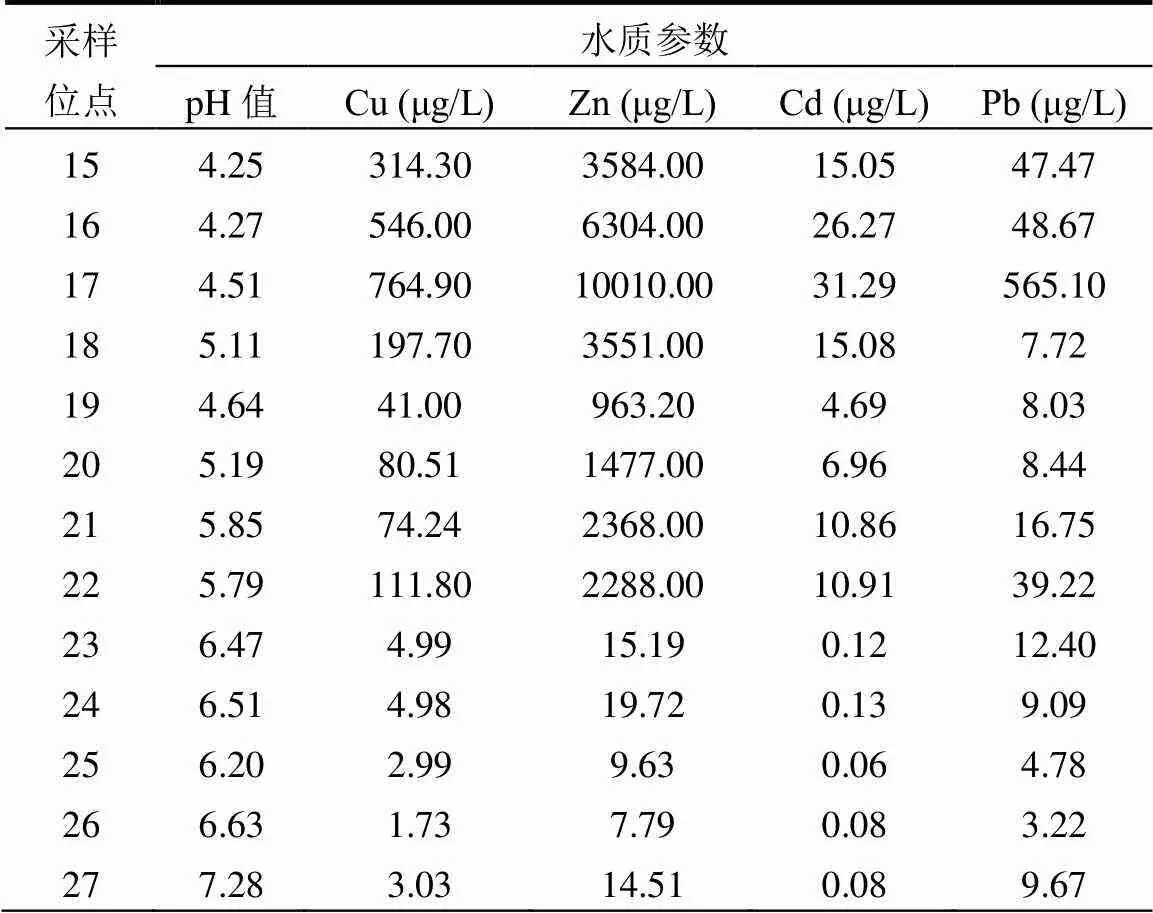
續表1
橫石河理化參數如表1所示,分析結果顯示,上下游水體和沉積物理化參數差異巨大,大部分參數的數值有沿橫石河顯著下降的趨勢.橫石河極端污染區域酸度極高,水體和沉積物中的pH值分別為2.98和2.64,到下游pH值逐漸回升,達到5.79和7.60.水體中所有重金屬和沉積物中大部分重金屬濃度有從上游到下游顯著下降的趨勢,但是,沉積物中Cu的濃度在橫石河下游反而上升.沉積物中電導率和硫酸根濃度也有沿橫石河顯著降低的趨勢.
2.2 測序結果
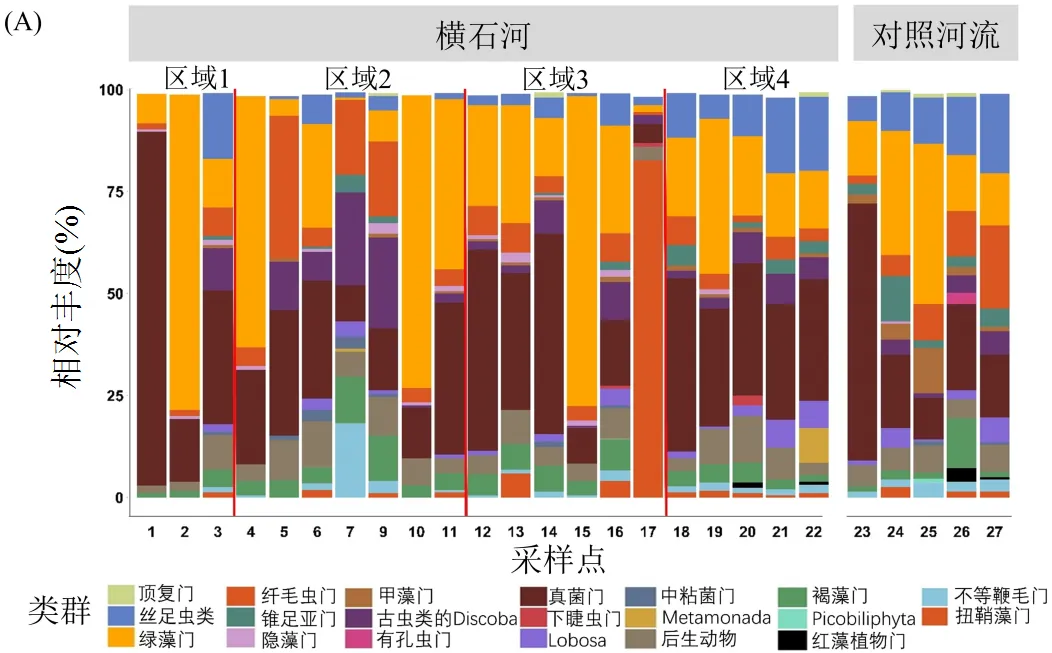
(A) 各采樣點真核微生物“門”分類水平相對豐度(B) 橫石河不同污染區域獨有和共有OTU數目.韋恩圖相對豐度小于0.5%的門類未展示
在橫石河和對照河流沉積物樣品中共獲得392596條reads,去除低質量樣品(8號樣品),根據97%的相似度可以分為3436個OTU,其中,在橫石河4個污染區域和對照河流都存在的OTU有756個.通過物種注釋結果,優勢種群為:后鞭毛生物類(Opisthokonta)的真菌門(Fungi)(4.51%~ 86.69%)、原始色素體生物的(Archaeplastida)的綠藻門(Chlorophyta)(0.61%~77.36%)、囊泡蟲類(Alveolata)的纖毛蟲門(Ciliophora)(0.81%~34.91%).其余豐度較高的門類還有古蟲類(Excavata)的Discoba(0.51%~22.67%)、有孔蟲類的(Rhizaria)絲足蟲類(Cercozoa)(0.65%~19.50%)、原始色素體生物的(Archaeplastida)的扭鞘藻門(Streptophyta) (0.56%~ 82.64%)、后鞭毛生物類(Opisthokonta)的Metazoa (1.82%~11.45%)、茸鞭生物界(Stramenopiles)的褐藻門(Ochrophyta)(1.11%~12.23%)和不等鞭毛門(Stramenopiles-X)(0.60%~18.26%)、變形蟲門(Amoebozoa)的錐足亞門(Conosa)(0.56%~11.08%) (圖2).
2.3 Alpha多樣性
Alpha多樣性指數可以反映特定環境中微生物群落豐富度和均勻度,是根據OTU數目評估生態系統多樣性的綜合性指標.一般來說,環境越惡劣,生態系統物種多樣性就越低[24],因此,可以根據群落的Alpha多樣性大小評估生態系統所承受的環境壓力大小.整體來看,橫石河沉積物種真核微生物群落的Alpha多樣性(Chao1指數和Shannon指數)(圖3(A)和(B))從上游到下游呈顯著上升的趨勢,并且橫石河內4個污染區域Alpha多樣性指數存在顯著差異.所以,橫石河上游到下游,隨著污染梯度下降,酸性礦山廢水污染對環境的脅迫水平逐漸降低.使用秩和檢驗對橫石河不同污染區域Alpha多樣性差異性進行檢驗(值<0.05*、<0.01**、<0.001***)
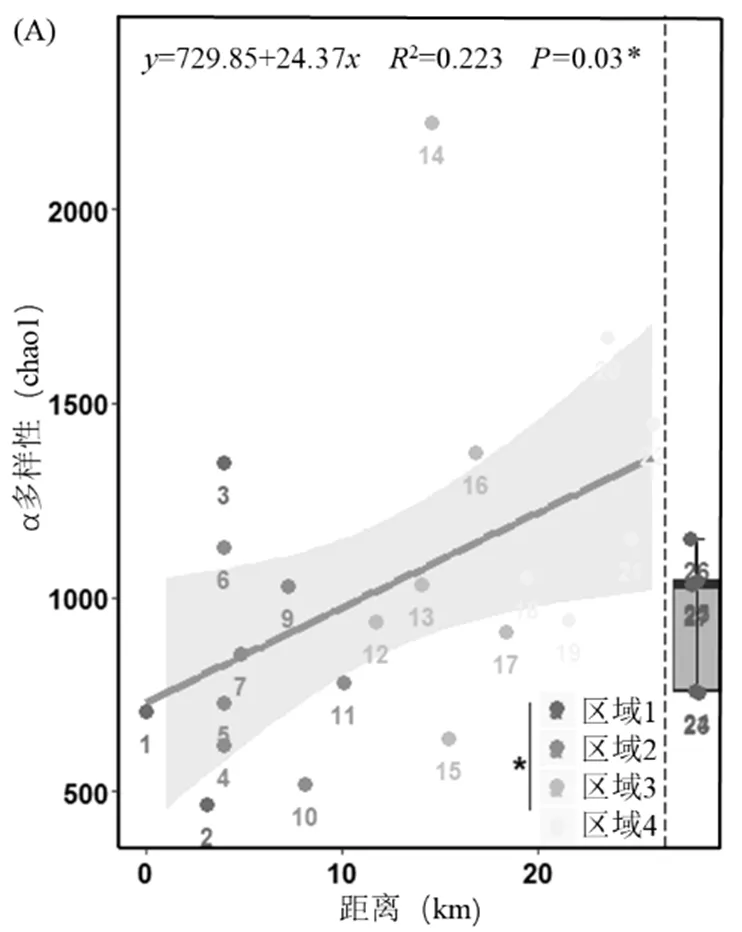
(A),(B) Alpha多樣性.;(C) Beta多樣性
2.4 Beta多樣性
根據各采樣點真核微生物群落Bray-Curtis相異性,使用非度量多元尺度分析(non-metric multidimensional scaling, NMDS)方法評估橫石河和對照河流沉積物樣本中微生物群落結構的差異性,并進一步使用非參數多元方差分析(PERMANOVA).結果顯示,橫石河不同污染區域(區域1、區域2、區域3和區域4)和對照河流沉積物中真核微生物群落結構存在顯著差異(=0.0214*);如果將橫石河當做整體來看,其沉積物中真核微生物群落結構和對照河流相比存在顯著差異(=0.0028**)(圖3(C)).
2.5 物種網絡分析

圖4 真核微生物“屬”分類水平相對豐度(相對豐度大于0.5%)和Alpha多樣性與橫石河理化參數相關性關系矩陣圖
右側數值為斯皮爾曼相關系數
使用Gephi v0.9.1和FruchtermanReingold放置算法的交互平臺[25],對橫石河真核微生物的關系進行網絡可視化和模塊檢測.在“科(family)”一級挑選橫石河沉積物中相對豐度大于0.5%,具有顯著共現關系(斯皮爾曼相關系數||>0.6且<0.01)的真核微生物進行共現網絡關系分析.結果發現橫石河中核心真核微生物有:傘菌綱下屬科(Agaricomycets)、異葉足綱下屬科(Heterolobosea_XX)、毛霉菌亞門下屬科(Mucoromycotina_X)、棘阿米巴科(Acanthamoebidae)、三足蟲科(Trinematidae)、Allapsidae沙棘科(Sandonidae)、Cercomonadidae、Schizoplasmodiids、卵菌門下屬科(Oomycota_XX)、Glissomonadida_X、多枝藻目下屬科(Chaetophorales)、小球藻科(Chlorellales_X)、衣藻(CW-Chlamydomonad)、Vermamoebidae、Neobodonid、鉤端螺旋體科(Leptophryidae)、擬阿米巴科(Paramoebidae)、顎足綱下屬科(Maxillopoda)、環藻目下屬科(Sphaeropleales)等.
2.6 真核微生物群落與理化參數的關系
橫石河中不同污染區域水體和沉積物中的理化參數,沉積物中真核微生物群落結構都顯著不同.為了了解環境因子對真核微生物群落的影響,首先將所測得的環境理化參數與在“屬”分類水平上相對豐度大于5%的真核微生物以及Alpha多樣性進行斯皮爾曼相關性分析.隨后、通過CCA分析影響微生物群落結構的主要環境理化因子.相關性分析結果顯示,與物種相對豐度和Alpha多樣性相關性較強的環境理化因子主要有:水體pH值(Aq. pH)、水體中銅(Aq. Cu)、鋅(Aq. Zn)、鎘(Aq. Cd)、鉛(Aq. Pb);沉積物pH值(Sd.pH)、沉積物中電導率(Sd.EC)、硫酸根(Sd.Sulfate)、三價鐵離子(Sd. Ferric)、總鐵(Sd.Total.Fe)、可提取態銅(Cu.Exchangeable)、可提取態鋅(Zn.Exchangeable)、可提取態鎘(Cd.Exchangeable)和還原態鎘(Cd.Reducible)(圖4).CCA結果顯示,對橫石河中真核微生物群落影響最大的環境理化因子是沉積物中硫酸根離子(Sd.Sulfate)和電導率(Sd.EC);其次則是水體的pH值(Aq.pH),水體中鉛濃度(Aq.Pb)、沉積物中三價鐵的濃度(Sd.Ferric)和沉積物中鉛(Sd.Pb)的濃度,隨后水體中鋅(Aq.Zn)的濃度、沉積物中二價鐵(Sd.Ferrous)的濃度、沉積物中總有機碳的含量(Sd.TOC)等環境因子也對真核生物的群落結構變化有著顯著的影響(圖5).
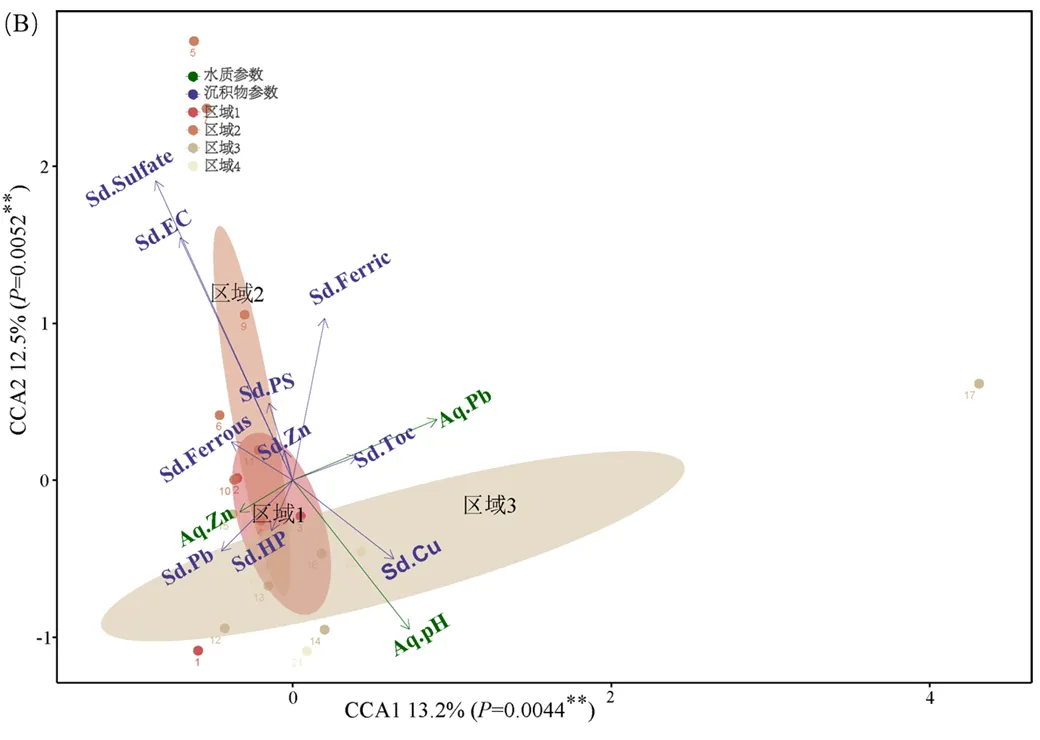
圖5 真核微生物和理化參數典型相關分析
每個點代表相應樣品中的真核微生物,半透明的橢圓代表95%的置信區間,箭頭的方向和長短代表該理化參數和真核微生物群落結構的關系
3 討論
目前真核微生物群落響應酸性廢水只報道過真菌和綠藻,不同酸性礦山廢水環境中真核微生物占整體生物量比重差異巨大,如西班牙Rio Tinto礦區的酸性礦山廢水環境中真核微生物生物量占比很大,其生物量和生物多樣性遠超原核微生物[26],而其它一些酸性礦山廢水環境,如美國加利福尼亞的Richmond Mine,其極端污染區域真核微生物多樣性比細菌甚至古菌都要低[27].真核生物在礦山廢水生態環境物質和能量流動方面所起到的作用不容忽視[10],如耐酸的藻類和真菌能夠給異養菌(如硫酸鹽還原菌(SRB)和Fe3+還原菌)提供必需的有機碳[10],通過吞食影響其它細菌和古菌的群落結構,或通過鐵氧化速率來調節酸性礦山廢水生成速率[28],并在酸性礦山廢水生態系統鐵/硫循環過程中發揮著重要作用[10].
結合對照河流沉積物中真核微生物分析,橫石河優勢種群有真菌門(Fungi)(4.51%~86.69%)、綠藻門(Chlorophyta)(0.61%~77.36%)和纖毛蟲門(Ciliophora) (0.81%~34.91%).不同污染區域物生物群落結構和多樣性有顯著差異,在上游污染較為嚴重的區域纖毛蟲門(Ciliophora)豐度較低,而隨著污染程度的降低,纖毛蟲門相對豐度逐漸升高.真核微生物群落的豐富度(Chao1)和均勻度(Shannon)也隨著污染程度的降低而顯著升高.
共現關系網絡分析發現一些真核微生物的存在與群落中其他真核微生物有著密切聯系,對這些微生物深入了解將有助于揭開真核微生物對酸性礦山廢水污染響應機制的神秘面紗.Acanthamoebidae和Sandonidae廣泛存在于淡水、土壤等各類環境介質中[29];Agaricomycetes可以存在于酸性環境中[30],具有金屬氧化功能,尤其能夠將溶解態的二價錳氧化為礦物態的三價/四價錳[31],多用于錳污染修復; Chlorellales[32]和Chlamydomona[33]是極端環境中常見的異養藻類,能夠在碳源稀缺,理化條件惡劣的環境中生存[34];Heterolobosea是一種在酸性礦山廢水環境中廣泛存在[35]并且能夠在極端環境中保持極高的代謝活力[6];Mucoromycotina之前多在植物根際土壤中發現,能夠幫助植物吸收礦物質[35]; Strombidiidae可以在高鹽水體中作為優勢種群生存[36-37].這些物種在重金屬降解、固碳、礦物吸收和能量代謝方面發揮著重要作用.
在酸性礦山廢水環境中,對原核微生物群落影響較大的環境理化因子主要有pH值[38-39]、鐵濃度梯度[40]、氧化梯度[41]、重金屬濃度[42]等.對大寶山礦區污染的橫石河區域的研究表明影響原核微生物Alpha多樣性的前3個因素分別是沉積物中電導率(Sd.EC)、沉積物pH值(Sd.pH)和水體中(Aq.Pb)[9].在本研究中,斯皮爾曼相關性分析結果顯示,與橫石河沉積物中真核微生物群落Alpha多樣性(Chao1和Shannon指數)顯著正相關的環境理化因子有:水體pH值(Aq.pH)和沉積物pH值(Sd.pH);與其顯著負相關的環境理化因子有:水體中Cu(Aq.Cu)、Cd(Aq. Cd)、Zn(Aq.Zn)、Pb(Aq.Pb)的濃度.另外,沉積物中三價鐵(Sd.Ferric)和總鐵(Sd.Total.Fe)、硫酸根(Sd.Sulfate)、可提取態銅(Cu.Exchangeable)、鋅(Zn.Exchangeable)、鎘(Cd.Exchangeable)和還原態鎘(Cd.Reducible)也和多種真核微生物相對豐度有著顯著的相關性關系,表明這些環境理化因子或許是酸性礦山廢水環境中真核微生物群落面臨的主要環境壓力.隨后使用CCA分析進一步對影響微生物群落結構的環境因子進行甄別,限制性排序分析結果顯示對真核微生物群落結構影響最大的環境理化因子是沉積物中硫酸根(Sd.Sulfate)和電導率(Sd.EC),其次才是水體pH值(Aq.pH)、水中鉛的含量(Aq.Pb)和沉積物中三價鐵的含量(Sd.Ferric).由此可見原核微生物和真核微生物對同樣環境脅迫響應并不相同,原核微生物對河流環境中pH值、鐵含量、氧化梯度、重金屬濃度的變化比較敏感,而真核微生物則對硫酸根和電導率變化響應更為強烈.與Alpha多樣性顯著相關的環境理化因子中Aq.pH、AqPb和Aq.Zn對真核微生物群落結構有較為顯著的影響,而從CCA結果來看,Aq.Cu和Aq.Cd對真核微生物群落結構影響不大.之前有學者對西藏地區森林生態系統研究發現,pH值能夠決定土壤真菌群落的Alpha多樣性,但是對Beta多樣性沒有顯著的影響[43].因此,對真核微生物群落Alpha多樣性產生顯著影響的環境理化因子與影響Beta多樣性的環境理化因子可能不同,橫石河沉積物中對真核微生物群落影響最大的環境因子是硫酸根和電導率.
4 結論
4.1 沉積物中真核微生物群落Alpha多樣性隨酸性礦山廢水污染梯度降低而逐漸升高.
4.2 原核微生物和真核微生物對同樣環境脅迫響應并不相同.酸性礦山廢水環境沉積物中真核微生物群落結構的變化主要受硫酸根和電導率的影響.
[1] 張河民,鐘銘君,吳啟堂.石灰石溝-堆肥濕地系統處理酸性礦山廢水的研究[J]. 中國環境科學, 2015,35(10):3032-3040. Zhang H M, Zhong M S, Wu Q T. The treatment of acidic mine drainage using limestone ditch and compost constructed wetland system. [J]. China Environmental Science, 2015,35(10):3032-3040.
[2] Galhardi J A, Bonotto D M. Hydrogeochemical features of surface water and groundwater contaminated with acid mine drainage (AMD) in coal mining areas: a case study in southern Brazil [J]. Environmental Science and Pollution Research, 2016,23(18):18911-18927.
[3] 王繼勇,肖 挺,何 偉.一株耐酸SRB的分離及其脫硫除鎘性能[J]. 中國環境科學, 2018,38(11):4255-4260.Wang J Y, Xiao T, He W. Isolation and characterization of an acid- resistance SRB strain with the efficient of desulfurization and cadmium- removal. [J]. China Environmental Science, 2018,38(11):4255-4260.
[4] Dean A P, Hartley A, McIntosh O A, et al. Metabolic adaptation of a Chlamydomonasacidophila strain isolated from acid mine drainage ponds with low eukaryotic diversity [J]. Science of the Total Environment, 2019,647:75-87.
[5] Kefeni K K, Msagati T A M, Mamba B B. Acid mine drainage: Prevention, treatment options, and resource recovery: A review [J]. Journal of Cleaner Production, 2017,151:475-493.
[6] Baker B J, Banfield J F. Microbial communities in acid mine drainage [J]. Fems Microbiology Ecology, 2003,44(2):139-152.
[7] Denef V J, Mueller R S, Banfield J F. AMD biofilms: using model communities to study microbial evolution and ecological complexity in nature [J]. Isme Journal, 2010,4(5):599-610.
[8] Aguinaga O E, Wakelin J F T, White K N, et al. The association of microbial activity with Fe, S and trace element distribution in sediment cores within a natural wetland polluted by acid mine drainage [J]. Chemosphere, 2019,231:432-441.
[9] Zhang X H, Tang S, Wang M, et al. Acid mine drainage affects the diversity and metal resistance gene profile of sediment bacterial community along a river [J]. Chemosphere, 2019,217:790-799.
[10] Chen L X, Huang L N, Mendez-Garcia C, et al. Microbial communities, processes and functions in acid mine drainage ecosystems [J]. Current Opinion in Biotechnology, 2016,38:150-158.
[11] Das B K, Roy A, Koschorreck M, et al. Occurrence and role of algae and fungi in acid mine drainage environment with special reference to metals and sulfate immobilization [J]. Water Research, 2009,43(4):883-894.
[12] Larson C. Environmental science. China gets serious about its pollutant-laden soil [J]. Science, 2014,343(6178):1415-1416.
[13] GB/T 5522-2008 糧油檢驗糧食、油料的過氧化氫酶活動度的測定 [S]. GB/T 5522-2008 Inspection of grains and oils - Determination of hydrogen peroxidase of grain and oilseeds [S].
[14] HJ 802-2016 土壤電導率的測定電極法 [S]. HJ 802-2016 Soil quality-Determination of conductivity-Electrode method [S].
[15] HJ 695-2014 土壤有機碳的測定燃燒氧化-非分散紅外法 [S]. HJ 695-2014 Soil-Determination of organic carbon-Combustion oxidation nondispersive infrared absorption method [S].
[16] Rauret G, López-Sánchez J F, Sahuquillo A, et al. Improvement of the BCR three step sequential extraction procedure prior to the certification of new sediment and soil reference materials [J]. Journal of Environmental Monitoring, 1999,1(1):57-61.
[17] Amaral-Zettler L A, McCliment E A, Ducklow H W, et al. A method for studying protistan diversity using massively parallel sequencing of V9hypervariable regions of small-subunit ribosomal RNA genes [J]. PLoS One, 2009,4(7):Article e6372.
[18] Fierer N, Hamady M, Lauber CL, et al. The influence of sex, handedness, and washing on the diversity of hand surface bacteria [J]. Proceedings of the National Academy of Sciences of the United States of America, 2008,105(46):17994-17999.
[19] Field D, Tiwari B, Booth T, et al. Open software for biologists: from famine to feast [J]. Nat Biotech, 2006,24(7):801-803.
[20] Caporaso J G, Kuczynski J, Stombaugh J, et al. QIIME allows analysis of high-throughput community sequencing data [J]. Nature methods, 2010,7(5):335-336.
[21] Edgar R C. UPARSE: highly accurate OTU sequences from microbial amplicon reads [J]. Nature Methods, 2013,10(10):996-998.
[22] Wang Q, Garrity G M, Tiedje J M, et al. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy [J]. Applied and Environmental Microbiology, 2007,73(16):5261-5267.
[23] Guillou L, Bachar D, Audic S, et al. The Protist Ribosomal Reference database (PR2): a catalog of unicellular eukaryote Small Sub-Unit rRNA sequences with curated taxonomy [J]. Nucleic Acids Research, 2013,41(1):597-604.
[24] Frontier S. Diversity and structure in aquatic ecosystems [J]. Oceanography and Marine Biology, 1985,23:253-312.
[25] Bastian M, Heymann S, Jacomy M. Gephi: An open source software for exploring and manipulating networks [C]. Proceedings of the Third International ICWSM Conference, 2009:361-362.
[26] Aguilera A, Zettler E, Gómez F, et al. Distribution and seasonal variability in the benthic eukaryotic community of Río Tinto (SW, Spain), an acidic, high metal extreme environment [J]. Systematic and Applied Microbiology, 2007,30(7):531-546.
[27] Baker B J, Tyson G W, Goosherst L, et al. Insights into the diversity of eukaryotes in acid mine drainage biofilm communities [J]. Applied and Environmental Microbiology, 2009,75(7):2192-2199.
[28] Baker B J, Lutz M A, Dawson S C, et al. Metabolically active eukaryotic communities in extremely acidic mine drainage [J]. Applied and Environmental Microbiology, 2004,70(10):6264-6271.
[29] De Jonckheere J F. Molecular identification of free-living amoebae of the Vahlkampfiidae and Acanthamoebidae isolated in Arizona (USA) [J]. European Journal of Protistology, 2007,43(1):9-15.
[30] Amaral-Zettler L. Eukaryotic diversity at pH extremes [J]. Frontiers in Microbiology, 2013,3: Article 441.
[31] Chaput D L, Hansel C M, Burgos W D, et al. Profiling microbial communities in manganese remediation systems treating coal mine drainage [J]. Applied and Environmental Microbiology, 2015,81(6):2189-2198.
[32] Tittel J, Bissinger V, Gaedke U, et al. Inorganic carbon limitation and mixotrophic growth in chlamydomonas from an acidic mining lake [J]. Protist, 2005,156(1):63-75.
[33] Vishnivetskaya T A. Viable cyanobacteria and green Aalgae from the permafrost darkness [M]. In Soil Biology, Margesin R, 2009,16:73-84.
[34] Méndez-García C, Peláez A I, Mesa V, et al. Microbial diversity and metabolic networks in acid mine drainage habitats [J]. Frontiers in Microbiology, 2015,6:Article 475.
[35] Delaux P M, Radhakrishnan G V, Jayaraman D, et al. Algal ancestor of land plants was preadapted for symbiosis [J]. Proceedings of the National Academy of Sciences of the United States of America, 2015,112(43):13390-13395.
[36] Edgcomb V, Orsi W, Leslin C, et al. Protistan community patterns within the brine and halocline of deep hypersaline anoxic basins in the eastern Mediterranean Sea [J]. Extremophiles, 2009,13(1):151-167.
[37] Stock A, Edgcomb V, Orsi W, et al. Evidence for isolated evolution of deep-sea ciliate communities through geological separation and environmental selection [J]. BMC Microbiology, 2013,13:Article 150.
[38] Bonilla J O, Kurth D G, Cid F D, et al. Prokaryotic and eukaryotic community structure affected by the presence of an acid mine drainage from an abandoned gold mine [J]. Extremophiles, 2018,22(5):699-711.
[39] Mendez M O, Neilson J W, Maier R M. Characterization of a bacterial community in an abandoned semiarid lead-zinc mine tailing site [J]. Applied and environmental microbiology, 2008,74(12):3899-3907.
[40] García-Moyano A, González-Toril E, Aguilera á, et al. Comparative microbial ecology study of the sediments and the water column of the Río Tinto, an extreme acidic environment [J]. FEMS Microbiology Ecology, 2012, 81(2):303-314.
[41] Gonzalez-Toril E, Aguilera A, Souza-Egipsy V, et al. Geomicrobiology of La Zarza-Perrunal acid mine effluent (Iberian Pyritic Belt, Spain) [J]. Applied and Environmental Microbiology, 2011,77(8):2685-2694.
[42] Yang Y, Li Y, Sun Q Y. Archaeal and bacterial communities in acid mine drainage from metal-rich abandoned tailing ponds, Tongling, China [J]. Transactions of Nonferrous Metals Society of China, 2014,24(10):3332-3342.
[43] Wang J T, Zheng Y M, Hu H W, et al. Soil pH determines the alpha diversity but not beta diversity of soil fungal community along altitude in a typical Tibetan forest ecosystem [J]. Journal of Soils and Sediments, 2015,15(5):1224-1232.
Effects of acid mine drainage on eukaryotic community in river sediments.
LIU Fan1, ZHANG Xiao-hui1, TANG Song2, WANG Mao3, LIU Hong-ling1*
(1.State Key Laboratory of Pollution Control & Resource Reuse, School of the Environment, Nanjing University, Nanjing 210023, China;2.National Institute of Environmental Health, Chinese Center for Disease Control and Prevention, Beijing 100021, China;3.School of Public Health, Sun Yat-sen University, Guangzhou 510080, China)., 2019,39(12):5285~5292
Analytical and molecular biology methods were integrated together to investigate the physicochemical factors in water and sediments of Hengshi River as well as the diversity of eukaryotic microorganism community in the sediments. Spearman correlation and canonical correspondence analysis (CCA) were further applied to identify the major factors affecting eukaryotic microorganism community. Results of 18S rRNA gene sequencing indicated that Eukaryotic community of Hengshi River was dominated by Fungi (4.51%~86.69%), Chlorophyta (61%~77.36%) and Ciliophora (0.81%~34.91%). Both the abundance and evenness of eukaryotic communities in sediments significantly and gradually increased along the Hengshi River with decreasing contamination, which suggested that composition of eukaryotic microorganism community partly reflected changes in physicochemical conditions. Sulfate and electric conductivity of the sediments were the major abiotic factors altering the eukaryotic communities in the sediments of AMD impacted Hengshi River, which was different from the findings of prokaryotic microbial communities.
acid mine drainage;eukaryotic microorganisms;sulfate;electric conductivity;next-generation sequencing
X172
A
1000-6923(2019)12-5285-08
劉 帆(1995-),女,山東濟南人,南京大學碩士研究生,研究方向為環境毒理學與風險評價.
2019-05-24
國家自然科學基金(21677073);國家重點研發項目(2018YFC1801505);國家科技重大專項(2018ZX07208001,2017ZX07301002)
* 責任作者, 副教授, hlliu@nju.edu.cn
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
河北地質(2021年2期)2021-08-21 02:43:50
神劍(2021年3期)2021-08-14 02:30:08
昆鋼科技(2021年2期)2021-07-22 07:47:06
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
礦產勘查(2020年7期)2020-12-25 02:43:42
中國生殖健康(2020年6期)2020-02-01 06:28:50
當代陜西(2019年7期)2019-04-25 00:22:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
領導決策信息(2018年26期)2018-10-12 02:18:26
