內質網通道蛋白Sec61在PM2.5誘導支氣管上皮細胞BEAS-2B高表達MUC5AC中的作用①
2019-12-27 06:53:02林秀山周向東王才春
中國免疫學雜志 2019年22期
林秀山 周向東 王才春
(海南醫學院第一附屬醫院呼吸內科,海口 570102)
PM2.5是指空氣動力學直徑小于或等于2.5 μm的細顆粒物質,易被人體吸入到達遠端支氣管和肺泡,并且長期沉積在呼吸道中,是一系列呼吸系統疾病的危險因素,包括慢性支氣管炎、哮喘、肺癌等[1,2]。氣道上皮細胞在受到PM2.5刺激時常呈黏液高分泌狀態,具有量大、黏稠等特點,纖毛無法有效的清除,而過量的黏液積聚又會加重局部呼吸道的炎癥和感染,最終導致呼吸道慢性疾病的遷延不愈[3]。黏蛋白5AC(Mucin 5AC,MUC5AC)是氣道黏液的主要蛋白成分,在呼吸道中發揮潤滑和保濕作用,但是在PM2.5刺激下,MUC5AC大量分泌,且呈高度的硫酸化和糖基化,是黏液高分泌狀態的主要原因[4],因此探討PM2.5誘導支氣管上皮細胞MUC5AC高表達的調節機制,并開展針對性的干預具有重要的臨床意義。
Sec61是一種異源性三聚體,為內質網通道蛋白,參與調節核糖體生成的多肽鏈向內質網腔的轉運[5,6]。研究發現,Sec61與內質網應激密切相關,如王軍等[7]發現使用抑制劑阻斷Sec61可以降低內質網應激標志物GRP78的表達,緩解肝細胞脂肪變性的程度。此外,Sec61可以促進鈣離子從內質網向細胞質中釋放,在鈣離子紊亂中發揮重要的病理作用[8]。而內質網應激和鈣離子紊亂也是PM2.5誘導氣道黏液高分泌的病理機制[9],因此我們推測Sec61可能參與PM2.5誘導支氣管上皮細胞MUC5AC高表達,本文首先使用PM2.5對支氣管上皮細胞BEAS-2B進行刺激,以探討Sec61在不同濃度和不同時間刺激下的變化規律,然后使用RNA干擾技術沉默Sec61表達,以研究Sec61沉默對細胞中MUC5AC表達的影響及相關機制。
1 材料與方法
1.1材料 siRNA-Sec61和siRNA-Control序列由吉凱基因公司設計和合成;siRNA-Sec61正義鏈:5′-GAGAGAAAGAUUCAGUUUAdTdT-3′,反義鏈:5′-UAAACUGAAUCUUUCUCUCdTdT-3′;siRNA-Control正義鏈:5′-GAUAAUAAAUCUAAGGGGUdTdT-3′,反義鏈:5′-ACCCCUUAGAUUUAUUAUCdTdT-3′;懸浮微粒采樣器(TH-150)購自武漢市天虹儀表有限責任公司;支氣管上皮細胞BEAS-2B購自美國ATCC公司,胎牛血清、DMEM培養基購自Gibco公司;LipofectamineTM2000、Fluo-4/AM熒光染料購自Invitrogen公司;PMSF、RIPA裂解液購自上海碧云天;一抗購自Santa Cruz公司;二抗購自Proteintech公司;逆轉錄試劑盒和熒光定量PCR試劑盒購自TaKaRa公司;引物合成自上海生工。
1.2方法
1.2.1PM2.5的采集與處理 于海口市的主要交通路口處,在空氣質量指數大于5級、PM2.5嚴重超標的霧霾天氣時分次采集PM2.5顆粒,采集時間為2018年3~5月,共采集8次,每次時間為8:00~18:00。采用懸浮微粒采樣器收集PM2.5,結束后將纖維濾膜放入干燥器中暫時保存。將纖維濾膜浸入去離子水,超聲振蕩30 min,洗脫液10 000 r/min離心30 min,將沉淀物真空凍干后稱重,于-20℃冰箱中保存。使用PBS將PM2.5配置成0、25、50、100、200 μg/ml[10,11]。
1.2.2細胞培養與處理 使用含10%胎牛血清的DMEM培養基培養BEAS-2B細胞,置于37℃、5%CO2的培養箱中。當細胞融合度達到70%時,更換為無血清培養基繼續培養6 h,然后依次加入0、25、50、100、200 μg/ml的PM2.5刺激48 h,收集細胞,使用蛋白質免疫印跡(Western blot)檢測細胞中Sec61的表達。使用100 μg/ml的PM2.5分別刺激細胞0、6、12、24、48 h,收集細胞,Western blot檢測細胞中Sec61的表達。使用100 μg/ml的PM2.5刺激48 h,細胞免疫熒光染色檢測Sec61的表達。
1.2.3細胞轉染與實驗分組 當細胞匯合度達到70%時,根據LipofectamineTM2000說明書分別將siRNA-Control和siRNA-Sec61干擾序列(工作濃度為50 nmol/L)轉染至BEAS-2B細胞。將實驗分為4組,即對照組、PM2.5組、siRNA-Control組、siRNA-Sec61組。對照組使用未進行轉染的BEAS-2B細胞,加入PBS作為對照;PM2.5組使用未進行轉染的BEAS-2B細胞,加入100 μg/ml的PM2.5刺激48 h;siRNA-Control組和siRNA-Sec61組分別使用轉染siRNA-Control或siRNA-Sec61序列的細胞,加入100 μg/ml的PM2.5刺激48 h。收集細胞,使用熒光定量聚合酶鏈式反應和Western blot檢測細胞中Sec61、MUC5AC、GRP78的表達,使用免疫熒光染色檢測細胞中MUC5AC和游離鈣離子的表達。
1.2.4蛋白質免疫印跡 使用含有PMSF的RIPA裂解液對細胞進行充分裂解,12 000 r/min離心5 min,收集上清液進行蛋白定量。取40 μg的總蛋白進行聚丙烯酰胺凝膠電泳,電轉膜后加入5%的牛血清白蛋白室溫封閉2 h,加入抗Sec61抗體(1∶500)、抗MUC5AC抗體(1∶1 000)、抗GRP78抗體(1∶500)和抗GAPDH抗體(1∶3 000)4℃過夜孵育。洗滌后,加入含辣根過氧化物酶的山羊抗兔抗體室溫孵育1 h,經洗滌、顯色后,使用凝膠成像儀拍照。使用Image Pro Plus 6.0軟件測定各條帶的灰度值,將對照組中目的蛋白與GAPDH蛋白的灰度值之比作為100。
1.2.5熒光定量聚合酶鏈式反應(qRT-PCR) Trizol法提取細胞中的總RNA,根據逆轉錄試劑盒說明書逆轉錄為cDNA,然后使用熒光定量PCR試劑盒擴增目的基因,反應條件:95℃預變性5 min;95℃變性30 s,55℃退火30 s,72℃延伸60 s,共進行40個循環,各基因的引物序列見表1。取擴增產物進行瓊脂糖凝膠電泳,使用凝膠成像儀拍照,Image Pro Plus 6.0軟件測定各條帶的灰度值,將對照組中目的基因與GAPDH基因的灰度值之比作為100。
1.2.6細胞免疫熒光染色 4%的多聚甲醛固定細胞30 min,5%的牛血清白蛋白封閉1 h,加入抗Sec61抗體(1∶100)和抗MUC5AC抗體(1∶100)4℃過夜。次日,PBS洗滌后加入含有FITC熒光標簽的山羊抗兔二抗37℃孵育30 min,PBS洗滌后DAPI染色10 min,洗滌后熒光顯微鏡下拍照。
1.2.7胞內游離鈣離子的檢測 PBS洗滌細胞2次后,加入Fluo-4/AM熒光染料(5 μmol/L)于37℃避光孵育60 min,PBS洗滌,于37℃繼續孵育20 min,以保證Fluo-4/AM在細胞內充分轉化為Fluo-4,然后使用激光共聚焦顯微鏡觀察和拍照,激發波長為488 nm,發射波長為516 nm[12]。
2 結果
2.1PM2.5刺激對Sec61表達的影響 隨著PM2.5刺激濃度的增加和刺激時間的延長,Sec61蛋白質的表達量逐漸升高,刺激濃度為100 μg/ml、刺激時間為48 h時,Sec61的表達量最高,見圖1。使用100 μg/ml的PM2.5刺激48 h,細胞中Sec61的免疫熒光強度明顯高于對照組,見圖2。
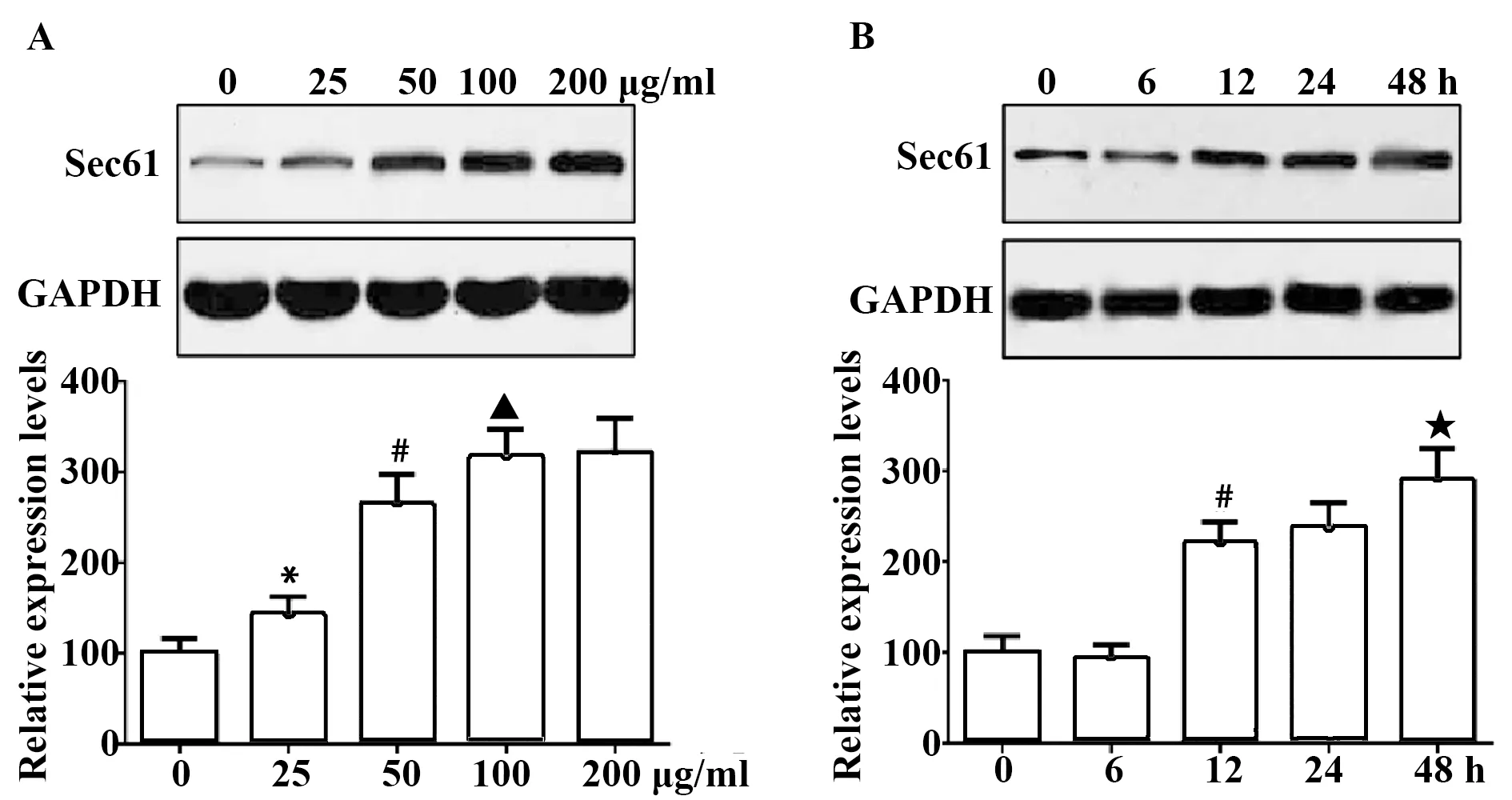
圖1 Western blot檢測PM2.5刺激對Sec61表達的影響Fig.1 Influence of PM2.5 stimulation on Sec61 express-ion was examined by Western blotNote:A.Influence of different stimulation concentrations on Sec61 expression;B.Influence of different stimulation time on Sec61 expression.Compared with 0 μg/ml group,*.P<0.05;compared with 25 μg/ml,#.P<0.05;compared with 100 μg/ml,▲.P<0.05;compared with 200 μg/ml,★.P<0.05.
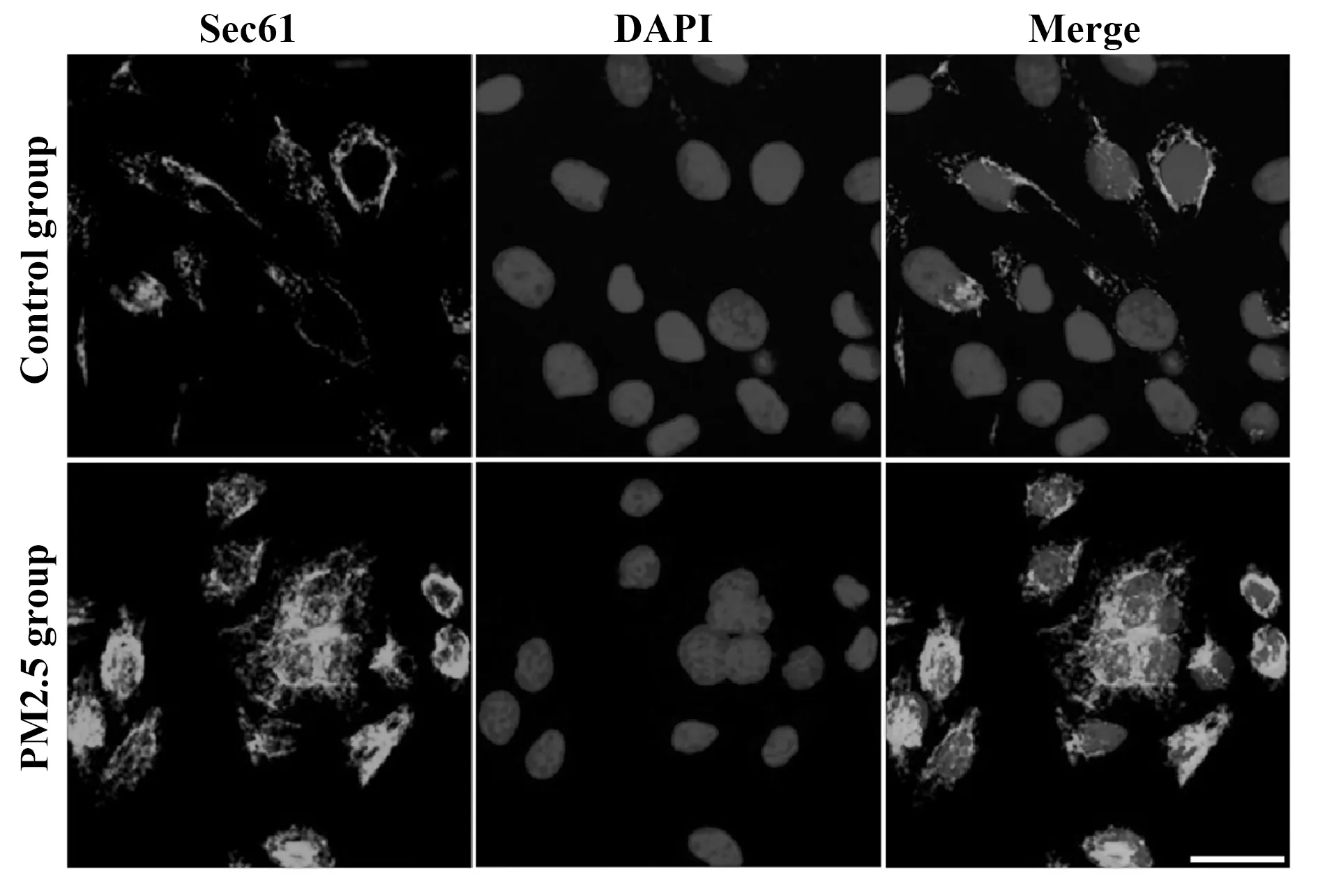
圖2 細胞免疫熒光染色檢測PM2.5刺激(100 μg/ml,48 h)對Sec61表達的影響Fig.2 Influence of PM2.5 stimulation (100 μg/ml,48 h) on Sec61 expression was examined by immunofluorescence stainingNote:Scale bar.25 μm.
2.2Sec61沉默對Sec61表達的抑制效果 與對照組比較,PM2.5組細胞中Sec61 mRNA和蛋白質的表達量升高,差異具有統計學意義(P<0.05);與PM2.5組比較,siRNA-Control組細胞中Sec61 mRNA和蛋白質的表達量差異無顯著統計學意義(P>0.05);與siRNA-Control組比較,siRNA-Sec61組細胞中Sec61 mRNA和蛋白質的表達量降低,差異具有統計學意義(P<0.05),見圖3。
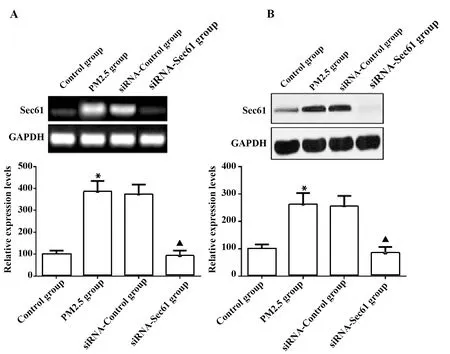
圖3 Sec61沉默對Sec61表達的抑制效果Fig.3 Inhibitory effect of Sec61 silencing on Sec61expressionNote:A.qRT-PCR was used to detect the mRNA expression of Sec61;B.Western blot was used to detect the protein expression of Sec61.Compared with contrd group,*.P<0.05;compared with siRNA-control group;▲.P<0.05.
表1 引物序列
Tab.1 Primer sequence
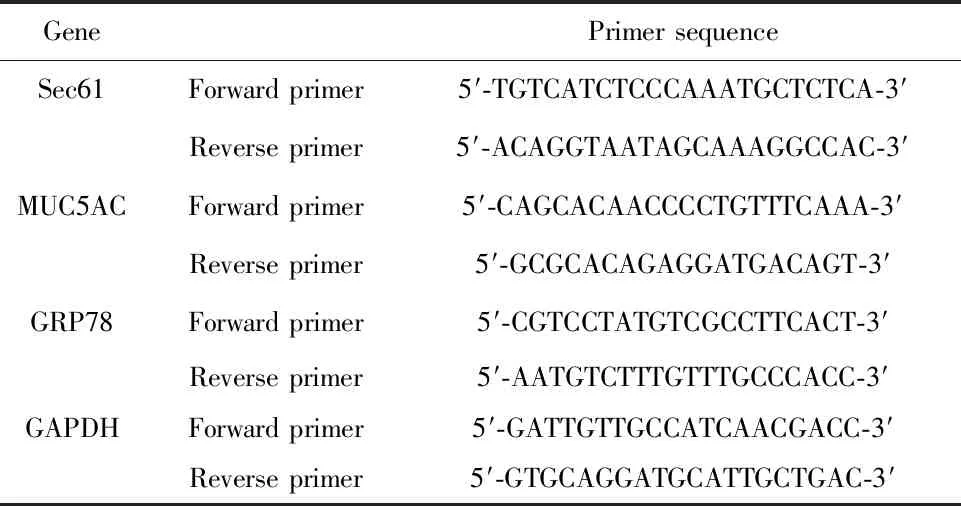
GenePrimer sequenceSec61Forward primer5′-TGTCATCTCCCAAATGCTCTCA-3′Reverse primer5′-ACAGGTAATAGCAAAGGCCAC-3′MUC5ACForward primer5′-CAGCACAACCCCTGTTTCAAA-3′Reverse primer5′-GCGCACAGAGGATGACAGT-3′GRP78Forward primer5′-CGTCCTATGTCGCCTTCACT-3′Reverse primer5′-AATGTCTTTGTTTGCCCACC-3′GAPDHForward primer5′-GATTGTTGCCATCAACGACC-3′Reverse primer5′-GTGCAGGATGCATTGCTGAC-3′
2.3Sec61沉默對MUC5AC表達的影響 與對照組相比較,PM2.5組細胞中MUC5AC mRNA和蛋白質的表達量升高,差異具有統計學意義(P<0.05);與PM2.5組相比較,siRNA-Control組細胞中MUC5AC mRNA和蛋白質的表達量差異無顯著統計學意義(P>0.05);與siRNA-Control組相比較,siRNA-Sec61組細胞中MUC5AC mRNA和蛋白質的表達量降低,差異具有統計學意義(P<0.05),見圖4。
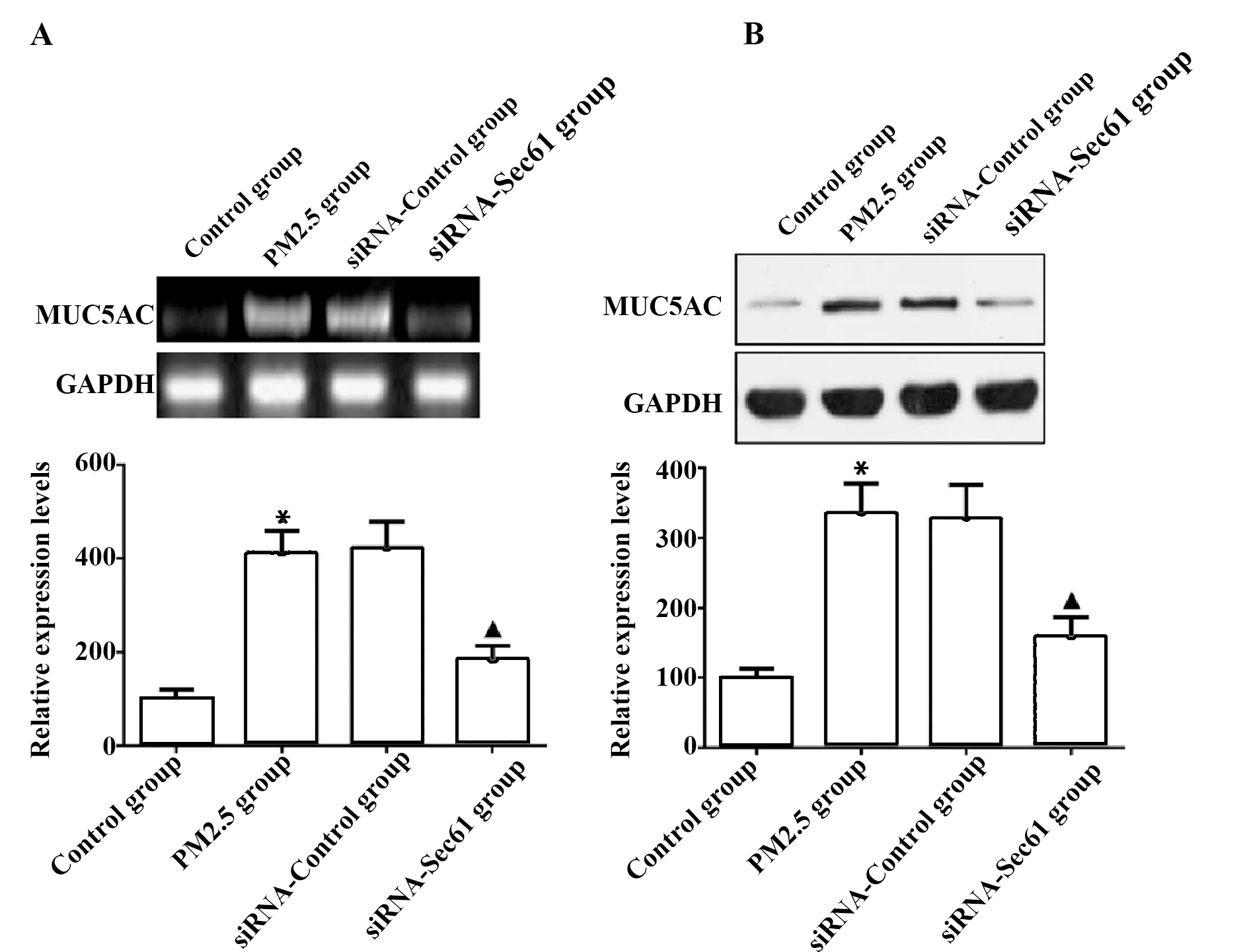
圖4 檢測Sec61沉默對MUC5AC表達的影響Fig.4 Influence of Sec61 silencing on MUC5AC express-ion was examinedNote:A.qRT-PCR was used to detect the mRNA expression of MUC5AC;B.Western blot was used to detect the protein expression of MUC5AC.Compared with control group,*.P<0.05;compared with PM2.5 group,#.P<0.05;compared with siRNA-control group,▲.P<0.05.
與對照組相比較,PM2.5組細胞中MUC5AC的熒光強度明顯升高;與PM2.5組相比較,siRNA-Control組細胞中MUC5AC的熒光強度無明顯差異;與siRNA-Control組相比較,siRNA-Sec61組細胞中MUC5AC的熒光強度明顯降低,見圖5。
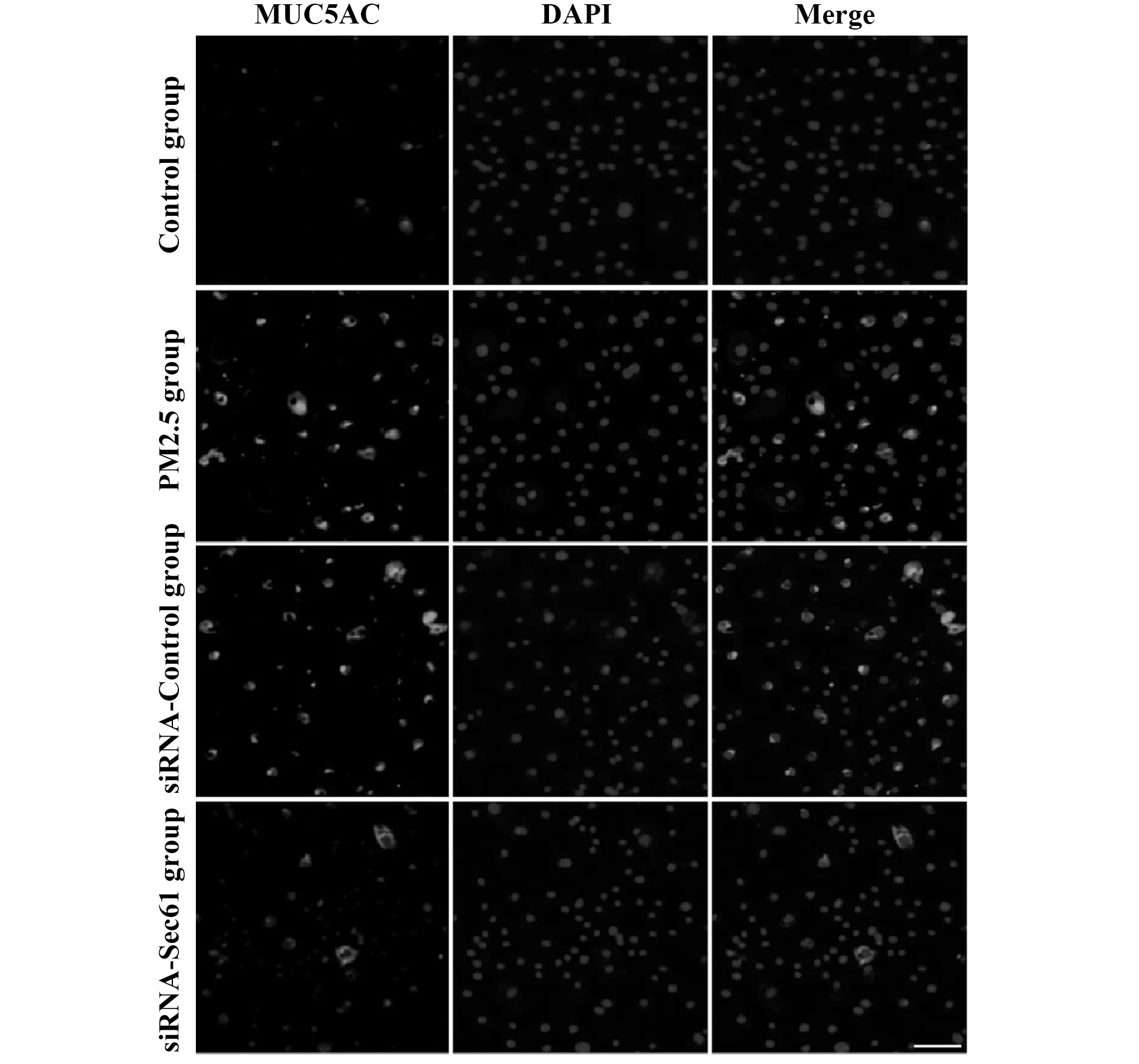
圖5 細胞免疫熒光染色檢測Sec61沉默對MUC5AC表達的影響Fig.5 Influence of Sec61 silencing on MUC5AC express-ion was examined by immunofluore-scence stainingNote:Scale bar.50 μm.
2.4Sec61沉默對GRP78表達的影響 與對照組相比較,PM2.5組細胞中GRP78 mRNA和蛋白質的表達量升高,差異具有統計學意義(P<0.05);與PM2.5組相比較,siRNA-Control組細胞中GRP78 mRNA和蛋白質的表達量差異無顯著統計學意義(P>0.05);與siRNA-Control組相比較,siRNA-Sec61組細胞中GRP78 mRNA和蛋白質的表達量降低,差異具有統計學意義(P<0.05),見圖6。
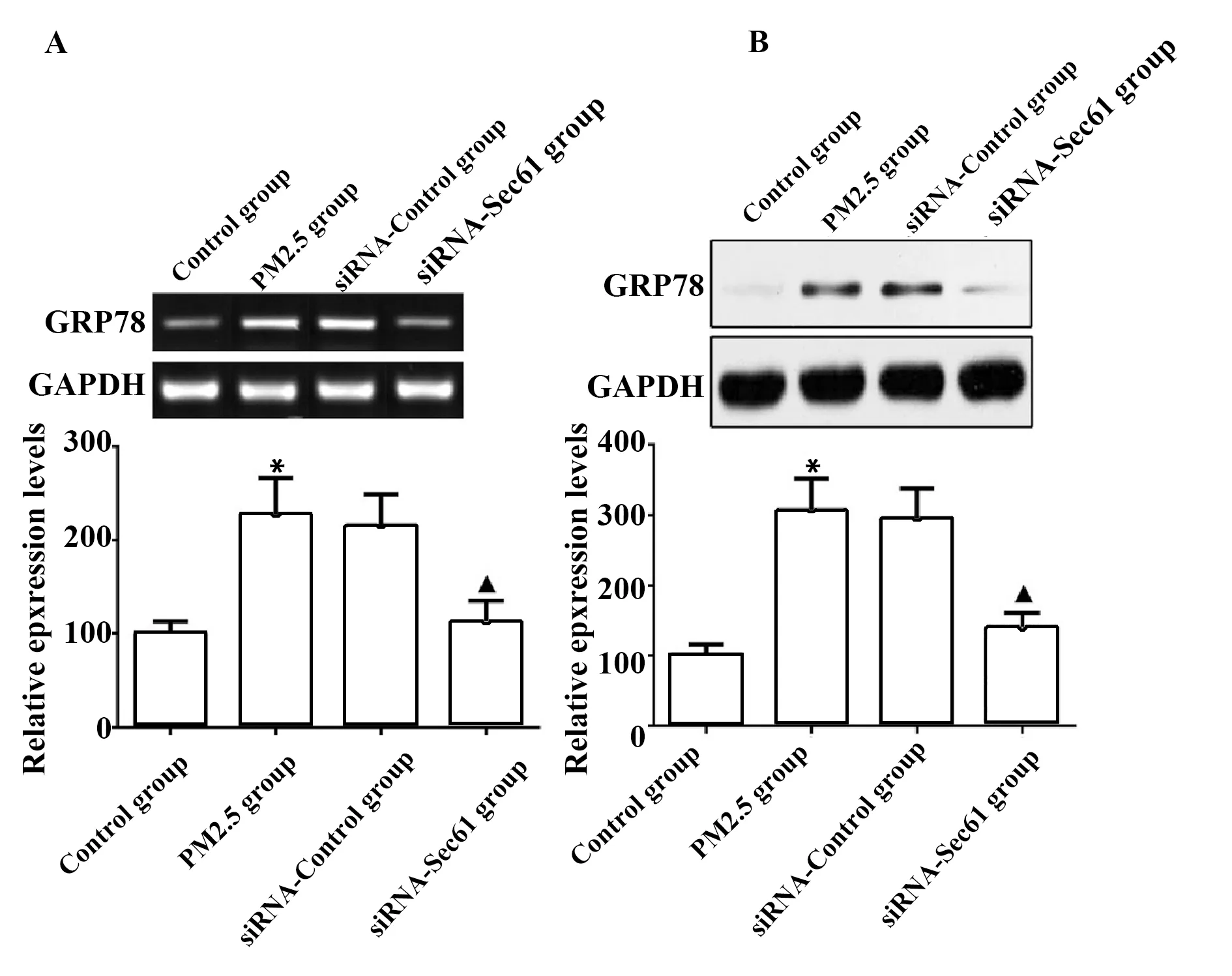
圖6 Sec61沉默對GRP78表達的影響Fig.6 Influence of Sec61 silencing on GRP78 expressionNote:A.qRT-PCR was used to detect the mRNA expression of GRP78;B.Western blot was used to detect the protein expression of GRP78.Compared with control group,*.P<0.05;compared with PM2.5 group,#.P<0.05;compared with siRNA-control group,▲.P<0.05.
2.5Sec61沉默對細胞內游離鈣離子的影響 與對照組比較,PM2.5組細胞中游離鈣離子的熒光強度明顯升高;與PM2.5組相比較,siRNA-Control組細胞中游離鈣離子的熒光強度無明顯差異;與siRNA-Control組相比較,siRNA-Sec61組細胞中游離鈣離子的熒光強度明顯降低,見圖7。
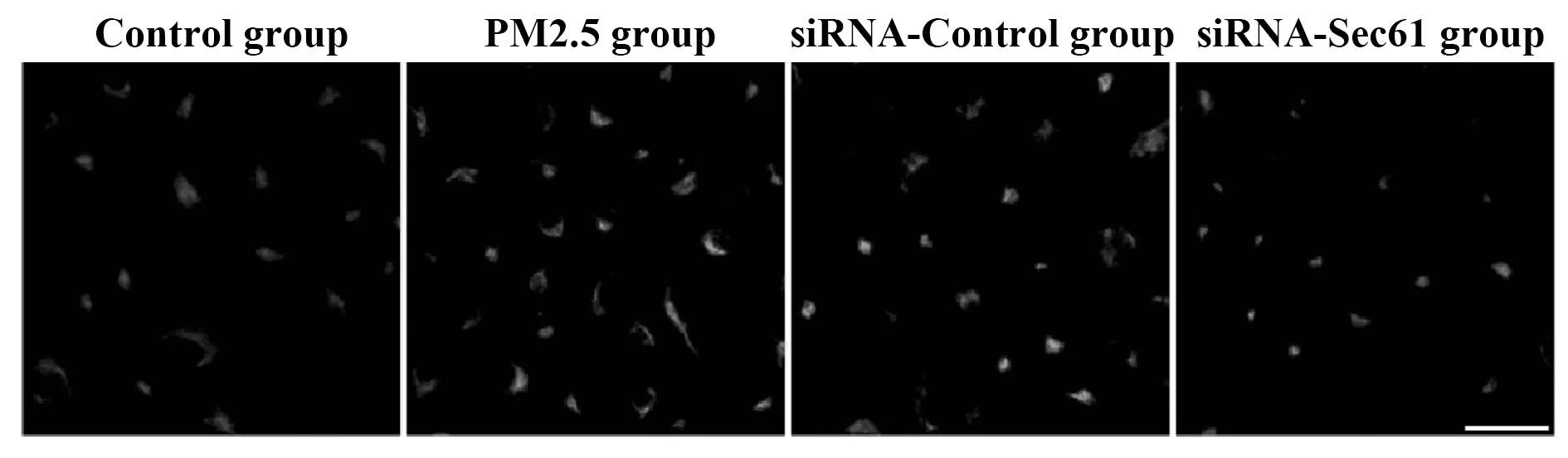
圖7 Sec61沉默對細胞內游離鈣離子的影響Fig.7 Influence of Sec61 silencing on concentration of intracellular calcium ionsNote:Scale bar.25 μm.
3 討論
PM2.5是空氣動力學直徑小于或等于2.5 mm的大氣顆粒物,是空氣污染的重要組成部分,主要包含重金屬顆粒、酸性氧化物、有機污染物、細菌、真菌、病毒等,吸入后可以沉積在遠端氣道和肺泡中,引起異常的免疫炎癥反應[13]。流行病學研究表明,空氣中PM2.5濃度每上升10 mg/m3,呼吸系統疾病的死亡率可增加2%,慢性阻塞性肺疾病患者的住院率可增加1.72%~6.87%[14],因此研究PM2.5對呼吸系統的損傷機制以及開展針對性的治療具有重要意義。
支氣管上皮細胞分泌黏液有助于清除進入肺部的病原體或顆粒物質,在呼吸道的防御機制中發揮重要作用,但是在PM2.5的長期刺激下,過多的黏液分泌會產生黏液栓,導致呼吸功能受損,甚至死亡。MUC5AC是呼吸道黏液的主要組成成分,可在病毒、細菌、煙霧等外界刺激物直接誘導產生,也可以在IL-4、IL-13、TNF-α等炎癥因子的間接作用下上調表達,因此在支氣管哮喘、呼吸道感染、慢性阻塞性肺疾病等呼吸系統疾病中大量表達和分泌[15-17]。同樣,PM2.5也可以誘導支氣管上皮細胞大量分泌MUC5AC,而且MUC5AC被糖基化和硫酸化修飾,使氣道黏液呈酸性和黏稠狀[3]。MUC5AC的初始肽鏈在細胞的核糖體中合成后,可轉運至內質網中進行三維結構的折疊與加工,糖基化和硫酸化修飾也在此完成。綜上,本文認為MUC5AC從核糖體向內質網的轉運過程在PM2.5誘導氣道上皮細胞黏液高分泌狀態中發揮關鍵作用,若阻斷該轉運過程中的關鍵分子可能在PM2.5相關損傷中發揮保護效應。
Sec61是位于內質網膜上的雙向通道蛋白,參與調節多肽鏈向內質網的轉運以及鈣離子向胞質的釋放[5]。為了研究Sec61是否參與PM2.5誘導MUC5AC高表達,本文使用PM2.5刺激支氣管上皮BEAS-2B細胞,結果發現隨著PM2.5刺激濃度的增加和刺激時間的延長,Sec61蛋白質的表達量逐漸升高,表明PM2.5可以誘導Sec61高表達,且呈濃度依賴性和時間依賴性。當刺激濃度為100 μg/ml、刺激時間為48 h時,Sec61的表達量最高,細胞中Sec61的免疫熒光強度也明顯高于對照組,因此本文在后續實驗中均使用該刺激條件進行研究。
本文設計和構建了Sec61的小分子RNA干擾序列,轉染后發現與siRNA-Control組比較,siRNA-Sec61組細胞中Sec61 mRNA和蛋白質的表達量降低,表明Sec61的小分子RNA干擾序列構建成功。為了研究Sec61的沉默是否影響MUC5AC的表達,本文分別使用RT-PCR、Western blot以及免疫熒光染色對MUC5AC的表達進行檢測,結果顯示與siRNA-Control組相比較,siRNA-Sec61組細胞中MUC5AC mRNA和蛋白質的表達量降低,免疫熒光強度也明顯降低,表明Sec61沉默可以抑制PM2.5誘導的MUC5AC高表達,這種現象的出現可能因為Sec61通道蛋白數量的降低,使MUC5AC新生肽鏈無法有效進入內質網腔,從而積聚在細胞質中,負反饋調節使MUC5AC mRNA轉錄和蛋白質翻譯水平降低。
內質網應激和鈣離子失衡在PM2.5誘導氣道上皮細胞損傷中發揮重要的病理作用,而且Sec61與內質網應激、鈣離子穩態密切相關,如Schauble等[18]發現綠膿桿菌外毒素A可以抑制鈣離子從內質網腔向胞質中釋放鈣離子,主要機制是其與Sec61α亞基的氨基端結合,介導Sec61通道蛋白的關閉;王軍等[7]也在脂肪變性的肝細胞中發現,下調Sec61蛋白的表達可以抑制內質網應激。為了進一步研究Sec61沉默抑制MUC5AC高表達的機制,本文對細胞的內質網應激水平和胞內游離鈣離子濃度進行檢測,結果顯示與siRNA-Control組相比較,siRNA-Sec61組細胞中內質網應激標志物GRP78 mRNA和蛋白質的表達量降低,細胞中游離鈣離子的熒光強度明顯降低,表明Sec61沉默可以降低PM2.5誘導的內質網應激,抑制鈣離子從內質網腔向胞質的釋放。
本文的不足和展望:①本文僅使用了體外細胞學實驗進行研究,但是細胞學實驗具有穩定性差、無法模擬體內環境等缺點,相關結論仍需要構建相關的動物模型進行驗證;②本文使用小分子RNA干擾的方法抑制Sec61的表達,但是該方法發揮抑制效應的時間較短,一般在7 d左右,因此未來需要進一步開發穩定、高效、安全的抑制藥物應用于臨床。
綜上所述,PM2.5可以誘導Sec61高表達,呈濃度依賴性和時間依賴性。使用小分子RNA沉默Sec61表達可以顯著抑制MUC5AC高表達,可能與調節內質網應激和鈣離子穩態有關。