長鏈非編碼RNA NEAT1對非小細胞肺癌增殖和凋亡的影響及其機制①
2019-12-27 06:07:30李寶輝侯明明李新萌孔祥軍
中國免疫學雜志 2019年23期
李寶輝 馬 輝 侯明明 李新萌 陳 虎 孔祥軍
(滄州市中心醫院中心實驗室,滄州 061001)
肺癌是世界上癌癥相關死亡的最常見原因之一,非小細胞肺癌(Non-small cell lung cancer,NSCLC)作為肺癌的主要亞型占所有病例的70%~80%[1]。雖然肺癌的診斷和治療取得了很大進展,但總體生存率仍然不高,高死亡率可能與早期腫瘤轉移和復發有關[2]。因此,迫切需要更好地了解NSCLC進展中涉及的分子機制,并為患者找到更有效的治療方法。近年來研究發現,許多長鏈非編碼RNA(long non-coding RNAs,lncRNAs)在生物過程中發揮重要作用,包括細胞增殖,轉移和分化[3-5]。許多失調的lncRNA參與NSCLC的各種生物學活性,例如,lncRNA PTV1、MALAT-1和HOTAIR[6-8]。然而,很少有研究關注富含核的豐富轉錄物1(Nuclear enriched abundant transcript 1,NEAT1)在NSCLC中的作用。本研究探討了NEAT1在NSCLC中的潛在功能,并初步分析了其相關機制,Wnt/β-catenin信號通路在調節腫瘤發展的多個方面(包括NSCLC)中起重要作用,NEAT1是一種新型的lncRNA,在許多癌癥中起著重要的調節作用,因此我們進一步探討了NEAT1是否可能參與Wnt/β-catenin信號通路。
1 材料與方法
1.1材料
1.1.1試劑及儀器 非小細胞肺癌細胞系A549、SPCA1和正常細胞系BESA-2B均購自中國科學院生物化學與細胞生物學研究所,Transwell、SOX9及GAPDH一抗購自美國BD公司,RPMI1640培養基購自美國Gibco公司,Trizol和細胞轉染試劑購自美國Invitrogen公司,一步法RT-PCR試劑盒購自大連寶生物工程有限公司,熒光檢測試劑盒購自美國Promega公司。實驗所用PCR儀為ABI 7500,酶標儀為Thermo FC。
1.1.2組織標本 收集自2015年1月至2016年12月來我院就診經病理學診斷確診為NSCLC的患者91例,手術前未進行局部或全身治療,獲取其NSCLC組織和相鄰非腫瘤組織,液氮中快速冷凍后于-80℃超低溫冰箱中保存。本研究已獲得滄州市中心醫院倫理委員會批準(2017-139-01)。
1.2方法
1.2.1細胞培養、轉染及分組 NSCLC細胞系A549、SPCA1及正常人肺細胞系BESA-2B均用含10%胎牛血清、100 U/ml青霉素和100 mg/ml鏈霉素的RPMI1640培養基于37℃、5%CO2培養,48 h消化傳代。分5組進行細胞轉染:si-NEAT1組、si-NC組(陰性對照組)、miR-101-3p模擬物組、miR-101-3p抑制劑組和si-SOX9組,擴增引物見表1,采用2-ΔΔCt法進行相對定量。
1.2.2MTT實驗 應用改良MTT實驗評估細胞活力,連續觀測96孔板(5 000細胞/孔)24 h。細胞轉染48 h后,加入20 μl MTT(5 mg/ml),繼續培養4 h,棄上清后加入總共200 μl二甲基亞砜(DMSO),測量490 nm吸光度。

表1 NEAT1、miR-101-3p擴增引物Tab.1 Amplification primer of NEAT1 and miR-101-3p
1.2.3Transwell實驗 通過Transwell實驗檢測細胞遷移和浸潤,應用Transwell小室檢測細胞遷移,細胞浸潤Transwell小室以30 mg/cm2基底膜(BD)包被后于37℃孵育1 h形成基質屏障。DMEM稀釋細胞至1×105細胞/孔,5%CO237℃孵育24 h后,甲醛固定10 min,結晶紫染色10 min,PBS洗滌后,顯微鏡下隨機選取10個視野(200×)計算膜下細胞數,重復3次,侵襲細胞數越多表示侵襲能力越強。
1.2.4Western blot實驗 通過免疫印跡實驗檢測NSCLC細胞中SOX9、β-catenin和c-Myc的表達情況。培養或轉染的細胞以RIPA緩沖液裂解,BCA法提取蛋白質定量,10%SDS-PAGE電泳,轉膜后于5%脫脂牛奶(含0.1%吐溫的PBS)孵育1 h,加入anti-SOX9、anti-β-catenin、anti-c-Myc和anti-GAPDH一抗4℃過夜,加入辣根過氧化物酶標記的二抗(1∶5 000,Abcam)孵育,EMD顯影,GAPDH為蛋白標準。

2 結果
2.1NEAT1在NSCLC組織和細胞中高表達 通過qRT-PCR檢測NSCLC組織、癌旁正常組織以及肺癌細胞系A549和SPCA1中NEAT1 mRNA的表達,從表2中可見NSCLC組織中NEAT1 mRNA表達量明顯高于正常組織(P<0.05),NSCLC細胞系A549和SPCA1中NEAT1 mRNA表達量也顯著高于人類正常肺細胞系BESA-2B(P<0.05)。另外,NEAT1的表達量與NSCLC患者的TNM分期及淋巴轉移顯著相關(P=0.000)。
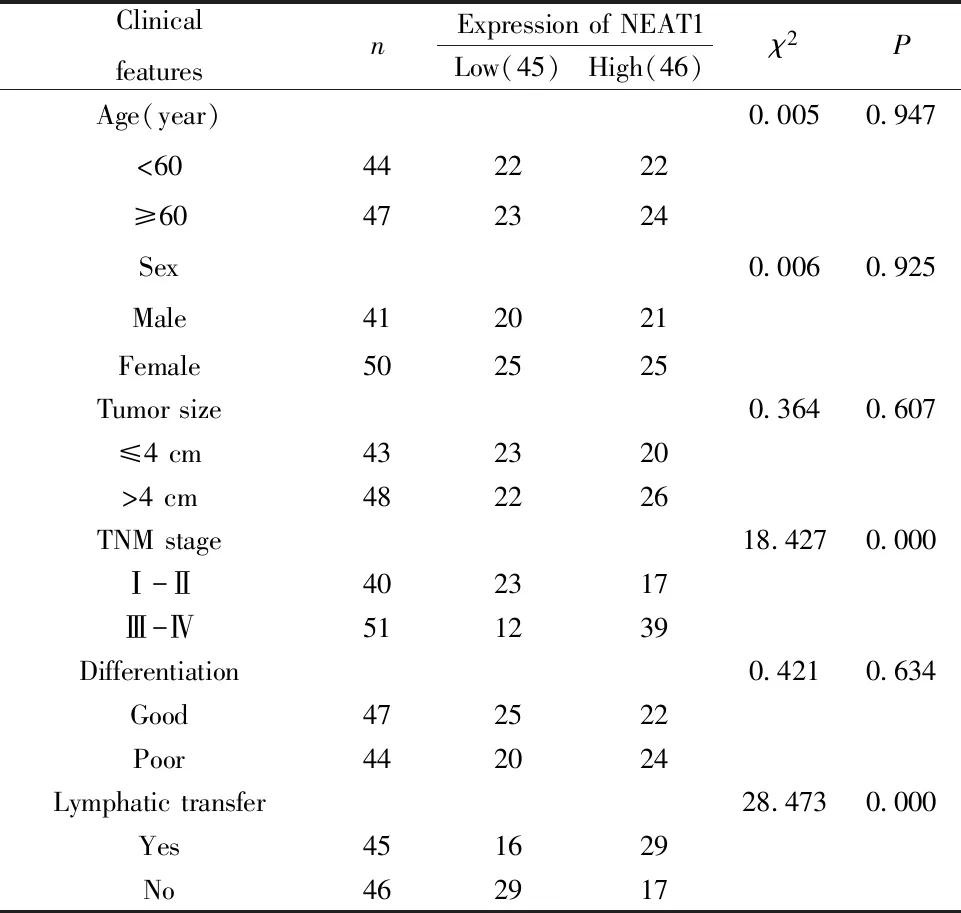
表2 NEAT1在NSCLC中的表達及與病理分期的關系(n=91)Tab.2 Relationship of expression of NSCLC with patho-logical stage(n=91)
2.2NEAT1沉默后抑制NSCLC細胞的增殖、遷移和浸潤 對si-NEAT1轉染細胞進行qRT-PCR檢測,NEAT1的表達量顯著下調(P<0.01,圖1A),MTT實驗也顯示A549和SPCA1轉染細胞與對照組相比NEAT1的表達受到抑制,NEAT1沉默組細胞的遷移和浸潤數量明顯減少,表明NEAT1沉默抑制了NSCLC細胞的代謝活性(圖1B)。

圖1 NEAT1沉默對NSCLC細胞代謝活性的影響Fig.1 Effect of NEAT1 silencing on metabolic activity of NSCLC cellsNote:A.The expression of NEAT1 after si-NEAT1 transfection;B.The metabolic activity of NSCLC cells after NEAT1 silenced.
2.3miR-101-3p的表達 通過檢測si-NEAT1轉染細胞A549和SPCA1中miR-101-3p的表達量發現,si-NEAT1轉染后miR-101-3p的表達顯著增高(P<0.05),然而miR-101-3p的表達上調并不影響NEAT1的表達(P>0.05),因此推斷miR-101-3p可能是NEAT1的抑制劑靶點(圖2)。

圖2 NEAT1與miR-101-3p的相關性Fig.2 Correlation of NEAT1 and miR-101-3pNote:A.The expression of miR-101-3p after si-NEAT1 transfection;B.The expression of NEAT1 after miR-101-3p overexpression.
2.4SOX9/Wnt/β-catenin信號通路與NSCLC的關系 應用Western斑點實驗檢測si-SOX9轉染A 549和SPCA1細胞系中的蛋白表達情況,結果如圖3所示,β-catenin和c-Myc的表達顯著降低,同時在miR-101-3p過量表達的細胞中SOX9、β-catenin和c-Myc的表達顯著降低,而在miR-101-3p受到抑制后其表達明顯升高,表明NEAT1/miR-101-3p/SOX9/Wnt/β-catenin信號通路在NSCLC發生發展中具有重要作用。
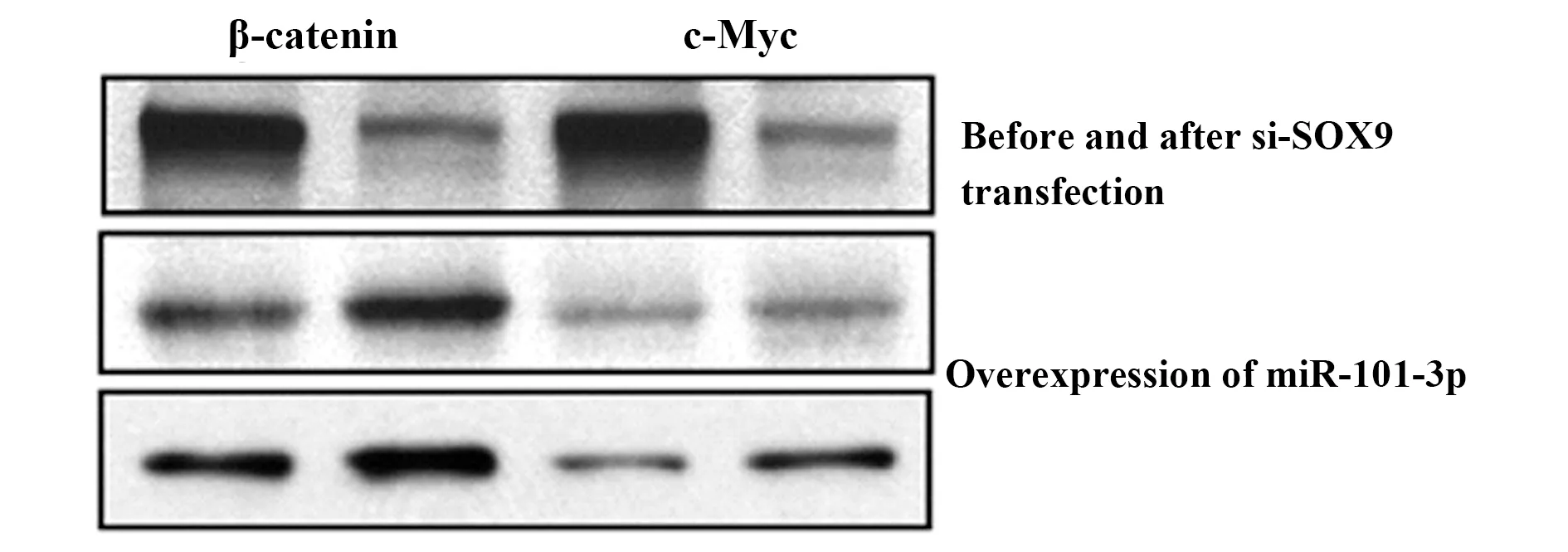
圖3 si-SOX9轉染及miR-101-3p過量表達前后β-catenin和c-Myc的表達情況Fig.3 Expression of β-catenin and c-Myc in NSCLC cells before and after si-SOX9 transfection and miR-101-3p overexpression
3 討論
近年來對于lncRNA的研究越來越多,大量證據表明其與癌癥的侵襲和轉移有關。Mang等[9]報道lncRNA NEAT1通過調節hnRNP A2的表達而促進肝癌的發生發展,且該調控與NEAT1-U2AF65蛋白復合物有關,可為肝癌的治療提供潛在靶點。Sun等[6]研究發現lncRNA NEAT1可作為has-miR-377-3p的競爭內源性RNA抑制其與E2F3的結合,是促進NSCLC進展的核心致癌基因,認為NEAT1作為內源性RNA調節E2F3的表達,建立起了調節性miRNA網絡與NSCLC發生發展的聯系。然而lncRNA NEAT1在NSCLC中的具體作用機制尚不清楚。
NEAT1在多種腫瘤中已有研究,邢宏松等[10]研究發現lncRNA NEAT1在肝癌細胞系中高表達,沉默lncRNA NEAT1表達可抑制肝癌細胞增殖、遷移和侵襲,其機制可能與調控miR-211/SATB2信號途徑有關。帥勇鋒等[11]研究發現lncRNA NEAT1在胃癌細胞株中高表達,lncRNA NEAT1沉默表達抑制胃癌增殖并誘導凋亡,可能與p-Akt、bcl-2下調bax上調有關。徐笑宇等[12]檢測宮頸病變患者血清中lncRNA NEAT1的表達水平發現,宮頸癌患者NEAT1表達水平低于健康對照組。潘翔等[13]檢測腎癌組織及其癌旁組織中NEAT1的表達量發現NEAT1在腎癌組織中表達明顯下調,并與癌組織類型、腎癌TNM分期和AJCC分期無明顯相關性。上述研究表明NEAT1在不同癌癥中分別發揮著促進和抑制的作用。
本研究發現NEAT1在NSCLC中發揮促進作用,其在NSCLC組織和細胞中均較正常對照呈高表達,且NEAT1的表達與NSCLC的臨床分期密切相關,NEAT1沉默能夠抑制NSCLC細胞的增殖、遷移和浸潤。既往研究表明lncRNA能夠調控miRNA的表達和活性,Lo等[14]發現BRCA1/NEAT1/miR-129-5p/WNT4信號通路在乳腺癌中發揮致癌作用。Wang等[15]發現lncRNA NEAT1通過調節miR-107/CDK6途徑促進喉鱗狀細胞癌的發生發展,認為NEAT1可作為其治療的潛在靶點。然而lncRNA和miRNA在NSCLC中的研究文獻還很少,尚需大量的研究予以證實。本研究中,我們通過證實了miR-101-3p為NEAT1的抑制靶標,下調NEAT1的表達能夠增加miR-101-3p的表達,但是miR-101-3p表達增高并不能影響NEAT1的表達,因此認為miR-101-3p是NEAT1的直接靶標。
Wnt/β-catenin信號通路能夠通過調節多功能β-catenin蛋白活性而在多種細胞間發揮重要作用,近來研究發現在腫瘤進展中Wnt/β-catenin信號通路與SOX9間具有顯著的相關性,比如Santos等[16]發現SOX9升高后作用于Wnt信號傳導驅動胃癌進展,Ma等[17]發現SOX9驅動前列腺癌中的Wnt信號通路,Prévostel等[18]發現SOX9是一種控制Wnt/β-catenin的非典型腸癌抑制因子。本研究發現SOX9可以作為miR-101-3p的靶標,NEAT1通過激活NEAT1/miR-101-3p/SOX9/Wnt/β-catenin軸在NSCLC進展中發揮致癌作用,因此認為NEAT1有望成為NSCLC基因治療的新靶標。
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
電子制作(2018年11期)2018-08-04 03:25:42
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25