小麥雄性不育系BS366和FA-101的配合力及雜種優勢分析
2020-01-05 14:06:20苑少華李艷梅白建芳段文靜譚照國劉子涵趙昌平張風廷陳兆波張立平
麥類作物學報 2020年12期
苑少華,李艷梅,白建芳,段文靜,譚照國,3,劉子涵,王 娜,趙昌平,張風廷,陳兆波,張立平
(1. 北京市農林科學院雜交小麥工程技術研究中心,北京 100097;2. 雜交小麥分子遺傳北京市重點實驗室,北京 100097;3. 長江大學生命科學學院,湖北荊州 434025)
小麥是我國重要的糧食作物之一。隨著社會經濟的發展以及人口的快速增長,小麥的需求量也呈增加趨勢。在耕地面積不斷減少的形勢下,小麥總產量的提升只能通過單產的大幅度提高來實現[1]。雜種優勢利用是提高作物產量的有效途徑之一[2],而小麥還未大規模實現雜種優勢利用[3]。近年來,我國基于小麥光溫敏不育系創建了“二系法雜交小麥應用技術體系”,由于其操作簡單、恢復源較廣、制種成本低等優勢,該雜優利用體系成為雜交小麥應用的主要途徑[4]。在生產中應用的BS型小麥光溫敏不育系為“京麥”系列雜交小麥品種的母本,其新一代不育系的光敏特性顯著,大幅提高了不育系的穩定性。F型三系不育系材料是我國新發現的不育源,是一種新型普通小麥細胞質雄性不育系[5],而且其育性具有一定的光溫敏感特性[6]。
小麥雜種優勢大面積利用中存在諸多制約因素,其中強優勢組合的選配是關鍵[7]。強優勢組合的選配主要是通過親本配合力分析對雜交種進行親本選擇和組合評價[8]。小麥產量構成的有效穗數、穗粒數和千粒重三要素中,千粒重的遺傳力最大[9-10]。通過分析不育系及恢復系主要農藝性狀的配合力,對篩選強優勢雜交組合、提高育種效率具有非常重要的意義。前人對于雜交小麥雜種優勢利用及配合力分析也有報道,但尚無針對兩系或F型雄性不育系的配合力及雜種優勢比較的相關研究。前期李曉燕[11]對光溫敏二系小麥BS210和BS366的雜種優勢和配合力進行了評價,陳 林[12]、溫輝芹[13]和任永康[14]等僅對F型不育系雜種優勢進行了評價,但并未對配合力進行深入分析。本研究利用光溫敏雄性不育系BS366和F型雄性不育系FA-101及其雜交組合對其產量性狀的雜種優勢和配合力進行分析,探索強優勢組合的選配規律,以期對二系及三系雜交小麥強優勢組合的組配提供理論 支持。
1 材料和方法
1.1 試驗材料
供試材料為兩系光溫敏核雄性不育系BS366、F型三系雄性不育系FA-101及16個恢復系材料(川麥56、濟麥12、綿陽35、川育21、鳳麥24、07品151-180、川麥43、川農16、蜀麥482、川麥52、Syn53、太原608、PI601477、PI17901、PI486146和PI598210,分別用P1~P16表示)。
1.2 試驗方法
2018年4月以BS366和FA-101為母本,與16個父本材料進行雜交,2018年9月下旬將雜交收獲的F0代材料種植于北京(N 39°54′, E 116°18′)生態區,隨機區組設計,三次重復,兩行區, 1.5 m行長,行距25 cm,株距5 cm,人工點播,田間管理同大田生產。2019年5月對親本、F1代株高、穗長、單株穗數、小穗數、穗粒數、千粒重和單株粒重進行調查。
單株粒重=單株穗數×穗粒數×千粒重/ 1 000
1.3 數據分析和處理
配合力分析采用不完全雙列法雜交[15],各供試親本的一般配合力(GCA)按gi=xi-X計算;各參試組合的特殊配合力(SCA)按sij=xij-X-gi-gj計算。其中xi和xj分別表示i親本和j親本所配組合的性狀平均值,X為所有組合的總平均值。對調查的親本7個性狀值以及7個性狀的GCA和SCA在DPS軟件下進行分析。
超中親優勢=(F1-PA)/PA×100%,PA為雙親平均值;超親優勢=(F1-P)/P×100%,P為較優親本值;對照優勢=(F1-CK)/CK×100%,CK為對照值。
2 結果與分析
2.1 親本及雜交組合的配合力分析
2.1.1 雜交組合配合力方差分析
對具有的7個相同父本(P1、P3、P5、P9、P12、P14和P15)的14個雜交組合的7個性狀進行方差分析表明,各性狀在組合間均存在極顯著差異(P<0.01)或顯著差異(P<0.05),而區組間除株高和穗長外均不顯著;父本間株高的GCA值差異顯著,母本間株高和小穂數GCA值差異極顯著,其余性狀均不顯著;組合間各性狀SCA值均極顯著(表1),因此可進一步分析親本的GCA和組合的SCA效應。
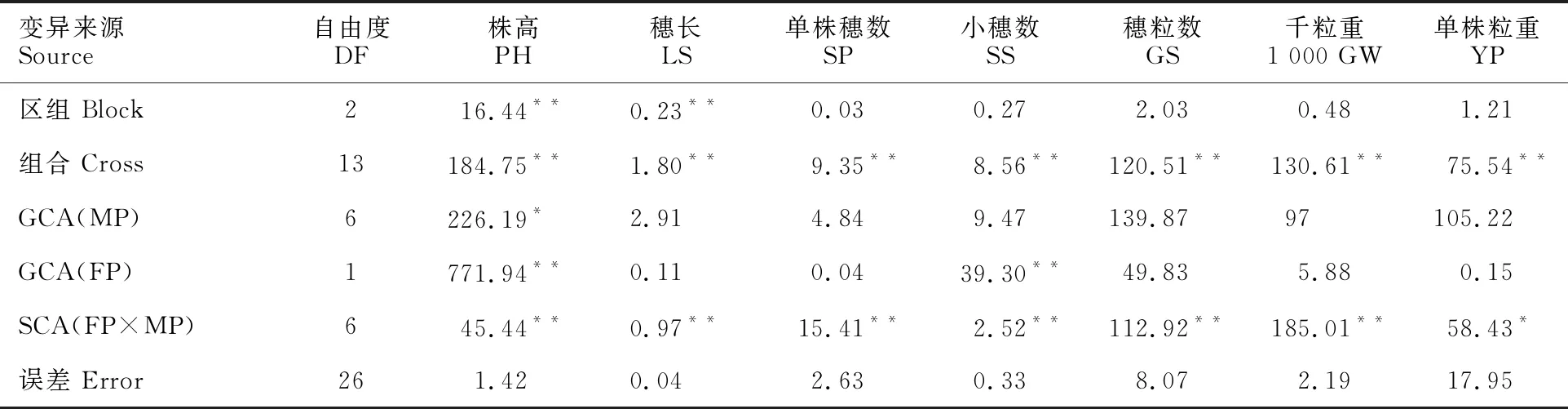
表1 7個性狀的方差分析(F值)
2.1.2 親本的GCA效應分析
GCA指一個親本與多個親本雜交后雜種后代在某個性狀上的平均表現,主要由基因的加性效應決定,是可以穩定遺傳的部分[16]。從表2可以看出,同一親本的不同性狀間和同一性狀的不同親本間GCA值都有較大差異,說明不同親本在性狀上的加性基因效應值大小不同。
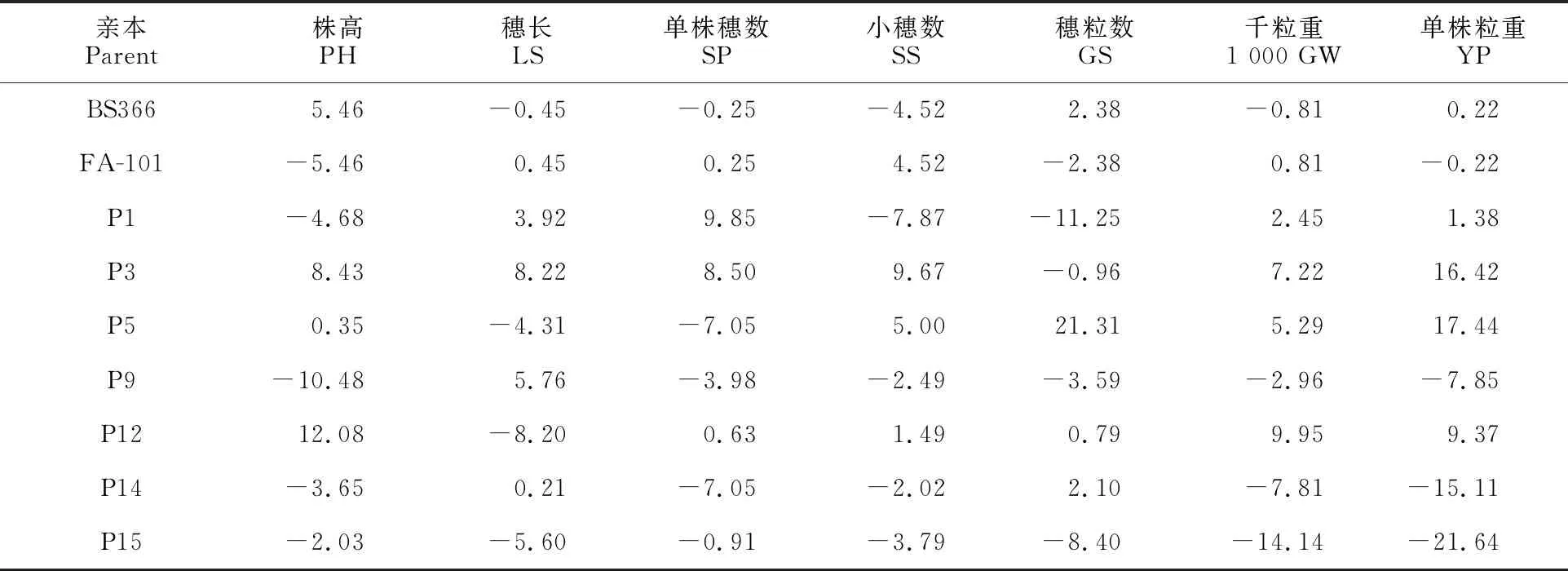
表2 9個小麥親本7個性狀的一般配合力(GCA)效應值
在7個父本中,P9、P1、P14和P15的株高GCA為負值,相對效應較大,說明這4個材料在矮稈育種中有一定的利用價值;穗長GCA值較大的有P3、P9和P1;小穗數GCA值較大的有P3和P5;單株粒重GCA值較大的有P5、P3和P12。此外,在單株粒重GCA值為正值的P1、P3、P5和P12的4個親本中,在單株穗數、穗粒數、千粒重等3個產量性狀中均至少具有2個或2個以上性狀存在GCA正向效應值(表3),表明本研究中影響產量的最終因素并不是某單一性狀特別突出而造成的,而是產量要素共同作用的結果。
2.1.3 雜交組合產量三要素和單株粒重的SCA效應分析
SCA是由非加性效應產生的,很難在后代中穩定遺傳,但親本間的特殊配合力效應大,有利于選育出該性狀表現優異的突破性雜交小麥組合。從表4中可以看出,不同組合單株粒重的SCA值有很大差異,其變化范圍為-21.99~21.99。其中SCA值較大的組合有BS/P9、FA/P1、FA/P12和FA/P15,其單株穗數、穗粒數和千粒重中至少有2個或2個以上性狀的SCA值表現出正向效應;SCA值較小的組合有FA/P9、BS/P1、BS/P12和BS/P15(表3)。

表3 14個雜交組合7個性狀的特殊配合力(SCA)效應值及其多重比較分析
2.2 雜交組合的雜種優勢分析
從23個雜交組合F1代的雜種優勢(表4)看,7個性狀超中親優勢組合比例為56.52%~100%,超中親優勢明顯,且7個性狀的超中親優勢均值均為正;超高親優勢組合比例為17.39%~65.22%,穗長、單株穗數、千粒重和單株粒重的超高親優勢均值為正;超對照優勢組合比例為30.43%~95.65%,穗長、單株穗數和小穗數的超對照優勢均值為正。
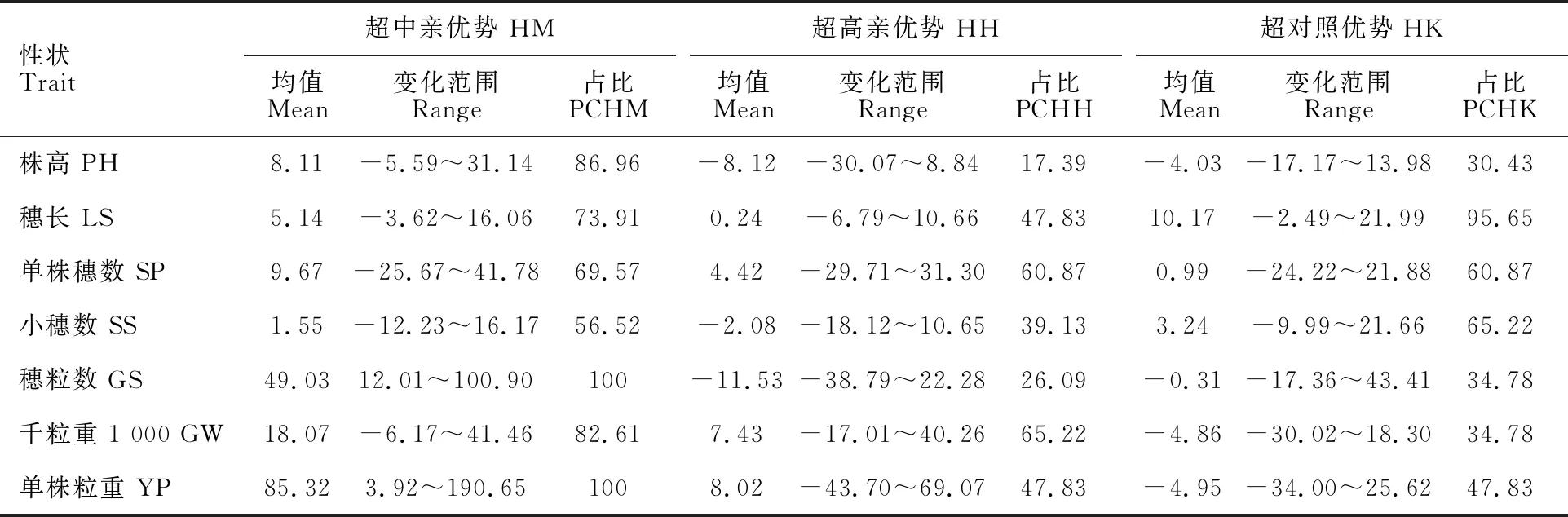
表4 雜交組合F1代各農藝性狀的中親優勢和超親優勢
從23個雜交組合中篩選出11個組合(6個BS366組合,5個FA-101組合),其單株粒重超對照優勢為正,范圍為2.73%~25.62%,株高超對照優勢為-13.53%~10.31%(表5)。其中BS/P2、BS/P5、BS/P6和FA/P4等4個組合的單株粒重超對照優勢均大于12%,且株高均低于 對照。
此外,除BS/P2、BS/P5、BS/P7和FA/P1以外的7個組合在單株穗數、穗粒數、千粒重等3個產量性狀中至少有2個或2個以上的雜種優勢為正,表明本研究中影響產量的最終因素并不是某單一性狀特別突出而造成的,大多是產量要素共同作用的結果。
另外發現,父本P3和P5組合的單株粒重均超對照(表5),因此這2個親本可能作為優良父本與其他不育系進行組合選配,進而發掘新的具有雜種優勢的組合。
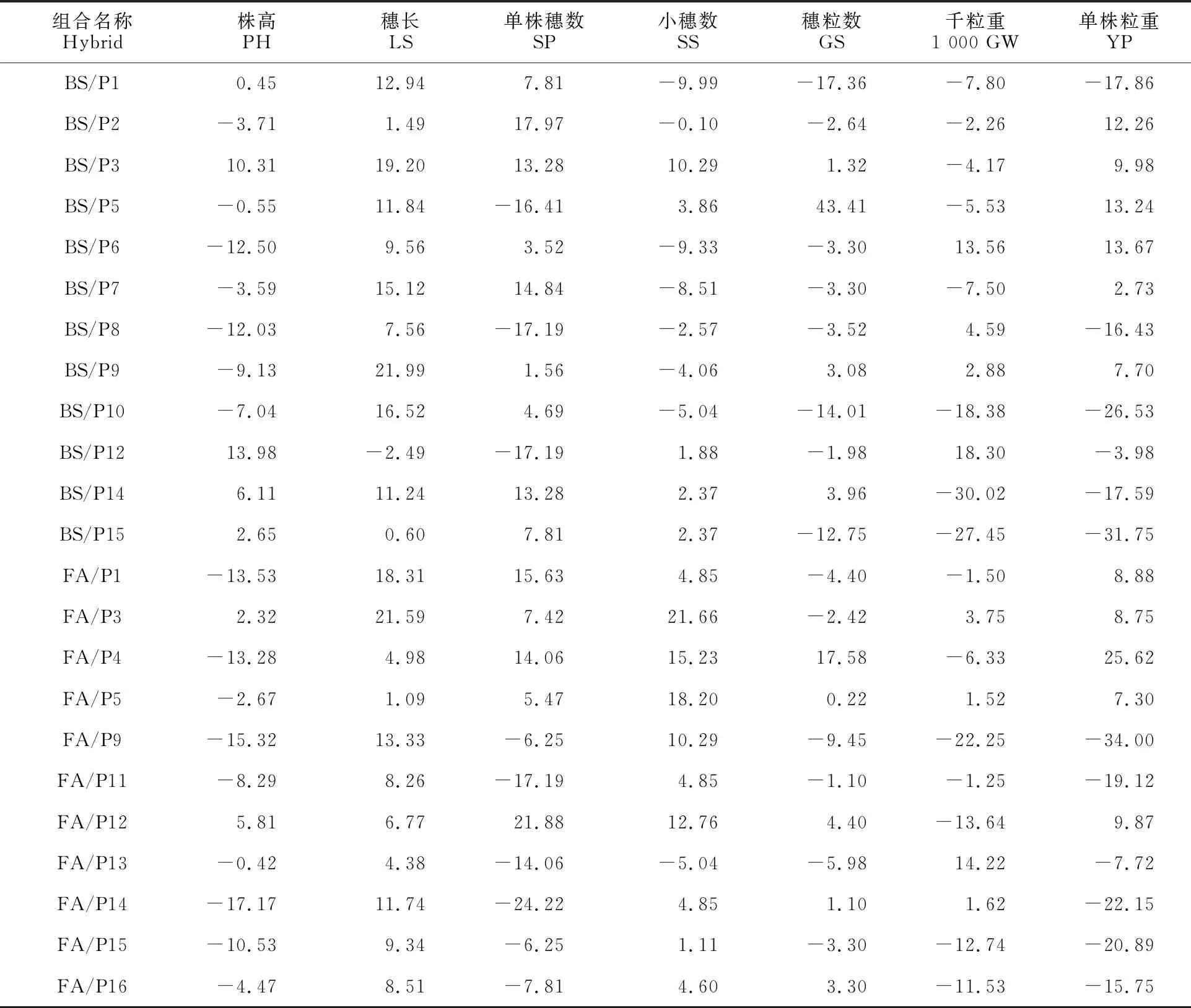
表5 7個農藝性狀的超對照優勢
2.3 雜種優勢與配合力的相關性分析
在小麥雜交育種中,雜種優勢與配合力有著密不可分的關系,具有良好配合力的親本或品系才能組配出更優異的雜交種,因而親本配合力是選擇優良雜交種的重要依據[16]。一個雜交組合的優劣由親本的GCA和組合的SCA效應共同決定[17]。目前,產量是評價雜種優勢的最主要標準之一[18]。
對14個雜交組合的雜種超對照優勢值與父本GCA值和組合SCA值的相關性分析表明,雜種優勢值與GCA和SCA都有較高的相關性,且與GCA的相關性要大于與SCA的相關性,但父本GCA值與組合SCA值幾乎無相關性(表6)。

表6 雜種優勢與GCA、SCA的相關性
2.4 BS不育系與FA不育系配合力及后代雜種優勢比較分析
2.4.1 BS366和FA-101配合力比較分析
BS366和FA-101在株高、小穗數和穗粒數等3個性狀上的GCA差異明顯(表2)。BS366單株穗數和千粒重的GCA值低于FA-101,但單株粒重要優于FA-101。因此,在雜交小麥新組合的選育上,應根據不育系的特點,定向選擇可以優勢互補的恢復系進行組配,有利于獲得產量水平較高的強優勢組合。
在單株穗數、穗粒數、千粒重和單株粒重等產量相關性狀中,相同父本的BS366和FA-101的7對組合SCA值之差的范圍分別是6.26~37.92、1.05~38.24、5.15~35.94和1.17~43.99,而且每個性狀下均有4對組合的SCA值之差大于10(表3),說明這2種不育系的相同父本組合大多數情況下的特殊配合力有較大差異。
2.4.2 BS366和FA-101雜種優勢比較分析
相同父本情況下分別對BS366和FA-101的所有組合的雜種優勢進行比較(表5)。結果發現,BS366和FA-101分別與P3、P5、P14和P15的雜交組合的超對照優勢比較一致,均為正或負。此外,兩個不育系與P1、P9和P12的雜交組合的超對照優勢值的正負完全相反,而其余9個父本的組合中均只有1個組合為高恢復性組合(結實率>90%),因此無法進行對比,表明這兩個不育系的組合的雜種優勢和恢復性存在明顯不同。
2.4.3 BS366和FA-101雜種優勢相關性分析
對上述相同父本的BS366和FA-101的7個雜交組合間的7個性狀的雜種優勢進行相關性分析發現,只有株高間的相關性大于0.6,具有較強的相關性;穗長、小穂數、穗粒數和株粒重只具有中度或中度以下的相關性;而單株穗數和千粒重具有負相關性(表7)。此外,分別對BS和FA組合的單株粒重與產量三要素的相關性分析發現,BS366組合中單株粒重與穗粒數和千粒重具有較強的相關性,與單株穗數具有一定的負相關性;FA-101組合中與單株穗數具有極強相關性,與穗粒數和千粒重具有一定的相關性(表8)。因此,BS366和FA-101兩種不育系配制的雜交組合,其雜種優勢趨勢不同,尤其是在單株穗數、穗粒數、千粒重和單株粒重等產量性狀方面存在較大差異(表7、表8)。

表7 具有相同父本的BS366和FA-101組合的各農藝性狀相關性

表8 BS366組合和FA-101組合的株粒重 與產量三要素的相關性
3 討 論
3.1 BS及F型不育系優良雜交組合及父本的 篩選
本研究結果表明,影響BS366組合產量的性狀順序為穗粒數、千粒重>單株穗數,影響FA-101組合的順序為單株穗數>穗粒數、千粒重(表8),兩者結果不同,與前人研究結論存在偏差[18-20],其原因應該是試驗材料不同導致的差異。同時發現穗粒數的超高親和超對照優勢均為負,且超高親優勢組合僅占26.09%(表4),在所有產量性狀里最低,說明不育系育性恢復性偏低影響結實率是二系雜交小麥產量提高的瓶頸。因此,應特別重視對恢復系的選擇,進而大幅度提高穗粒數的超親(或超標)優勢,是超高產雜交小麥組合選育的關鍵,也是難點之一。
本研究通過對雜種優勢的分析,篩選出BS/P2、BS/P5、BS/P6和FA/P4等4個高產雜交小麥新組合,其是否能進行生產應用,還需要從雙親花期、株高、制種產量等方面進行試驗;父本P3和P5可作為優良核心恢復系使用。
3.2 BS366與FA-101的組合的比較分析
總體上講,小麥雜種優勢利用相對落后,經過幾代人努力目前已取得了較大的成績,但是關于小麥光溫敏兩系和F型三系不育系利用相同父本配制雜交組合,并比較其雜種優勢和配合力的研究鮮有報道。前期苑少華、孫輝等[6,21]對光溫敏二系不育系BS366和F型三系不育系的恢復性研究發現,二者都具有不同程度的光溫敏特性,但其恢復源不完全一致,其不育及育性恢復機制存在一定差異。本研究發現,BS366和FA-101兩個不育系各性狀的一般配合力不同(表2),具有相同父本的BS和FA組合間的雜種優勢趨勢、規律(表5、表7、表8)以及特殊配合力效應值(表3)也大多存在差異,進一步驗證了BS和FA屬于不同類型的不育系,而且兩者提高雜種優勢潛力的策略有所不同。
