雙免耕覆蓋對旱地土壤微生物、酶活性和冬小麥干物質轉運的影響
2020-01-05 14:03:18邵運輝李俊紅岳俊芹丁志強張德奇方保停李向東
麥類作物學報 2020年12期
邵運輝,李俊紅,岳俊芹,丁志強,張德奇,方保停,楊 程,李向東
(1.河南省農業科學院小麥研究所,河南鄭州 450002; 2.洛陽農林科學院,河南洛陽 471023; 3.中國農業科學院洛陽旱農試驗基地,河南洛陽 471023)
在豫西旱區,干旱少雨且水土流失嚴重,自然降水是作物生產的主要水資源[1]。保護性耕作主要指秸稈還田、免耕播種和殘茬覆蓋等耕作手段,可有效增加土壤有機質含量,調節土壤的C/N、溫度和水分變化,對作物有一定的增產效果[2]。免耕等保護性耕作基本上不擾動土層,秸稈和根茬等有機物腐解后的殘茬和根系分泌物主要積累在表層土壤中,為土壤微生物的生命活動提供了足夠的能源[3]。土壤微生物通過分解動植物殘體參與生態系統的能量流動和物質循環,其數量是評價土壤肥力的重要指標之一。土壤酶則主要來源于活體植物、死亡植株殘體,直接參與植物與土壤之間的物質循環和土壤中生物的生化反應,可提高土壤養分的可用性,其活性大小直接影響土壤肥力[4]。保護性耕作可增加和維護土壤微生物種類和數量,提高土壤酶的活性,改善作物根區環境,提高作物抗逆性,促進作物高產、穩產,并提高土壤質量和改善生態環境[5-6]。土壤微生物和土壤酶共同影響并推動著土壤的代謝過程,二者的活性關系密切[7]。土壤酶活性、土壤微生物的代謝和種類與植物根系生長狀況有一定關系。林雁冰等[6]認為,冬小麥根系生理代謝對根區土壤微生物數量和種類有顯著影響,覆蓋可以提高土壤細菌、真菌和放線菌的數量。土壤酶活性高,則小麥根系衰老較慢,冬小麥根系的生長狀況直接影響地上部生長狀況和抗旱能力,最終影響其產量[8-11]。冬小麥產量的物質來源主要有兩個方面,一是花后光合同化物的直接輸入,另一個是花前貯藏在營養器官的光合同化物的再運轉。王征宏等[9]研究發現,干旱對冬小麥干物質轉運有較大的影響,灌漿前期干旱會導致花前貯藏物質向籽粒的轉運量降低。
目前,國內外對保護性耕作下土壤微生物、酶活性及干物質積累等方面研究有較多的報道,但有關旱作區雙免耕覆蓋對冬小麥土壤微生物和酶活等的影響尚未見報道。鑒于此,本研究擬對雙免耕覆蓋下旱地冬小麥田土壤微生物、酶活及冬小麥花前、花后干物質積累和轉運進行深入研究,旨在為旱作區冬小麥增產提供理論依據,為選擇合理有效的保護性耕作模式提供參考。
1 材料和方法
1.1 試驗地概況
試驗設在洛陽農科院旱農基地防滲精確水分池,土壤為潮褐土,質地中壤,耕層容重1.53 g·cm-3,田間相對持水量23.48%,飽和含水量 33.36%。該區域年均輻射量491.5 kJ·cm-2,年均氣溫14 ℃,日平均溫度超過10 ℃的天數約210 d,大于10 ℃的積溫可達4 000 ℃,年蒸發量 1 841.7 mm。土壤耕層有機質為15.8 g·kg-1,堿解氮(N)為62.7 mg·kg-1,速效磷(P2O5)為10.39 mg·kg-1,速效鉀(K2O)為166.0 mg·kg-1。種植模式為冬小麥、玉米一年兩 熟制。
1.2 試驗設計
本研究于2013年10月—2014年6月進行。設2個處理:(1)雙免耕覆蓋(NT),冬小麥收獲時留茬35~40 cm,秸稈脫粒后還田并覆蓋于地表,不翻耕土壤,鐵茬播種玉米;玉米收獲后留根茬,其秸稈覆蓋于冬小麥行間,冬小麥采用人工開溝種植,盡量少擾動土壤,等行播種;(2)傳統耕作(CK),即深翻耕25~30 cm(冬小麥、玉米秸稈均不還田)。小區面積為4 m×4 m。供試冬小麥品種為洛旱7號,種植密度13.5×104株·hm-2。玉米品種為洛玉7號,種植密度為4.5×104株·hm-2。
1.3 測定項目與方法
1.3.1 土樣采集
在冬小麥苗期(seedling stage,SS)、越冬期(over winter stage,OS)、拔節期(jointing stage,JS)、灌漿期(filling stage,FS)、成熟期(maturation stage,MS)的晴天上午10:00左右,每個小區按五點法取土樣,每點取土深度為0~60 cm,每10 cm取一次,剔除石塊、植物殘根、蚯蚓等雜物后裝袋,樣品風干后經研磨,過1 mm篩,采用四分法選取待測樣品。
1.3.2 土壤相關酶活性測定[7]
土壤脲酶活性采用苯酚鈉比色法測定;土壤蛋白酶活性采用茚三酮比色法測定。
1.3.3 土壤微生物數量測定[4]
細菌采用牛肉膏蛋白胨培養基平板計數法測定;真菌采用馬丁氏培養基平板計數法測定;放線菌采用淀粉銨鹽培養基平板計數法測定。
1.3.4 冬小麥干物質積累相關指標測定
在冬小麥開花期和成熟期,每個小區隨機取10株具代表性植株地上部,開花期取樣后將植株分為葉片、莖桿+葉鞘和穗三部分,成熟期取樣后將植株分為葉片、莖稈+葉鞘、穎殼+穗軸、籽粒四部分。將樣品于105 ℃殺青15 min,65 ℃烘干至恒重,測定干物質積累量。相關指標計算公式如下[12]:
花前同化物轉運量=開花期植株干重-成熟期營養器官干重
(1)
花前同化物轉運效率=花前同化物轉運量/開花期植株干重×100%
(2)
花前同化物對籽粒產量貢獻率=花前同化物轉運量/成熟期籽粒干重×100%
(3)
花后同化物積累量=成熟期籽粒干重-花前同化物轉運量
(4)
花后同化物對籽粒產量貢獻率=花后同化物積累量/成熟期籽粒干重×100%
(5)
1.3.5 產量及其構成因素測定
在冬小麥收獲期,按一般方法調查穗數、穗粒數,測定千粒重,整區收獲計產。
1.4 數據處理
采用Excel 2010 和DPS 7.05進行數據處理與統計分析,用LSD法進行差異顯著性檢驗。
2 結果與分析
2.1 雙免耕覆蓋對麥田土壤中微生物數量的 影響
2.1.1 對麥田土壤中細菌數量的影響
從圖1可知,隨著冬小麥生育進程的推進,不同土層的細菌數量呈現先上升后下降的趨勢;以越冬期最少,拔節期最多,拔節期以后逐漸減少。雙免耕覆蓋下0~10 cm土層細菌數量在冬小麥不同生育時期均顯著高于傳統耕作(圖1A),尤其是成熟前,二者差異達極顯著水平,雙免耕覆蓋下細菌數量平均較傳統耕作高94.7%。雙免耕覆蓋下10~20 cm土層冬小麥不同生育時期細菌數量在苗期和越冬期顯著高于傳統耕作外,之后二者差異達極顯著水平(圖1B),雙免耕下細菌數量平均較傳統耕作高53.3%。這種現象可能與秸稈覆蓋能保存較充足的土壤水分和維持適宜的土壤溫度有較大關系。
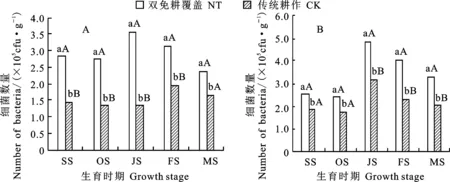
相同時期圖柱上不同小寫字母和大寫字母分別表示處理間差異達5%和1%顯著水平。下同。
2.1.2 對麥田土壤中真菌數量的影響
土壤中的真菌參與土壤中有機質的分解,在土壤腐殖質的合成、氨化作用以及團聚體的形成等過程中發揮著重要的作用,可直接影響土壤肥力[6]。 由圖2可知,0~20 cm土層土壤真菌數量在越冬期最低,拔節期和灌漿期較高;雙免耕覆蓋下麥田土壤真菌數量均不同程度高于傳統耕作。0~10 cm土層,雙免耕處理土壤中真菌數量均顯著或極顯著高于傳統耕作(圖2A),拔節期和灌漿期分別較傳統耕作高23.4%和34.1%。除拔節期外雙免耕覆蓋處理下10~20 cm土層土壤 中真菌數量均顯著或極顯著高于傳統耕作 (圖2B)。

圖2 兩種耕作模式對麥田不同生育時期土壤真菌數量的影響
2.1.3 對麥田土壤中放線菌數量的影響
放線菌能分解土壤腐殖質中最穩定的有機化合物,提高土壤肥力。從圖3可以看出,0~20 cm土層土壤中的放線菌數量以苗期最高,0~10 cm(圖3A)和10~20 cm土層(圖3B),雙免耕覆蓋較傳統耕作分別高28.5%和74.7%,差異均達極顯著水平;越冬期土壤中放線菌的數量最少。從越冬期至成熟期,雙免耕覆蓋0~20 cm土層中放線菌數量有不同程度的增加;不同時期雙免耕覆蓋土壤中放線菌數量均顯著高于傳統耕作。
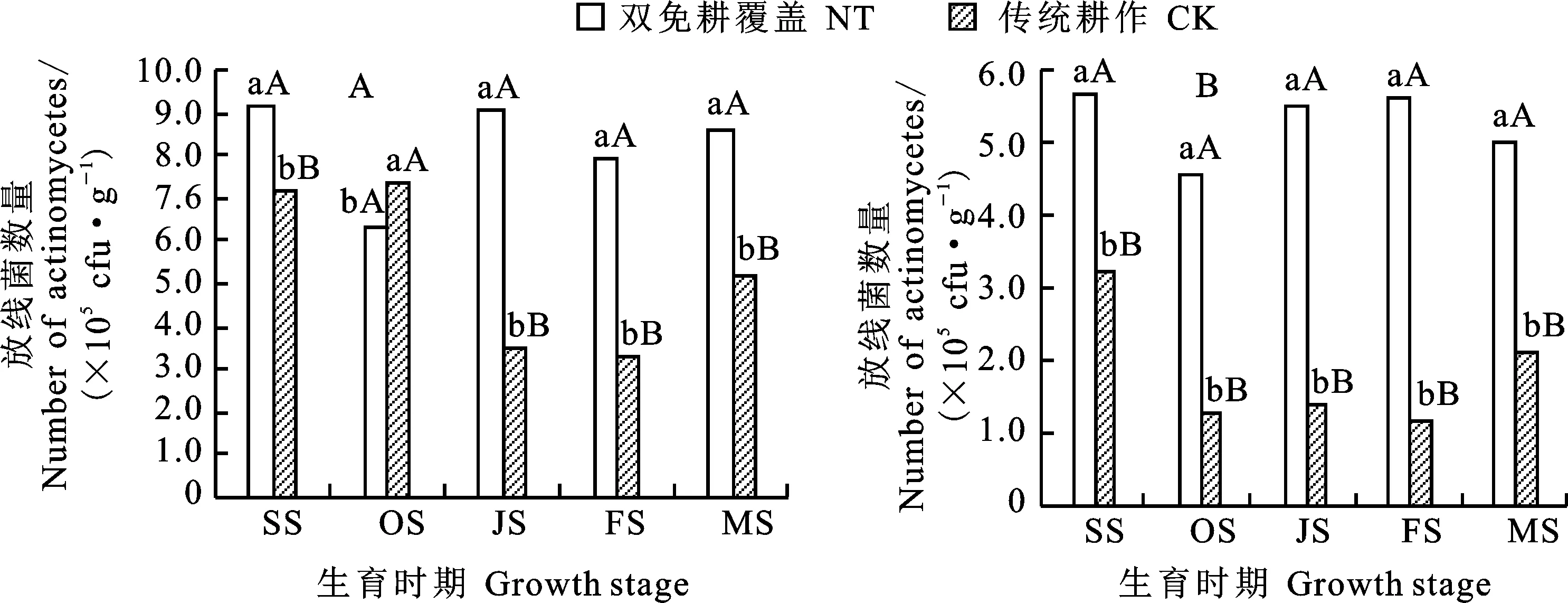
圖3 兩種耕作模式對麥田不同生育時期土壤中放線菌數量的影響
2.1.4 雙免耕覆蓋下麥田土壤中微生物總量的垂直分布
由圖4可知,雙免耕覆蓋和傳統耕作下,冬小麥土壤中微生物總量(細菌、真菌和放線菌數量之和)隨土層深度的增加均呈現逐漸下降趨勢。雙免耕覆蓋下冬小麥土壤微生物總量在不同土層均極顯著高于傳統耕作,0~60 cm土層較傳統耕作平均高 68.2%。雙免耕覆蓋下冬小麥土壤0~40 cm土層中微生物總量迅速遞減,而傳統耕作則在0~20 cm土層呈現迅速遞減趨勢。
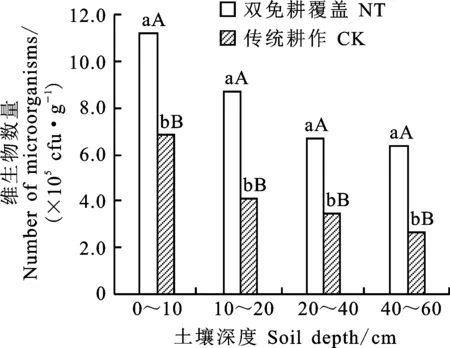
圖4 兩種耕作模式對麥田土壤微生物總量垂直分布的影響
2.2 雙免耕覆蓋對麥田土壤酶活性的影響
2.2.1 對冬小麥不同生育期土壤酶活性的影響
從圖5可知,冬小麥不同生育時期0~20 cm土層土壤中蛋白酶活性的變化幅度較為緩和(圖5A),脲酶活性在越冬期最低(圖5B),其余生育時期的變化與蛋白酶的活性變化表現一致,均隨生育進程的推進逐漸減小。雙免耕覆蓋下,冬小麥不同生育期土壤中的兩種酶活性均高于傳統耕作。在冬小麥全生育期內,雙免耕覆蓋下0~20 cm土層土壤中蛋白酶活性較傳統耕作平均增加了33.6%,差異達極顯著水平(P<0.01)。土壤中蛋白酶活性以苗期最高,較傳統耕作增加了15.0%,拔節期次之,成熟期最低,成熟期雙免耕覆蓋仍較傳統耕作高27.6%(圖5A)。
雙免耕覆蓋下土壤中脲酶活性在冬小麥全生育期較傳統耕作平均高28.6%,除成熟期外,其他生育期二者差異均達極顯著水平(圖5B)。土壤脲酶活性在冬小麥苗期最大,雙免耕覆蓋較傳統耕作高28.8%。越冬期脲酶活性急劇下降,在冬小麥整個生育期中為最低,此期雙免耕覆蓋較傳統耕作高55.3%。這可能因為雙免耕覆蓋有利于提高地溫,從而有利于提高土壤酶活性。拔節期土壤脲酶活性逐漸上升,雙免耕覆蓋較傳統耕作高28.6%,灌漿期又迅速下降,成熟期二者差異不顯著。

圖5 兩種耕作模式對冬小麥不同生育時期土壤酶活性的影響(0~20 cm)
2.2.2 雙免耕覆蓋下麥田土壤中酶活性的垂直分布
從表1可以看出,在0~60 cm土層,雙免耕覆蓋下麥田土壤中蛋白酶和脲酶的活性均隨著土層深度的增加逐漸降低,蛋白酶和脲酶活性分別較傳統耕作平均增加了27.3%和32.7%。雙免耕覆蓋下蛋白酶活性在0~10 cm和40~60 cm分別較傳統耕作高31.5%和37.7%,脲酶活性在10~20 cm和40~60 cm土層均較傳統耕作高35.9%和36.4%,差異均達極顯著水平。這表明在干旱條件下,雙免耕覆蓋有利于提高耕層土壤的酶活性。
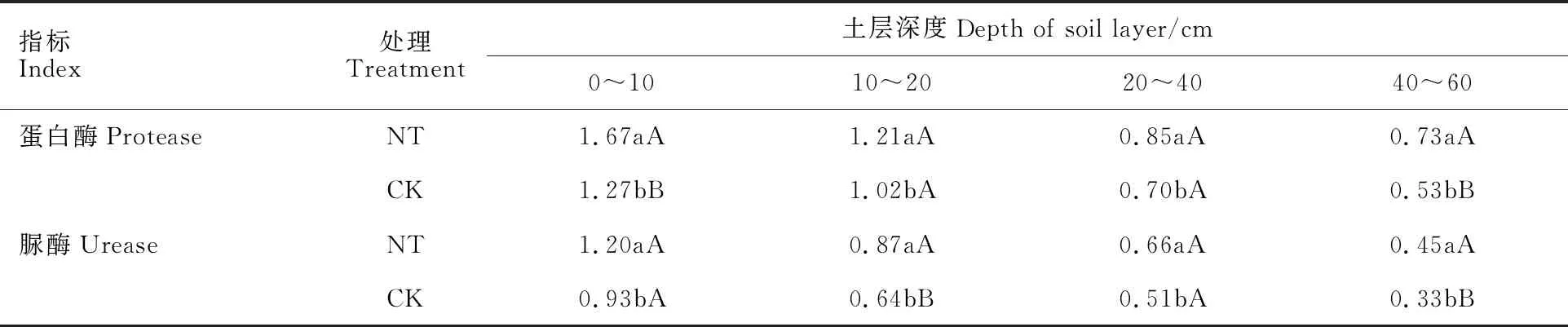
表1 兩種耕作模式下麥田土壤酶活性分布
2.3 雙免耕覆蓋對冬小麥開花前后干物質積累和轉運的影響
灌漿期是冬小麥籽粒干物質積累的主要時期,大約有70%~90%的籽粒干物質來自于灌漿期的光合同化物[13]。從表2可以看出,雙免耕覆蓋下,冬小麥較傳統耕作增產18.4%,差異極顯著;花后干物質積累量及其對籽粒產量的貢獻率均極顯著高于傳統耕作;花后干物質積累量對籽粒產量的貢獻率為44.2%,較傳統耕作高 61.9%。傳統耕作冬小麥花前干物質積累量對籽粒產量的貢獻率為72.7%,顯著高于雙免耕覆蓋,但干物質轉運效率顯著低于雙免耕覆蓋。整體而言,冬小麥籽粒干物質主要源自開花前干物質的轉運,雙免耕覆蓋較傳統耕作提高了旱地冬小麥開花后干物質積累量及其對籽粒產量的貢獻率,為雙免耕覆蓋下冬小麥獲得高產奠定了基礎。

表2 旱作區兩種耕作模式對冬小麥開花前后干物質積累和轉運的影響
3 討論與結論
3.1 雙免耕覆蓋對冬小麥土壤微生物的影響
本研究發現,雙免耕覆蓋下冬小麥土壤中細菌、真菌和放線菌的數量均顯著高于傳統耕作,這與趙亞麗等[2]、張星杰等[3]和王 靜等[5]的研究結果基本一致,但土壤中微生物數量的多少和種類是否受秸稈還田量及還田年限的影響還需進一步研究。免耕覆蓋可提高微生物增長的空間,可明顯提高微生物種群數量,尤其是耕層微生物更為活躍[13-16]。隨土壤深度的增加,微生物數量逐漸減少,這可能與土壤含水量、溫度等條件的變化有關。雙免耕覆蓋下,微生物數量的變化在冬小麥各生育期不同深度土層的表現不完全一致,細菌數量以拔節期最多,真菌數量以拔節期和灌漿期較多,放線菌數量以苗期最多,這可能與冬小麥根系生長狀況、土壤含水量、溫度及微生物生長所需環境等有關。
3.2 雙免耕覆蓋對冬小麥土壤酶活性的影響
雙免耕覆蓋下冬小麥土壤中蛋白酶和脲酶的活性均隨著土層深度的增加逐漸降低,雙免耕覆蓋下冬小麥土壤中酶活性均極顯著高于傳統耕作,這是因為在豫西旱作區,干旱是影響作物生長的主要因素,與傳統耕作相比,雙免耕覆蓋有利于提高土壤水分含量,有利于作物根系的生長,提高土壤酶活性。這與王 靜等[5]、林雁冰等[6]和邱現奎等[14]的研究結論一致。雙免耕覆蓋有利于冬小麥耕層土壤酶活性的增加,這與秸稈覆蓋后保持土壤水分有較大關系[17],但秸稈類型、還田量及腐解速率對土壤酶活性的影響還需繼續研究。
3.3 雙免耕覆蓋對冬小麥花前花后干物質積累及轉運的影響
雙免耕覆蓋下冬小麥花后干物質積累量和對籽粒的貢獻率均高于傳統耕作,雙免耕覆蓋下冬小麥產量極顯著高于傳統耕作,可能因為雙免耕覆蓋可延緩植株衰老[18],提高冬小麥花后干物質積累,提高花前干物質積累和花后同化物向籽粒轉運的能力,有利于冬小麥產量的提高,這與雷鈞杰[8]和王健波[10]研究結論較為一致。但不同降水年型下雙免耕覆蓋對冬小麥花前、花后干物質積累及轉運的影響還有待做深入研究。
