長鏈非編碼RNA-SLC52A3c在食管癌中的表達特征、功能與臨床意義
2020-01-07 01:26:28張家盛吳健誼廖連娣許秀娥許麗艷李恩民
食管疾病 2019年4期
龍 琳,張家盛,雷 菲,吳健誼,潘 峰,廖連娣,許秀娥,許麗艷,李恩民
食管癌是常見的上消化道惡性腫瘤之一,為全球十大高發(fā)腫瘤之一。主要包括食管鱗癌(esophageal squamous cell cancer,ESCC)與食管腺癌(esophageal adenocarcinoma,EAC)兩種病理類型。在中國,主要以ESCC為主[1-3]。大多數(shù)ESCC患者發(fā)病早期無明顯或特異性臨床癥狀和體征,發(fā)現(xiàn)時多處于中晚期,預后差,5 a生存率僅20%[4-5]。因此,迫切需要從分子水平揭示ESCC發(fā)生的分子機制,為食管癌的診斷和防治提供新的線索與策略。
核黃素轉(zhuǎn)運蛋白家族RFVT/SLC52成員SLC52A3是調(diào)控核黃素吸收及轉(zhuǎn)運的關(guān)鍵蛋白[6]。研究表明,SLC52A3蛋白在ESCC癌組織中呈現(xiàn)異常高表達,敲降SLC52A3會明顯抑制ESCC細胞的增殖能力和平板克隆形成能力;相反,高表達SLC52A3可促進癌細胞增殖,并增強癌細胞在裸鼠上成瘤能力[7]。2018年,本課題組研究發(fā)現(xiàn),SLC52A3蛋白編碼基因編碼兩種不同的蛋白亞型:SLC52A3a和SLC52A3b,它們在食管癌組織細胞中同樣異常高表達,SLC52A3a高表達促進癌細胞增殖,并且在細胞核高表達與食管癌患者術(shù)后生存時間縮短呈顯著正相關(guān)[8-9]。近期,又發(fā)現(xiàn)了一個新的不編碼蛋白的亞型SLC52A3c,然而在ESCC中,此新亞型SLC52A3c對ESCC細胞功能有何影響還不明確,SLC52A3c表達與食管鱗癌患者預后關(guān)系也不清楚。因此,本研究檢測非編碼RNA-SLC52A3c對ESCC細胞增殖和遷移能力的影響,并探討SLC52A3c表達與食管癌患者預后的關(guān)系,期望為探究ESCC發(fā)生的分子機制,以及診斷和防治ESCC提供新的理論依據(jù)。
1 材料與方法
1.1 細胞培養(yǎng)人食管鱗癌細胞系TE3、KYSE180、KYSE150和KYSE510在含10%胎牛血清(Gibco)的RPMI-1640(Invitrogen)培養(yǎng)基中培養(yǎng),人食管鱗癌細胞系SHEEC和人永生化食管鱗狀上皮細胞系SHEE在含10%新生牛血清的DMEM/F12培養(yǎng)基中培養(yǎng),香港永生化食管鱗狀上皮細胞系[10]NE1和NE2在Defined Keratinocyte-SFM/EpiLife Medium with 60 μM Calcium(含0.5%EpiLife Defined Growth Supplement,EDGS;0.1%Defined Keratinocyte-SFM)培養(yǎng)基中培養(yǎng)。細胞培養(yǎng)條件:CO2濃度5%,濕度80%,溫度37 ℃。細胞長成單層后根據(jù)后續(xù)實驗要求,取對數(shù)生長期細胞,胰蛋白酶消化后進行接種或傳代常規(guī)培養(yǎng)。各細胞詳細信息詳見本課題組之前發(fā)表的文章[8,11]。
1.2 3′RACE擴增SLC52A3 cDNA 的3′末端3′RACE實驗采用大連寶生物工程有限公司的3′-Full RACE Core Set Ver.2.0(Code No. D314)試劑盒。選用TE3細胞總RNA,按3′RACE說明書進行3′RACE Adaptor的連接,反轉(zhuǎn)錄獲得雙鏈cDNA,再使用TaKaRa LA Taq?(Code No.DRR02AM)進行套式PCR反應,取5 μL的PCR反應液進行瓊脂糖凝膠電泳,確認3′ RACE PCR擴增產(chǎn)物。PCR擴增產(chǎn)物與pGEM-T vector載體連接,轉(zhuǎn)化到Trans5α感受態(tài)細胞,取陽性克隆菌送往華大基因公司進行測序,測序結(jié)果利用NCBI數(shù)據(jù)庫BLAST進行序列比對分析。3′RACE套式PCR引物見表1。
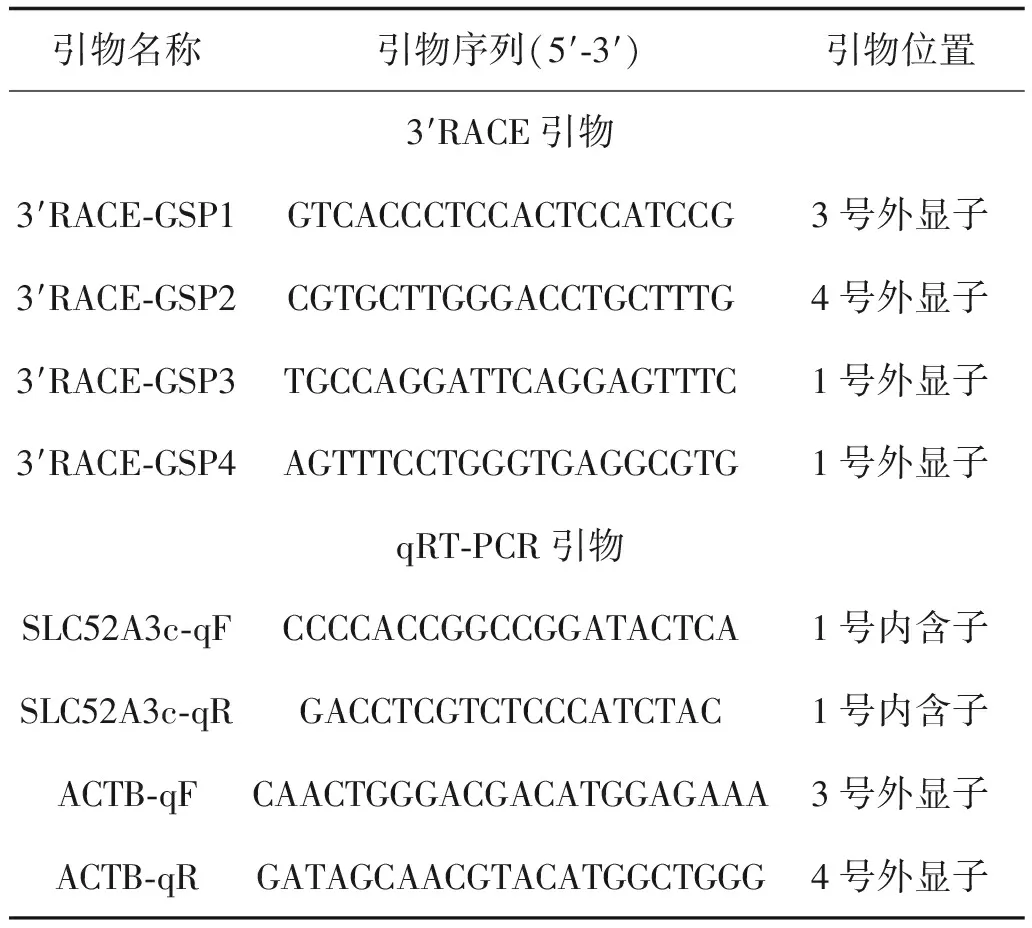
表1 3′RACE擴增SLC52A3本文所使用的引物
1.3 質(zhì)粒構(gòu)建和細胞轉(zhuǎn)染根據(jù)3′RACE擴增的序列,提交SLC52A3c全長699 bp序列由金唯智公司(GENEWIZ)合成,構(gòu)建到表達載體pcDNA3.1上獲得pcDNA3.1-SLC52A3c質(zhì)粒,經(jīng)測序驗證,插入序列準確無誤。ESCC細胞正常傳代后接種到6孔板中,于37 ℃、5% CO2培養(yǎng)箱常規(guī)培養(yǎng)至細胞密度為70%左右。按Lipofectamine3000(Invitrogen)說明書將質(zhì)粒與Lipo3000形成的復合物加入到6孔板中,轉(zhuǎn)染后的細胞繼續(xù)于37 ℃、5% CO2培養(yǎng)箱中培養(yǎng)。6 h后更換10%新鮮培養(yǎng)基繼續(xù)培養(yǎng)48 h。
1.4 RNA提取與實時定量PCR食管鱗癌患者的組織以及轉(zhuǎn)染后的細胞選用Invitrogen的TRIzol 試劑提取總RNA。再采用大連寶生物工程有限公司的反轉(zhuǎn)錄試劑盒和SYBR Green定量PCR試劑盒對提取的總RNA進行反轉(zhuǎn)錄反應和定量PCR檢測。以β-actin引物作內(nèi)參對照。反應結(jié)束后確認定量 PCR 的擴增曲線和融解曲線,以2-△△CT方法分析基因表達相對變化。定量PCR引物見表1。
1.5 細胞增殖能力檢測細胞增殖能力檢測選用實時細胞功能分析(RTCA,xCELLigence Real-Time Cell Analyzer,Roche公司)[12]、平板克隆形成實驗和MTS 3種實驗方法進行。ESCC細胞轉(zhuǎn)染質(zhì)粒36 h后,換無血清培養(yǎng)基培養(yǎng)饑餓細胞12 h,再用胰酶消化、重懸,細胞計數(shù)后,將細胞濃度調(diào)整為1×105個細胞/mL后將細胞分別接種到RTCA E-plate和96孔板中,每孔100 μL(1×104個細胞/孔),另取10 μL細胞懸液到含2 mL培養(yǎng)基的6孔板中(每孔1×103個細胞)。E-plate板中的細胞直接進行細胞增殖能力檢測,每15 min掃描1次;96孔板中細胞選用MTS試劑(Promega)分別檢測各組ESCC細胞在 0、12、36、60和84 h的活性細胞數(shù)量;6孔板中細胞在常規(guī)培養(yǎng)條件下培養(yǎng)2周,當有肉眼可見明顯的細胞克隆時終止培養(yǎng),0.5%結(jié)晶紫染色后計數(shù)每孔的克隆數(shù)。
1.6 細胞遷移能力檢測細胞遷移能力檢測選用劃痕實驗和Transwell移動實驗兩種實驗方法進行。ESCC細胞轉(zhuǎn)染質(zhì)粒36 h后,換用相應的無血清培養(yǎng)基培養(yǎng)饑餓細胞12 h,再用胰酶消化、無血清重懸,細胞計數(shù)后,將細胞濃度調(diào)整為1.25×105個細胞/mL,取400 μL細胞懸液加到Transwell小室上室中,下室加入含血清的正常培養(yǎng)基,CO2培養(yǎng)箱中常規(guī)培養(yǎng) 48 h 后棄去培養(yǎng)基,取出小室,用0.5%結(jié)晶紫染液染色,顯微鏡下觀察細胞并計數(shù)。對于劃痕實驗,將上述細胞接種到6孔板中,每孔2 mL,4 h后換成含2%血清的培養(yǎng)基培養(yǎng),并分別于0、24和48 h在顯微鏡下觀察細胞并拍照,計算細胞遷移率。
1.7 臨床樣本收集本研究實驗選取了汕頭市中心醫(yī)院病理科2007年至2012年183例進行了手術(shù)的癌患者的食管鱗癌組織,183例配對的手術(shù)切緣正常組織。所有病例的臨床資料均來自病歷記錄,根據(jù)隨訪資料,術(shù)后因其他原因死亡或有嚴重術(shù)后并發(fā)癥的病例已剔除。本研究已獲得汕頭大學醫(yī)學院倫理委員會和汕頭市中心醫(yī)院倫理委員會的批準。
1.8 統(tǒng)計分析功能實驗數(shù)據(jù)采用 GraphPad Prism 5軟件計算平均值及標準差,同時繪制成圖。應用SSPS 16.0軟件對各組實驗數(shù)據(jù)進行 Two-way ANOVA 檢驗,確定它們之間差異是否有統(tǒng)計學意義。通過威爾科克森符號秩檢驗(Wilcoxon)分析SLC52A3c在食管鱗癌患者的食管鱗癌組織和手術(shù)切緣正常組織中的表達量是否有差異。然后通過Kaplan-Meier單變量生存期分析分別評估SLC52A3c的表達量與食管鱗癌患者生存和無瘤生存情況的關(guān)系。采用多因素Cox風險回歸模型逐步向前法評估各臨床病理參數(shù)及SLC52A3c是否可作為食管鱗癌患者預后的獨立影響因素。以P<0.05為差異有統(tǒng)計學意義。
2 結(jié)果
2.1 3′RACE擴增SLC52A3-3′UTR為了確定SLC52A3基因不同亞型的3′末端,作者在SLC52A3基因第3、第4外顯子以及第1外顯子上分別設(shè)計了3′RACE引物(見表1)擴增SLC52A3(見圖1A),瓊脂糖電泳各得到了多個條帶,進行DNA測序鑒定后發(fā)現(xiàn)其中有亞型SLC52A3a的3′末端和新亞型SLC52A3c(圖1B)。從圖1C可以看出:① SLC52A3c-lncRNA與來自同一基因結(jié)構(gòu)框架的蛋白編碼轉(zhuǎn)錄本 SLC52A3a和SLC52A3b是一種同向順式序列關(guān)系;② SLC52A3c-lncRNA的轉(zhuǎn)錄起始位點雖與SLC52A3a相同,但其1號外顯子序列比Ensemble數(shù)據(jù)庫提供的序列多出115 bp,其2號外顯子序列與Ensemble數(shù)據(jù)庫提供的序列相比,多出13 bp,因此其總長為699 bp。SLC52A3c轉(zhuǎn)錄本的RNA信息已提交到了GenBank數(shù)據(jù)庫(GenBank:MN534336)。
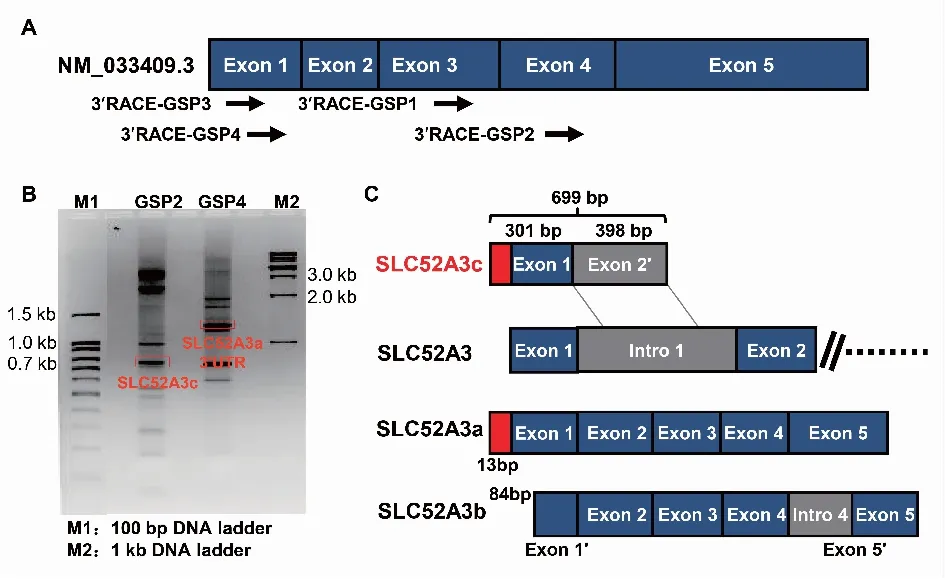
A:3′RACE擴增SLC52A3-3′UTR引物位置示意圖;B:3′RACE PCR擴增產(chǎn)物瓊脂糖電泳圖;C:測序結(jié)果模擬圖,SLC52A3c全長699 bp。圖1 3′RACE鑒定lncRNA SLC52A3c
2.2 pcDNA3.1-SLC52A3c質(zhì)粒表達鑒定為了獲得高表達SLC52A3c的食管鱗癌細胞模型,本研究選用食管鱗癌細胞系TE3、SHEEC、KYSE510和KYSE150分別轉(zhuǎn)染pcDNA3.1-SLC52A3c質(zhì)粒,然后提取各細胞總RNA,通過定量PCR方法鑒定SLC52A3c的高表達,如圖2所示,在各個食管鱗癌細胞系中,SLC52A3c均高表達成功。
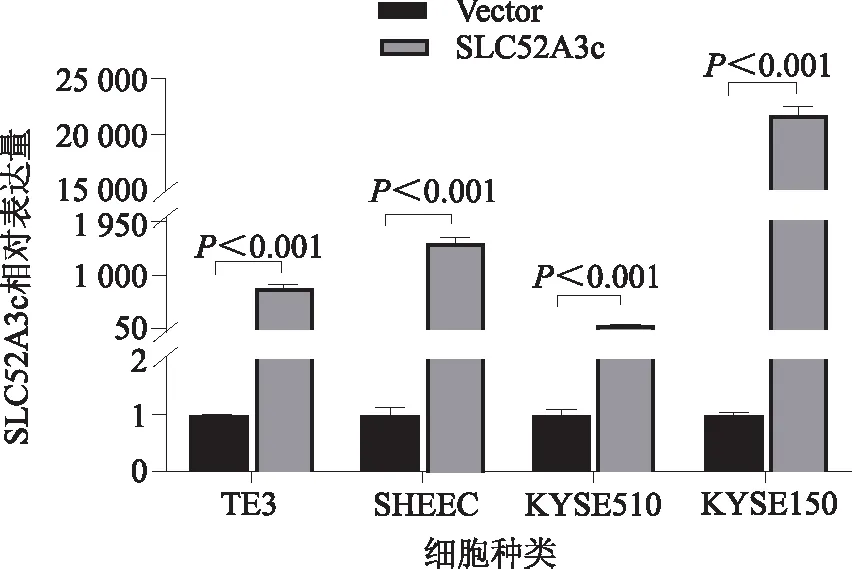
圖2 SLC52A3c在食管鱗癌細胞系中的高表達效果
2.3 高表達SLC52A3c抑制食管鱗癌細胞的分裂增殖能力實時細胞功能分析實驗、平板克隆形成實驗和MTS 3種實驗來確認SLC52A3c高表達對食管鱗癌細胞增殖能力的影響。RTCA實驗結(jié)果如圖3A,在TE3、SHEEC和KYSE510細胞中,相對于轉(zhuǎn)染pcDNA3.1空載體的對照組,高表達SLC52A3c后細胞生長曲線較低,細胞的生長速率較慢,細胞的增殖能力明顯受到抑制。細胞平板克隆形成實驗結(jié)果和統(tǒng)計結(jié)果如圖3B、3C,在TE3、SHEEC和KYSE510細胞中,無論是克隆的大小還是克隆的數(shù)量相對于對照組,高表達SLC52A3c后3種食管鱗細胞生長速度均較慢,克隆較小,克隆數(shù)目較少,細胞的增殖能力明顯受到抑制。在TE3和SHEEC細胞中,用MTS實驗進行了重復實驗,結(jié)果如圖3D,高表達SLC52A3c后細胞的生長曲線同樣較低,生長速度較慢,食管鱗癌細胞的增殖速度明顯減弱。這些結(jié)果表明,高表達SLC52A3c可以抑制食管鱗癌細胞的分裂增殖能力。
2.4 高表達SLC52A3c抑制食管鱗癌細胞的遷移能力細胞劃痕實驗結(jié)果如圖4A,在食管鱗癌細胞SHEEC和KYSE510中,與對照組相比,轉(zhuǎn)染SLC52A3c后劃痕“愈合”速度變慢,SHEEC細胞移動能力顯著減弱,相同時間內(nèi)愈合范圍更小。因為SHEEC細胞在劃痕實驗中劃痕“愈合”速度減弱程度更顯著,在本實驗中再選用SHEEC細胞使用Transwell移動實驗確證SLC52A3c高表達抑制食管鱗癌細胞遷移能力的實驗結(jié)果。實驗結(jié)果和統(tǒng)計結(jié)果如圖4B,與對照組相比,SLC52A3c高表達組誘導穿過小室的細胞數(shù)目明顯減少。上述實驗結(jié)果提示,在食管鱗癌細胞中SLC52A3c高表達顯著抑制癌細胞的遷移能力。
2.5 食管鱗癌細胞系和食管鱗癌組織中SLC52A3c的表達特征細胞功能實驗表明,食管鱗癌細胞中,SLC52A3c高表達發(fā)揮抑制癌細胞分裂增殖和遷移的作用,那么,在食管鱗癌細胞系和相對正常的永生化食管鱗狀上皮細胞系以及食管鱗癌組織與手術(shù)切緣正常食管上皮組織中,SLC52A3c表達水平如何?為了解這一情況,作者采用熒光定量PCR方法檢測了5種食管鱗癌細胞和3種永生化食管上皮細胞以及183對食管鱗癌組織與手術(shù)切緣正常食管上皮組織中SLC52A3c的表達水平,結(jié)果如圖5所示,SLC52A3c在食管鱗癌細胞中的表達水平顯著低于永生化鱗狀食管上皮細胞,食管鱗癌組織中SLC52A3c表達水平也明顯低于手術(shù)切緣正常食管上皮組織,根據(jù)這些結(jié)果結(jié)合功能實驗,作者推測SLC52A3c可能發(fā)揮抑癌作用。
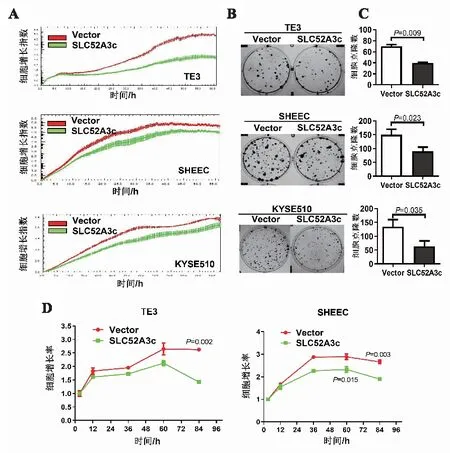
A:食管鱗癌細胞中高表達SLC52A3c,RTCA檢測細胞增殖能力變化;B:食管鱗癌細胞中高表達SLC52A3c,平板克隆形成實驗檢測細胞增殖能力變化;C:細胞增殖能力對比;D:食管鱗癌細胞中高表達SLC52A3c,MTS檢測細胞活力。圖3 高表達SLC52A3c抑制食管鱗癌細胞的分裂增殖能力
2.6 SLC52A3c高表達與食管鱗癌患者無瘤生存時間正相關(guān)接下來,作者進一步將定量PCR結(jié)果與有完整生存狀態(tài)的183例食管鱗癌患者資料結(jié)合起來,分析SLC52A3c與患者預后的關(guān)系。選用X-tile軟件,根據(jù)SLC52A3c和病人無瘤生存狀態(tài)找到Cutoff值,即卡方最大點,將SLC52A3c分成兩個組(即High expression組及Low expression組),分析分組后SLC52A3c與食管癌患者生存時間或無瘤生存時間之間的關(guān)系。Kaplan-Meier曲線法分析結(jié)果發(fā)現(xiàn),SLC52A3c表達水平與總生存時間未見顯著相關(guān)性(P>0.05),但SLC52A3c表達水平對病人無瘤生存時間發(fā)現(xiàn)有明顯影響,相對于SLC52A3c低表達的病人,SLC52A3c高表達的病人無瘤生存時間更長,且具有顯著相關(guān)性(P=0.017)。這些實驗結(jié)果說明SLC52A3c是一個保護性因子,在食管鱗癌中起抑制腫瘤的作用,SLC52A3c高表達可預警食管鱗癌患者有較好的預后。
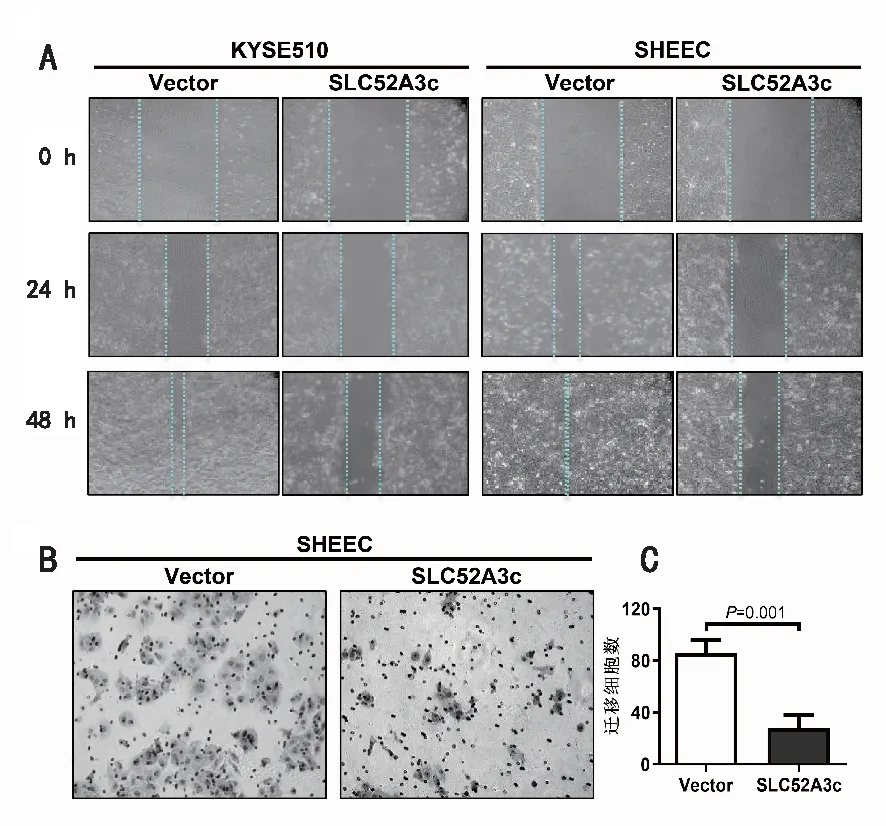
A:劃痕實驗檢測高表達SLC52A3c后食管鱗癌細胞遷移能力的變化; B:Transwell移動實驗檢測高表達SLC52A3c后食管鱗癌細胞移動能力的變化; C:對圖B的定量結(jié)果。圖4 高表達SLC52A3c抑制食管鱗癌細胞的遷移能力
2.7 SLC52A3c高表達是食管鱗癌患者無瘤生存的獨立保護因素通過對年齡、性別、飲酒、TNM分期、吸煙、腫物大小與食管鱗癌病人癌組織中SLC52A3c表達量進行單因素及多因素Cox回歸分析,結(jié)果顯示TNM分期(P=0.000)和SLC52A3c(P=0.017)可以作為食管鱗癌術(shù)后患者復發(fā)轉(zhuǎn)移的獨立影響因素,SLC52A3c可作為食管鱗癌組織中獨立預后因子,SLC52A3c表達水平越高,病人的預后越好。詳見表2。
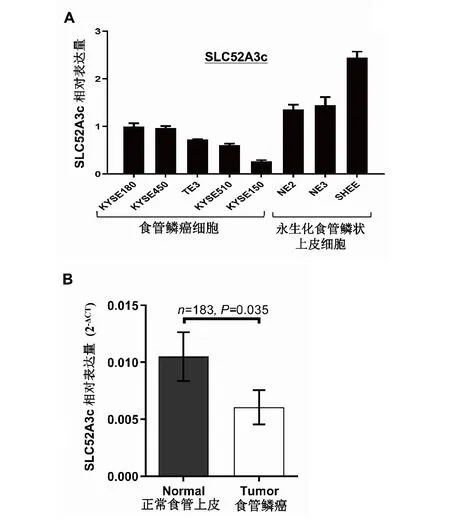
A:定量PCR檢測5種食管鱗癌細胞和3種永生化食管鱗狀上皮細胞中SLC52A3c的表達量;B:定量PCR檢測183對食管鱗癌組織和配對的手術(shù)切緣正常食管上皮組織中SLC52A3c的表達量。圖5 食管鱗癌細胞系和食管鱗癌組織中SLC52A3c的表達特征
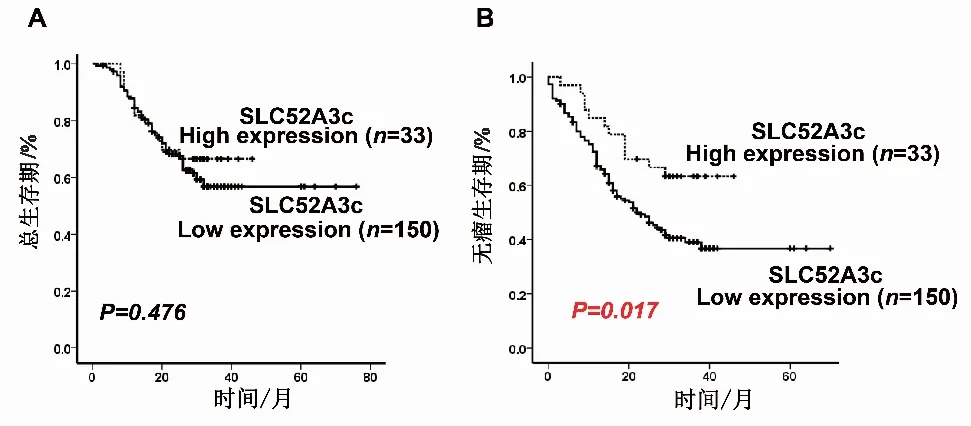
A:SLC52A3c表達水平與食管鱗癌患者總生存時間的Kaplan-Meier分析;B:SLC52A3c表達水平與食管鱗癌患者無瘤生存時間的Kaplan-Meier分析。圖6 SLC52A3c表達水平與食管鱗癌患者總生存時間和無瘤生存時間的Kaplan-Meier分析
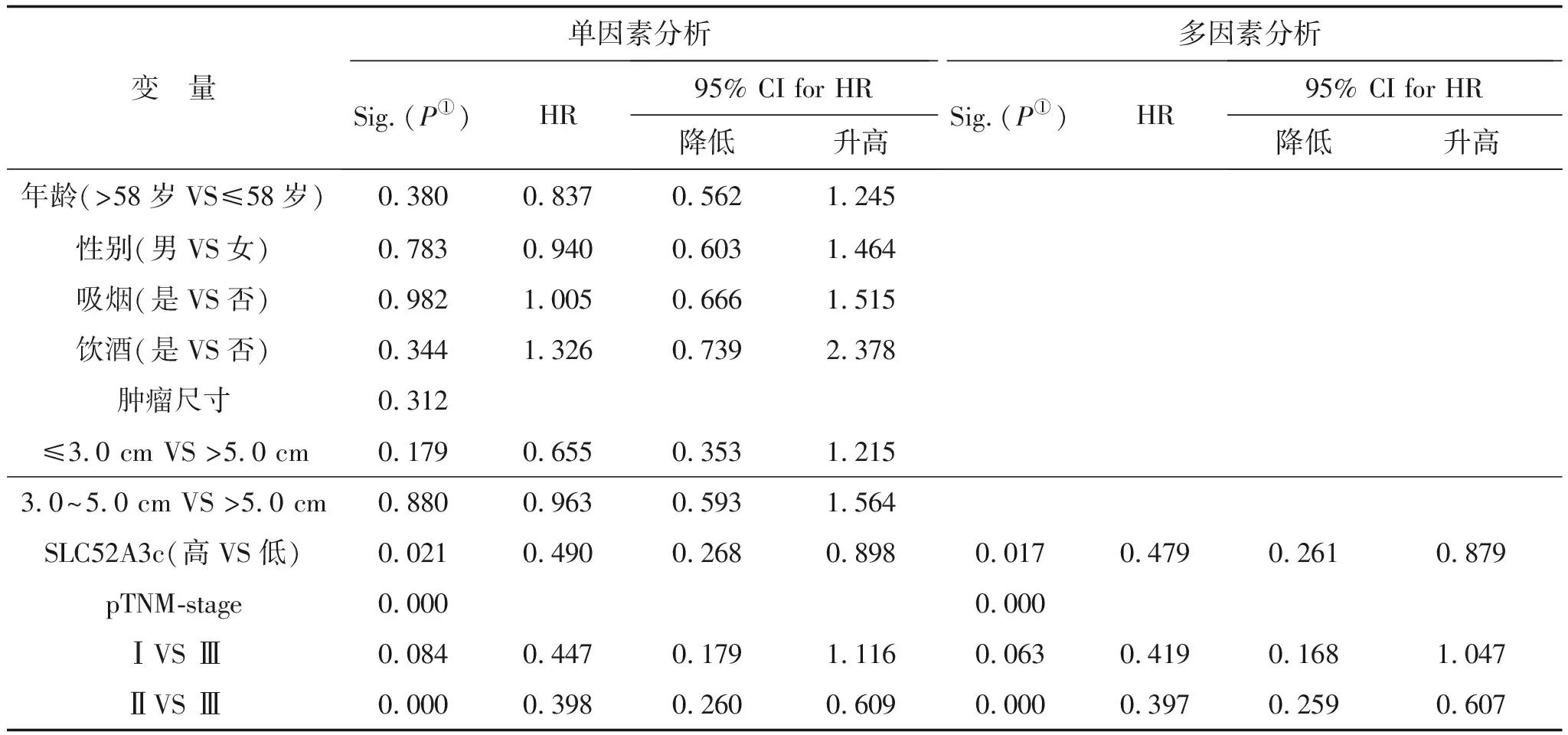
表2 SLC52A3c與食管鱗癌患者無瘤生存預后的單因素和多因素COX回歸分析
注:①P<0.05為差異有統(tǒng)計學意義;HR:hazards ratio,風險率;CI:confidence interval,置信區(qū)間;多因素分析采用COX比例風險模型;單因素分析采用變量作為預測指標。
3 討論
核黃素是黃素腺嘌呤單核苷酸(flavin mononucleotide,FMN)和黃素腺嘌呤二核苷酸(flavin adenine dinucleotide,FAD)的前體,在細胞能量代謝中起著至關(guān)重要的作用,其缺乏可能導致DNA損傷、細胞應激反應和腫瘤的發(fā)生[11,13-14]。SLC52A3,SLC52家族的一員,是一個非常重要的核黃素轉(zhuǎn)運者,在維持體內(nèi)核黃素動態(tài)平衡中發(fā)揮著至關(guān)重要的作用[15-16]。研究表明,SLC52A3在ESCC的發(fā)生和發(fā)展中同樣發(fā)揮重要作用。在食管鱗癌細胞中,SLC52A3表達敲降,抑制細胞增殖;SLC52A3a高表達促進細胞增殖,SLC52A3a在細胞核高表達與食管鱗癌患者術(shù)后生存時間呈現(xiàn)顯著負相關(guān)關(guān)系,SLC52A3b在細胞質(zhì)中高表達與食管鱗癌患者術(shù)后生存時間呈顯著正相關(guān)關(guān)系,檢測SLC52A3 在食管鱗癌組織細胞中表達定位可以預測食管癌患者預后[7-8]。
在本研究中發(fā)現(xiàn)了SLC52A3的一個新的非編碼蛋白的lncRNA亞型,SLC52A3c,且研究發(fā)現(xiàn)SLC52A3c對于食管鱗癌細胞的功能作用與其他亞型作用相反,SLC52A3c高表達抑制食管鱗癌細胞的增殖和遷移。那么,SLC52A3c與SLC52A3a和SLC52A3b的表達調(diào)控以及與食管癌發(fā)生發(fā)展之間究竟有何種聯(lián)系?分析SLC52A3基因3個不同轉(zhuǎn)錄本SLC52A3a/SLC52A3b/SLC52A3c,可以知道它們來自于同一基因結(jié)構(gòu)框架,在SLC52A3 5′側(cè)翼區(qū)轉(zhuǎn)錄調(diào)控區(qū)和3′端非翻譯區(qū)上,序列相互重疊,因此SLC52A3a/SLC52A3b/SLC52A3c三者間在轉(zhuǎn)錄層面或轉(zhuǎn)錄后可能存在相互作用,相互影響。有研究表明,PTEN基因的其中的一個轉(zhuǎn)錄本lncRNA PTENpg1,可作為miRNA海綿捕獲靶向PTEN的miRNA,在轉(zhuǎn)錄后水平調(diào)控PTEN的表達[17]。基于此,可以猜測,SLC52A3c也許同樣會通過捕獲miRNA來調(diào)控SLC52A3的表達,SLC52A3c作為lncRNA,很有可能與SLC52A3一起,形成自反饋調(diào)節(jié),影響核黃素的轉(zhuǎn)運和能量代謝途徑,改變食管鱗癌的發(fā)生發(fā)展進程。在今后的研究中,作者擬進一步探究三者表達過程中的相關(guān)性,深入探究SLC52A3在食管鱗癌中發(fā)揮作用的分子機制。
臨床上,治療食管鱗癌的方案,主要有手術(shù)、放療和化療等。一般來說,對于那些晚期,可以直接手術(shù)的食管鱗癌患者,往往是術(shù)后輔助放療或/和化療。目前,指導臨床食管鱗癌治療方案選擇的標準是TNM分期。與其它惡性腫瘤一樣,食管鱗癌有著很強的異質(zhì)性,這使得僅僅依靠TNM分期,往往并不能達到精準治療的目的,許多時候會出現(xiàn)治療不足,或過度治療的情況。所以,人們寄希望于尋找分子標志物,以完善食管鱗癌患者的治療方案。以往報道的食管鱗癌以及其他癌癥的預后預警分子標志物,主要集中在蛋白編碼基因。近年來,miRNA和lncRNA已成為新的腫瘤預后預警分子標志物的研究熱點。
HOTAIR是研究最廣泛的具有致癌作用的lncRNA之一,在原始乳腺癌的轉(zhuǎn)移中被發(fā)現(xiàn)。越來越多的研究表明,HOTAIR高表達使食管癌患者變差,促進淋巴結(jié)轉(zhuǎn)移,總生存率變低,促進癌細胞的轉(zhuǎn)移[18-20]。Meta分析發(fā)現(xiàn),高表達的HOTAIR是強有力的預后因子,可作為食管癌患者治療的靶標[21]。在本研究中發(fā)現(xiàn)的新lncRNA-SLC52A3c,在食管鱗癌細胞系中表達水平明顯低于永生化鱗狀食管上皮細胞,在食管鱗癌組織表達量明顯低于手術(shù)切緣正常食管上皮組織,并且SLC52A3c高表達與食管鱗癌患者無瘤生存時間正相關(guān),是食管鱗癌患者無瘤生存的獨立的保護性因素,這些結(jié)果表明SLC52A3c的確是一個抑癌性lncRNA,在食管鱗癌中發(fā)揮保護性作用,并抑制食管鱗癌的轉(zhuǎn)移。
綜上所述,SLC52A3c作為一個保護性因子,通過抑制食管鱗癌細胞增殖和遷移能力,抑制食管鱗癌的發(fā)生發(fā)展。在食管鱗癌的lncRNA研究中,許多l(xiāng)ncRNA,如HOTAIR、MALAT1、NEAT1、POU3F3和CCAT2均可發(fā)揮致癌作用[18,22-26],因此SLC52A3c作為抑制性因子為食管鱗癌中l(wèi)ncRNA研究打開新的局面。此外,SLC52A3c還可成為食管鱗癌的早期診斷、預后預警及分子分型的有效的分子標志物。同時SLC52A3c還可能作為食管鱗癌靶向治療的分子藥靶,開發(fā)以SLC52A3c為靶點的食管鱗癌的靶向藥物。總之,本研究探究SLC52A3c在ESCC發(fā)生的分子機制,為ESCC的早診和防治提供了理論依據(jù)。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
發(fā)明與創(chuàng)新(2022年30期)2022-10-03 08:40:56
中學生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
人大建設(shè)(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
