羅非魚性別相關(guān)微衛(wèi)星標(biāo)記的初步篩選
2020-01-15 07:45:50陳文偉曹建萌劉志剛盧邁新
淡水漁業(yè) 2020年1期
陳文偉,曹建萌,劉志剛,楊 娜,盧邁新
(1.中國水產(chǎn)科學(xué)研究院珠江水產(chǎn)研究所,農(nóng)業(yè)農(nóng)村部熱帶亞熱帶水產(chǎn)資源利用與養(yǎng)殖重點實驗室,廣州 510380;2.上海海洋大學(xué)水產(chǎn)與生命學(xué)院,上海 201306)
羅非魚性成熟時間早,性成熟后產(chǎn)卵周期短,如果雌雄混養(yǎng)會因群體內(nèi)自繁進而導(dǎo)致人工養(yǎng)殖的羅非魚種群密度過大,規(guī)格參差不齊,極大降低了羅非魚的經(jīng)濟效益,并且羅非魚雄魚比雌魚更具生長優(yōu)勢,因此目前羅非魚的商業(yè)化生產(chǎn)往往依賴雄性的單性養(yǎng)殖。現(xiàn)已有許多方法可控制羅非魚雄性化[1-6],但在實際生產(chǎn)過程中都有一定的局限性[7]。
微衛(wèi)星標(biāo)記具有多態(tài)性高、雜合度大、容易擴增與檢測等優(yōu)勢,被廣泛應(yīng)用于遺傳圖譜的構(gòu)建、親子鑒定、遺傳多樣性分析、動物經(jīng)濟性狀的連鎖分析等,羅非魚性別相關(guān)微衛(wèi)星標(biāo)記的開發(fā)已有大量研究。Lee等[8]利用UNH995與UNH104(189 bp等位基因與Y染色體連鎖)正確地推測了兩個尼羅羅非魚(Oreochromisniloticus)家系中95%的雄性和雌性個體的性別,利用GM354(129 bp雄性獨有)、UNH168、GM271(121 bp雄性獨有)和UNH131(193 bp等位基因與Z染色體連鎖,187 bp的等位基因與W染色體連鎖)正確地推測了一個奧利亞羅非魚(O.aureus)家系中97%雄性和85%雌性個體的性別[9]。Khan等[10]發(fā)現(xiàn)UNH995與一個尼羅羅非魚家系(HRT26)的性別顯著相關(guān)。張庭等[11]在篩選奧利亞羅非魚微衛(wèi)星位點時發(fā)現(xiàn)UNH168在奧利亞羅非魚家系的雌性個體中可擴增出135 bp和171 bp兩個條帶,而在該家系雄性個體中只有171 bp一個條帶。根據(jù)目前的研究,尼羅羅非魚的性別決定區(qū)域定位于連鎖群(Linkage group,LG)1和LG23上[8,12-15],而奧利亞羅非魚的性別決定區(qū)域定位于LG1和LG3上[9,16]。本研究擬通過對不同羅非魚群體遺傳多樣性的分析,以及對兩個尼羅羅非魚群體和兩個奧利亞羅非魚群體性別相關(guān)微衛(wèi)星標(biāo)記的篩選和分析,以期為今后羅非魚育種以及羅非魚遺傳性別的鑒定提供基礎(chǔ)資料。
1 材料和方法
1.1 羅非魚來源及基因組DNA提取
實驗魚詳細(xì)信息見表1,本實驗所用實驗魚均為混合群體。剪取所有實驗魚尾鰭,浸泡在無水乙醇中于-20 ℃保存?zhèn)溆谩J褂肈Neasy Blood & Tissue Kit試劑盒(購自北京柏奧易思生物科技公司)提取基因組DNA,1.5%瓊脂糖凝膠電泳檢測DNA的質(zhì)量,微量分光光度計(OSE-260,購自北京天根生化科技公司)檢測DNA的濃度及純度;用ddH2O將基因組DNA稀釋至終濃度為50 ng/μL,置于-20 ℃保存?zhèn)溆谩?/p>
1.2 PCR擴增及產(chǎn)物檢測
1.2.1 兩個尼羅羅非魚群體及兩個奧利亞羅非魚群體性別相關(guān)微衛(wèi)星標(biāo)記的初步篩選
參考羅非魚微衛(wèi)星遺傳圖譜,從LG1、LG3及LG23上選擇24個微衛(wèi)星位點,從GenBank上下載這些微衛(wèi)星位點引物序列 ,并由金唯智生物科技公司合成,引物詳細(xì)信息見表2。對每對引物的擴增條件進行優(yōu)化后,在兩個尼羅羅非魚群體及兩個奧利亞羅非魚群體中進行初步擴增,PCR反應(yīng)體系為20 μL,包括:50 ng/μL DNA模板1 μL,10 μmol/L上、下游引物各0.4 μL,2×EasyTaq PCR SuperMix for PAGE 10 μL,ddH2O 8.2 μL。PCR熱循環(huán)程序為:94 ℃預(yù)變性3 min,94 ℃變性30 s,X ℃退火30 s,72 ℃延伸30 s,循環(huán)數(shù)為25~28個,最后72 ℃終延伸5 min,4 ℃保存。PCR產(chǎn)物經(jīng)12%非變性聚丙烯酰胺凝膠電泳分離,產(chǎn)物上樣量為5 μL,50 bp ladder上樣量為3 μL,電泳緩沖液為1×TBE。先使用50 V低電壓電泳30 min,再使用150 V電壓電泳150 min。銀染參考許紹斌等[17]的方法并適當(dāng)調(diào)整,同一位點等位基因由大到小記為A、B、C…。統(tǒng)計各微衛(wèi)星位點的等位基因種類和數(shù)目,并在所選樣本內(nèi)部雌雄群體之間進行卡方檢驗,初步篩選跟性別相關(guān)的微衛(wèi)星標(biāo)記。
1.2.2 初步篩選的微衛(wèi)星標(biāo)記在各群體中的擴增驗證
在兩個尼羅羅非魚群體與兩個奧利亞羅非魚群體中初步篩選到與性別相關(guān)的微衛(wèi)星標(biāo)記后,在這些微衛(wèi)星標(biāo)記上游引物的5′端添加不同顏色的熒光標(biāo)記序列(FAM,HEX),統(tǒng)一在全部群體中進行擴增,擴增采用降落PCR,PCR反應(yīng)體系為10 μL,包括:50 ng/μL DNA模板1 μL,10 μmol/L上、下游引物各0.5 μL,2×Taq PCR Master Mix 5 μL,ddH2O 3 μL。PCR擴增程序為:95 ℃預(yù)變性5 min,95 ℃變性30 s,62~52 ℃退火30 s,72 ℃延伸30 s,共進行10個循環(huán),每個循環(huán)退火溫度降低1 ℃;之后95 ℃變性30 s,52 ℃退火30 s,72 ℃延伸30 s,共進行22~25個循環(huán),最后72 ℃末端延伸20 min后,4 ℃保存。帶熒光的PCR產(chǎn)物經(jīng)DNA測序儀ABI 3730xl進行熒光電泳檢測,并使用軟件GeneMarker V2.2.0 對原始數(shù)據(jù)進行條帶分型。統(tǒng)計所有群體的各項遺傳多樣性參數(shù),并進行聚類分析,統(tǒng)計各微衛(wèi)星位點的等位基因種類和數(shù)目,并在兩個尼羅羅非魚及兩個奧利亞羅非魚群體內(nèi)部雌雄之間進行卡方檢驗,進一步篩選跟性別相關(guān)的微衛(wèi)星標(biāo)記。
1.3 數(shù)據(jù)分析
用軟件Popgone32和GenAlEx 6.50計算各微衛(wèi)星位點的觀測等位基因數(shù)(Ne)、有效等位基因數(shù)(Na)、觀測雜合度(Ho)、期望雜合度(He)、香農(nóng)信息指數(shù)(I)、Nei’s遺傳距離和遺傳一致度,并檢驗各位點各群體是否偏離哈代溫伯格定律,用軟件PIC-CALC以及Cervus計算各位點的多態(tài)信息含量(PIC),用POPTREE軟件對各群體進行NJ聚類分析。

表2 24對微衛(wèi)星引物信息Tab.2 Information of 24 pairs of microsatellite primers
2 結(jié)果與分析
2.1 兩個尼羅羅非魚群體及兩個奧利亞羅非魚群體性別相關(guān)微衛(wèi)星標(biāo)記的初步篩選結(jié)果與分析
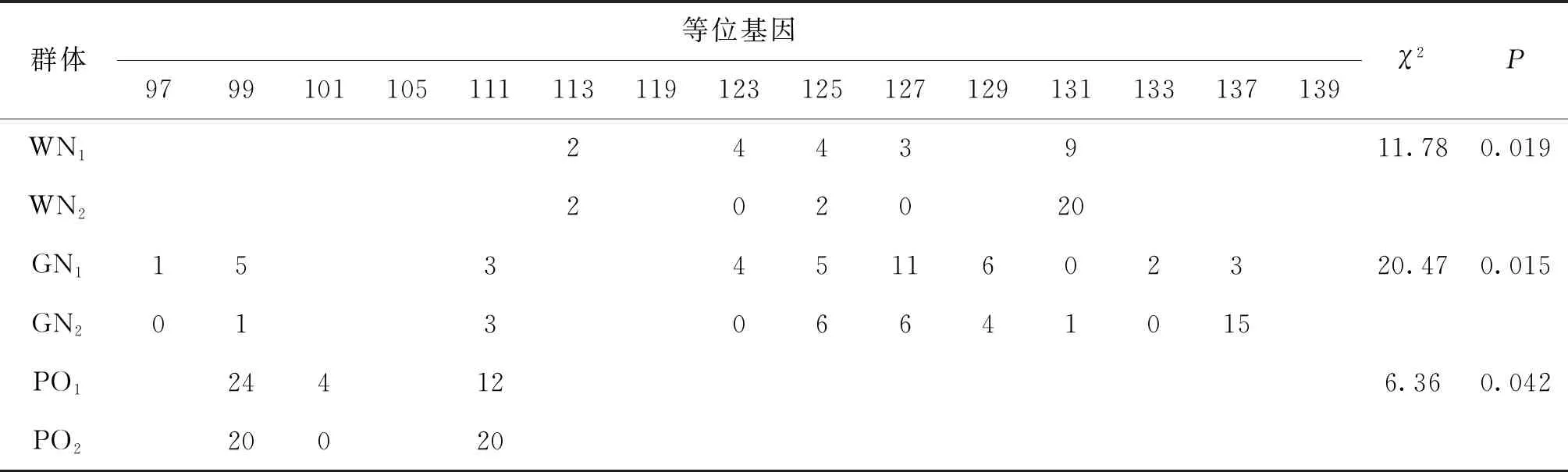
將24個微衛(wèi)星標(biāo)記在WN1、WN2、GN1、GN2、WO1、WO2、PO1以及PO2群體中擴增,利用Popgene32軟件統(tǒng)計各微衛(wèi)星位點的等位基因種類和數(shù)目,并在所有羅非魚樣本群體內(nèi)部雌雄群體之間進行卡方檢驗,最終篩選到UNH931、GM128、GM201、GM258、GM597以及UNH898共6個與性別相關(guān)的微衛(wèi)星標(biāo)記。在無錫尼羅羅非魚群體(WN1,WN2)中,UNH898與性別顯著相關(guān)(P<0.05),GM597與性別極顯著相關(guān)(P<0.01)。在高要尼羅羅非魚群體(GN1,GN2)中,UNH931與性別顯著相關(guān),GM597、UNH898、GM128與性別極顯著相關(guān)。在無錫奧利亞羅非魚群體(WO1,WO2)中,GM201與性別顯著相關(guān)。在番禺奧利亞羅非魚群體(PO1,PO2)中,GM258、GM597與性別顯著相關(guān),詳細(xì)結(jié)果見表3。
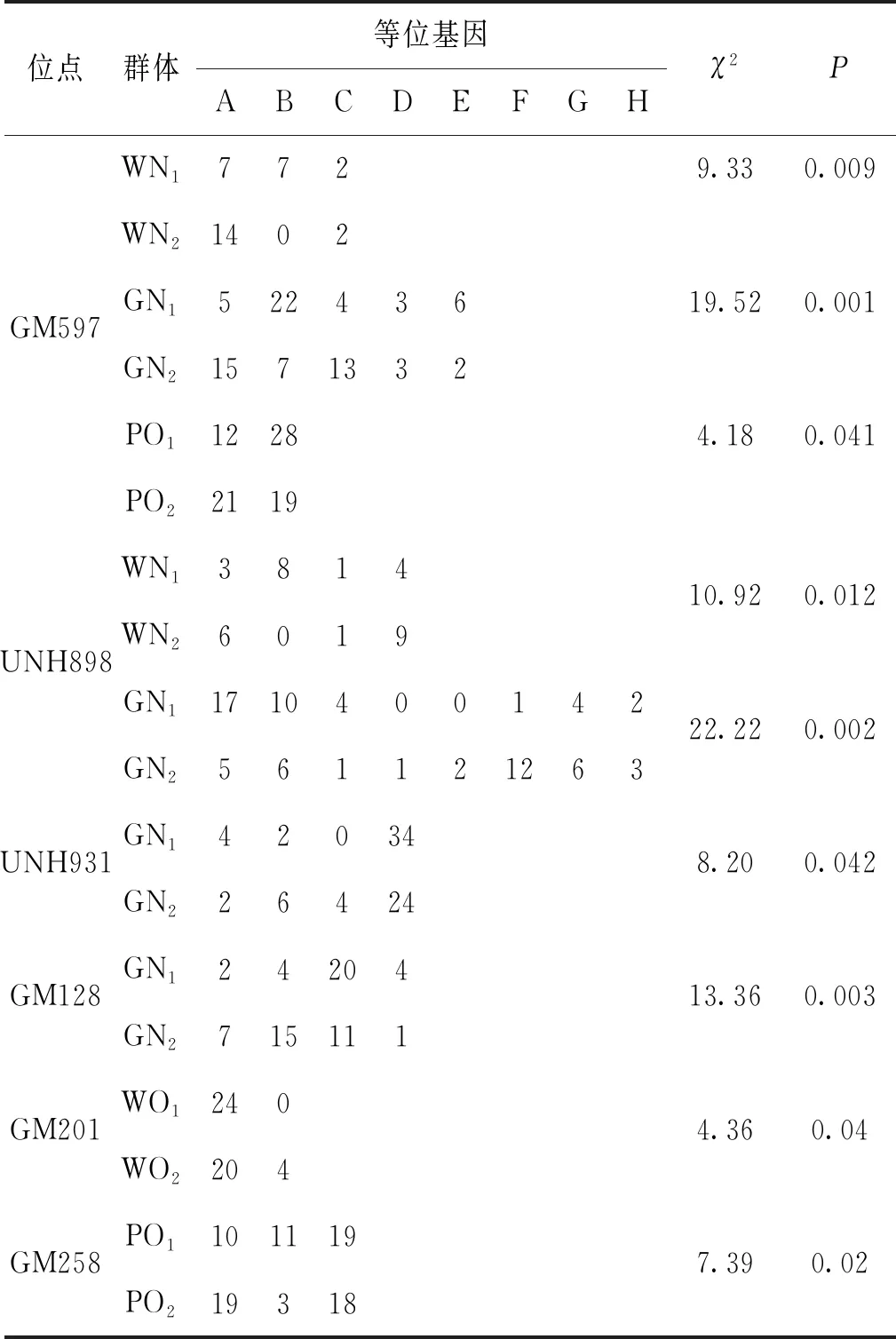
表3 兩個尼羅羅非魚群體及兩個奧利亞羅非魚群體性別相關(guān)微衛(wèi)星標(biāo)記統(tǒng)計Tab.3 Statistics of sex-related microsatellite markers of two O.niloticus populations and two O.aureus populations
2.2 初步篩選到的性別相關(guān)微衛(wèi)星標(biāo)記在不同養(yǎng)殖群體中的擴增結(jié)果與分析
2.2.1 微衛(wèi)星擴增結(jié)果
將初步篩選到的6個跟性別相關(guān)的微衛(wèi)星標(biāo)記在全部群體中擴增,均可得到相應(yīng)的產(chǎn)物。6個微衛(wèi)星標(biāo)記在所有群體297個樣本中共檢測到95個等位基因,其大小在97~302 bp之間,各位點在各群體等位基因數(shù)在1~13個之間。
2.2.2 群體遺傳多樣性參數(shù)統(tǒng)計
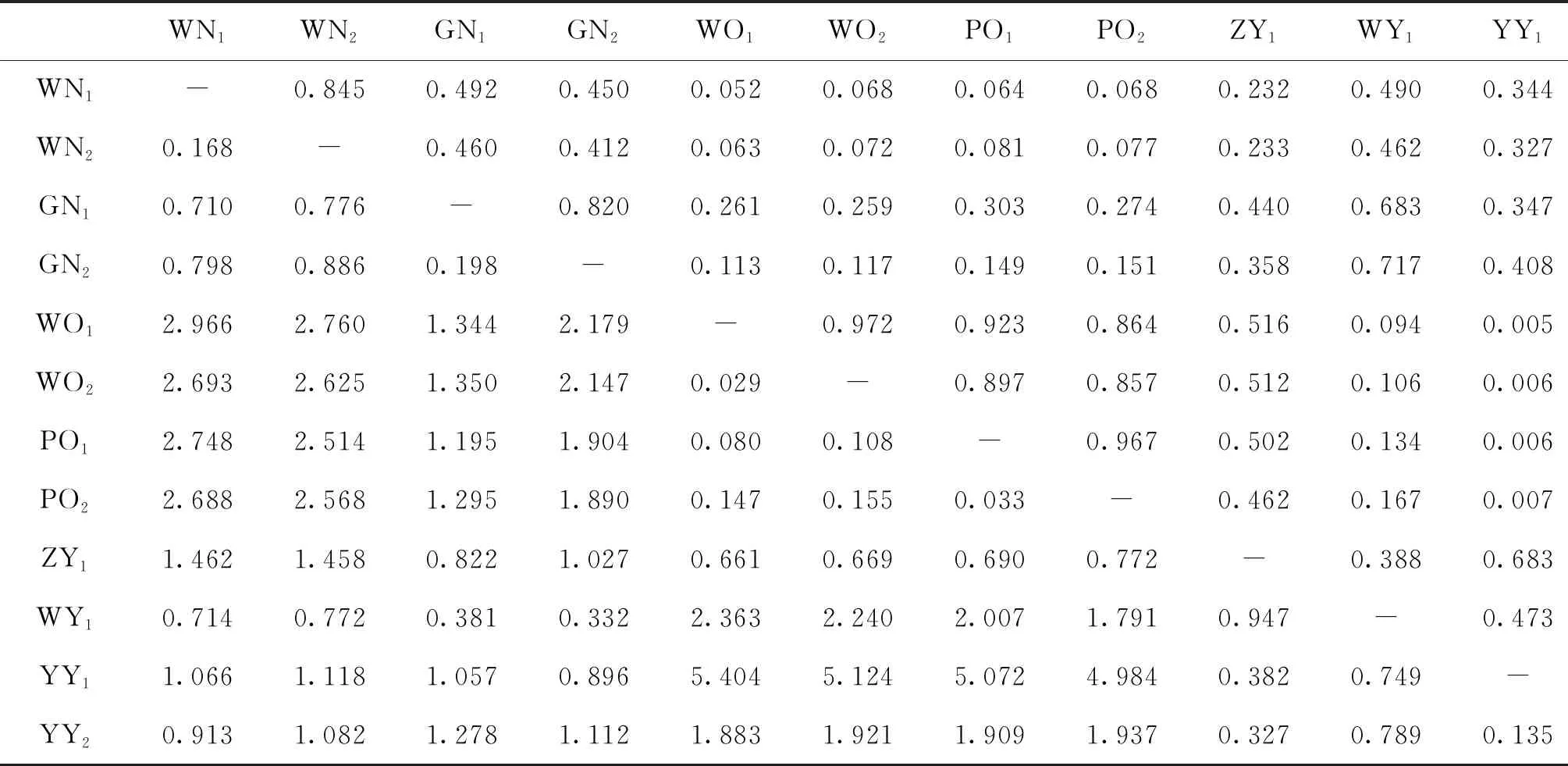
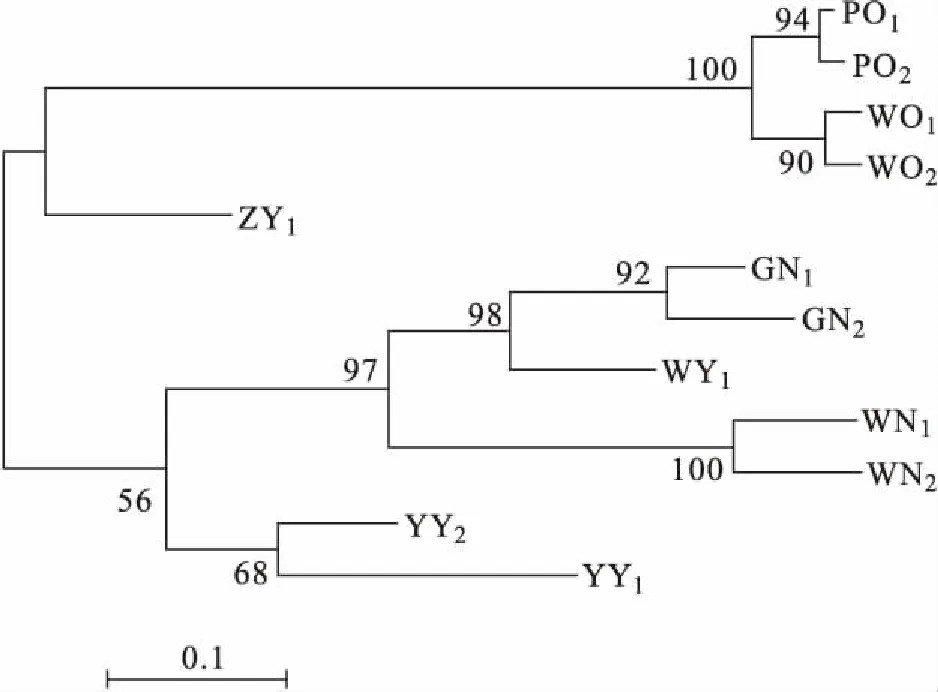
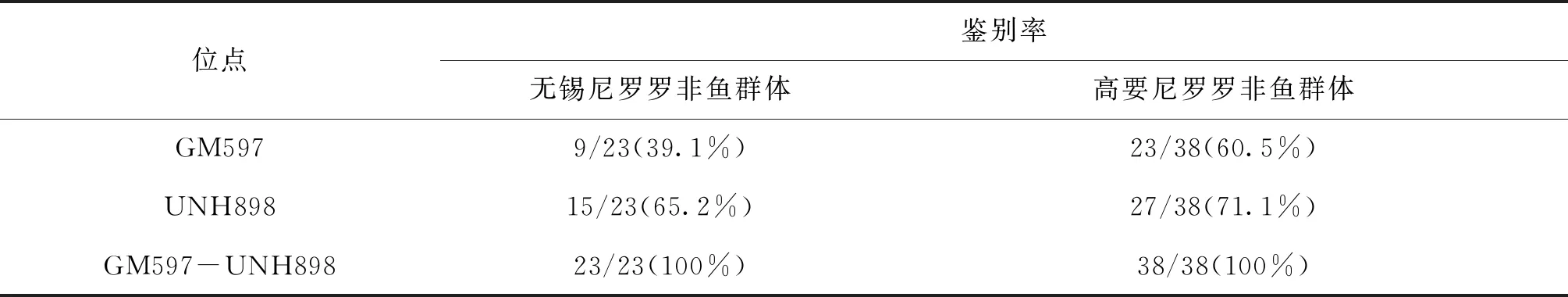
各群體平均觀測等位基因數(shù)為2.167~9.333,平均有效等位基因數(shù)為1.624~4.966,平均觀測雜合度為0.324~0.983,平均期望雜合度為0.329~0.782,平均多態(tài)信息含量為0.275~0.753。YY1群體多態(tài)性最低,WY群體多態(tài)性最高,WN1、WN2、GN1、GN2、WY1以及YY2群體達到高度多態(tài)(PIC>0.5),WO1、WO2、PO1、PO2、ZY1以及YY1群體為中度多態(tài)(0.25 表4 所有羅非魚群體的平均遺傳多樣性參數(shù)Tab.4 Average genetic diversity parameters for all tilapia populations 2.2.3 遺傳距離、遺傳一致度以及聚類分析 YY1群體與WO1群體間遺傳距離最大(5.404),遺傳一致度最小(0.005),因此兩者親緣關(guān)系最遠(yuǎn)。WO2群體與WO1群體之間遺傳距離最小(0.029),遺傳一致度最大(0.972),因此兩者親緣關(guān)系最近。無錫尼羅羅非魚群體(WN1,WN2)、高要尼羅羅非魚群體(GN1,GN2)、無錫奧利亞羅非魚群體(WO1,WO2)以及番禺奧利亞羅非魚群體(PO1,PO2)內(nèi)雌雄之間遺傳距離較小,遺傳一致度較大,說明這些群體雌雄之間分化程度不高,詳細(xì)結(jié)果見表5。根據(jù)群體間Nei’s遺傳距離用NJ法得到聚類圖,所有群體聚為兩大支:WO1、WO2、PO1、PO2以及ZY1群體聚為一支,GN1、GN2、WY1、WN1、WN2、YY1以及YY2群體聚為另一支,見圖1。 表5 所有羅非魚群體之間的遺傳距離及遺傳一致度Tab.5 Genetic distance and genetic similarity coefficient among all tilapia populations 注:對角線以上為遺傳一致度,對角線以下為遺傳距離。 圖1 所有羅非魚群體的聚類分析圖Fig.1 Cluster dendrogram of all tilapia populations 2.2.4性別相關(guān)微衛(wèi)星位點的篩選 將初步篩選到的6個與性別相關(guān)的微衛(wèi)星標(biāo)記進一步在12個群體中擴增驗證后發(fā)現(xiàn):GM597與無錫尼羅羅非魚群體(WN1,WN2)、高要尼羅羅非魚群體(GN1,GN2)以及番禺奧利亞羅非魚群體(PO1,PO2)的性別顯著相關(guān)(P<0.05),鑒別率(鑒別率=群體內(nèi)可準(zhǔn)確鑒定性別的個體數(shù)/群體內(nèi)個體總數(shù)×100%)分別為39.1%、60.5%與7.5%;UNH898與高要尼羅羅非魚群體(GN1,GN2)的性別極顯著相關(guān)(P<0.01),鑒別率為71.1%。將GM597與UNH898聯(lián)合分析可100%鑒定兩個尼羅羅非魚群體的性別。將GM597與GM258在無錫奧利亞羅非魚群體和番禺奧利亞羅非魚群體中聯(lián)合分析,兩個群體的雌雄鑒別率分別為33.3%和40.0%。YY1群體中UNH898的基因型與無錫尼羅羅非魚群體及高要尼羅羅非魚群體均不相同,利用UNH898可以100%將YY1群體與兩個尼羅羅非魚群體區(qū)分開,部分結(jié)果見表6~8。 表4和表5列出了各個羅非魚群體的遺傳參數(shù),無錫尼羅、高要尼羅、無錫奧利亞以及番禺奧利亞各個群體內(nèi)部雌、雄之間各項遺傳參數(shù)(Na、Ne、Ho、He、I、PIC)差別不大,表明這些群體內(nèi)部雌雄之間遺傳多樣性基本相同。無錫尼羅、高要尼羅、無錫奧利亞以及番禺奧利亞各個群體內(nèi)部雌、雄之間的遺傳距離(0.029~0.198)相對較小,遺傳一致度相對較大(0.820~0.972),表明這些群體內(nèi)部雌雄之間基因組差異較小。董在杰等[18]利用RAPD技術(shù)對尼羅羅非魚和奧利亞羅非魚雌、雄群體進行分析,結(jié)果表明,奧利亞羅非魚雌雄群體的遺傳多樣性程度接近,而尼羅羅非魚雄性群體的遺傳多樣性要比雌性群體豐富,遺傳變異比雌性群體大,本研究中兩個奧利亞羅非魚群體的結(jié)果與之一致,而兩個尼羅羅非魚群體內(nèi)雌、雄之間的遺傳多樣性并無明顯差異,一方面可能是群體差異性導(dǎo)致的,另一方面可能是本研究所選標(biāo)記數(shù)較少的原因。無錫尼羅與高要尼羅雌雄群體的平均多態(tài)信息含量均達到了高度多態(tài)(0.580~0.729),說明兩個尼羅羅非魚群體的遺傳多樣性較高,這一結(jié)果與宋紅梅等[19]在構(gòu)建羅非魚DNA遺傳圖譜時得到的三個尼羅羅非魚群體平均多態(tài)信息含量為0.608(達到高度多態(tài))的結(jié)果相差不大,高于袁文華等[20]在奧尼羅非魚及其親本的微衛(wèi)星分析中尼羅群體的平均多態(tài)信息含量(0.469)。無錫奧利亞與番禺奧利亞雌雄群體的平均多態(tài)信息含量均為中度多態(tài)(0.307~0.354),說明兩個奧利亞羅非魚群體的遺傳多樣性中度偏低,這一結(jié)果略高于張庭等[21]對四個奧利亞群體進行微衛(wèi)星分析時得到的4個群體的平均多態(tài)信息含量為0.276(達到中度多態(tài))的結(jié)果。同種羅非魚不同群體間多態(tài)信息含量的差別可能是不同選育群體之間的遺傳差異性導(dǎo)致的。ZY1、WY1、YY1以及YY2型羅非魚是經(jīng)過激素誘導(dǎo)以及種內(nèi)和種間雜交后得到的羅非魚,目前微衛(wèi)星標(biāo)記在這些群體中研究較少,WY1以及YY2群體的平均多態(tài)信息含量分別為0.753和0.528(達到高度多態(tài)),說明這兩個群體遺傳多樣性較高,ZY1以及YY1型羅非魚的平均多態(tài)信息含量分別為0.486和0.275(達到中度多態(tài)),說明這兩個群體遺傳多樣性適中。劉志剛等[22]在利用線粒體DNA構(gòu)建羅非魚“粵閩1 號” 及其繁育群體的NJ 系統(tǒng)進化樹時發(fā)現(xiàn)YY1與YY2群體并沒有聚為一支,而在本研究中YY1與YY2群體在聚類分析中聚為一支,這一結(jié)果可能是線粒體DNA只能通過母系遺傳而微衛(wèi)星DNA可通過父系及母系共同遺傳的差異導(dǎo)致的。 表6 GM597與羅非魚性別的關(guān)聯(lián)分析Tab.6 Sex-related analysis between GM597 and tilapia 表7 UNH898與羅非魚性別的關(guān)聯(lián)分析Tab.7 Sex-related analysis between UNH898 and tilapia 表8 GM597與UNH898在兩個尼羅羅非魚群體中雌雄鑒別率統(tǒng)計Tab.8 Statistics on male and female identification rates of GM597 and UNH898 in two O.niloticus populations 本實驗采用兩種實驗方案對兩個尼羅羅非魚以及兩個奧利亞羅非魚群體進行性別相關(guān)微衛(wèi)星標(biāo)記的篩選。第一種實驗方案采用常規(guī)PCR擴增微衛(wèi)星位點,12%非變性聚丙烯酰胺凝膠電泳分離,篩選到與性別相關(guān)的標(biāo)記較多。第二種實驗方案采用降落PCR擴增微衛(wèi)星位點,毛細(xì)管電泳分離,篩選到與性別相關(guān)的標(biāo)記較少。造成這一差異的原因有兩個方面:一方面由于所擴增的微衛(wèi)星位點多態(tài)性較高,等位基因較為豐富,大量等位基因只相差2 bp,造成基因分型存在不同程度的誤差,另一方面?zhèn)鹘y(tǒng)的PCR和聚丙烯酰胺凝膠電泳相較于降落PCR和毛細(xì)管電泳在實際操作過程中不可控的因素更多,誤差更大。 采用兩種實驗方案均發(fā)現(xiàn)GM597與無錫尼羅羅非魚群體(WN1,WN2)、高要尼羅羅非魚群體(GN1,GN2)以及番禺奧利亞羅非魚群體的性別顯著相關(guān)(P<0.05),UNH898與高要尼羅羅非魚群體(GN1,GN2)的性別極顯著相關(guān)(P<0.01)。將GM597與UNH898聯(lián)合分析可100%鑒定兩個尼羅羅非魚群體的性別,利用UNH898可以100%將YY1群體與兩個尼羅羅非魚群體區(qū)分開。Eshel等[13,14]研究發(fā)現(xiàn)位于LG23內(nèi) Scaffold 101上長度為1.5 Mbp的區(qū)域是尼羅羅非魚的一個性別決定區(qū)域,UNH898與一個尼羅羅非魚家系的性別極顯著相關(guān),其276 bp的等位基因是雄性個體的一個標(biāo)記。本研究中篩選到的微衛(wèi)星標(biāo)記GM597與UNH898位于該性別決定區(qū)域內(nèi),因此GM597與UNH898可作為篩選YY1超雄魚的候選標(biāo)記。Sun等[15]篩選到一個與尼羅羅非魚性別相關(guān)的AFLP標(biāo)記SCAR-5位于上述性別決定區(qū)域附近,后續(xù)可以在該區(qū)域附近開發(fā)多種類型的分子標(biāo)記應(yīng)用到尼羅羅非魚的性別鑒定過程中。目前的研究顯示奧利亞羅非魚的性別決定區(qū)域位于LG1與LG3上[9,16],本研究發(fā)現(xiàn)位于LG23上的GM597與番禺奧利亞羅非魚群體(PO1,PO2)的性別顯著相關(guān)(P<0.05),但其雌雄鑒別率只有7.5%,將GM597與GM258在無錫奧利亞羅非魚群體(WO1,WO2)和番禺奧利亞羅非魚群體(PO1,PO2)中聯(lián)合分析,兩個群體的雌雄鑒別率分別為33.3%和40.0%,均低于50%,可能是樣本量少的原因,導(dǎo)致結(jié)果存在一定誤差。與Lee等[8,9]的研究結(jié)果相比本實驗中并沒有檢測到UNH995和UNH104與尼羅羅非魚群體的性別相關(guān),也沒有檢測到GM354與奧利亞羅非魚群體的性別相關(guān),這可能是由于不同選育群體之間的遺傳差異性導(dǎo)致的,此外本實驗篩選到的與羅非魚性別相關(guān)的微衛(wèi)星標(biāo)記數(shù)量較少,可能與選取的引物數(shù)量相對較少有關(guān),后續(xù)可以開發(fā)多種性別相關(guān)的分子標(biāo)記,應(yīng)用到羅非魚的性別鑒定過程中。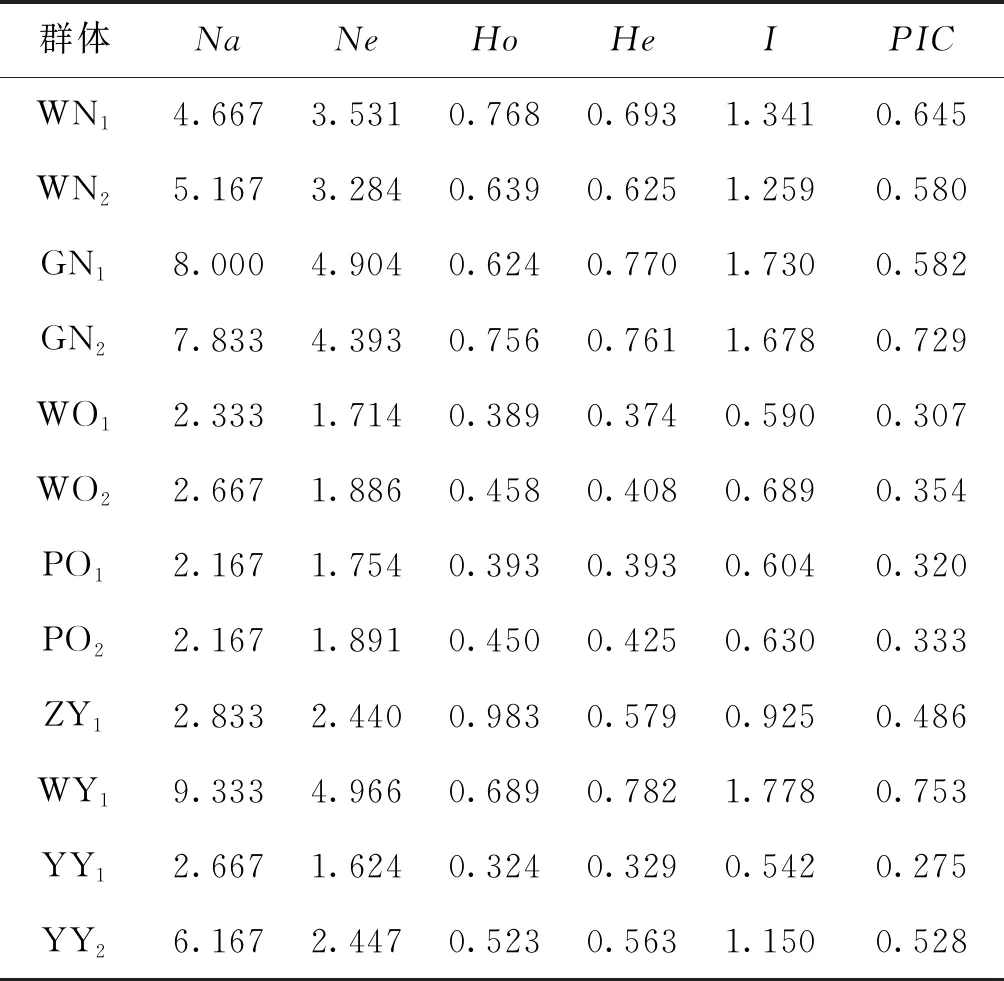
3 討論
3.1 群體內(nèi)以及群體間的遺傳差異

3.2 性別相關(guān)微衛(wèi)星標(biāo)記的分析
