馬鈴薯根際與非根際土壤微生物群落結(jié)構(gòu)及多樣性特征
2020-02-21 04:19:50葛應(yīng)蘭孫廷
生態(tài)環(huán)境學(xué)報(bào) 2020年1期
關(guān)鍵詞:植物
葛應(yīng)蘭,孫廷
南陽(yáng)農(nóng)業(yè)職業(yè)學(xué)院農(nóng)業(yè)科學(xué)系,河南 南陽(yáng) 473007
土壤微生物是土壤中元素循環(huán)、有機(jī)質(zhì)分解、土壤礦化等地球化學(xué)循環(huán)過(guò)程中的重要參與者,也是土壤物質(zhì)循環(huán)的主要驅(qū)動(dòng)者,參與并維持農(nóng)業(yè)土壤的健康和生產(chǎn)力(Leff et al.,2015;Carini et al.,2017;Lange et al.,2015)。土壤微生物的多樣性對(duì)土壤生態(tài)系統(tǒng)的功能、可持續(xù)性和完整性至關(guān)重要,近年來(lái)越來(lái)越多的研究表明,較高的微生物多樣性可以增強(qiáng)生態(tài)系統(tǒng)和微生物功能的穩(wěn)定性(Lu et al.,2015;Deng et al.,2016)。根際土壤是直接受植物根系和分泌物影響的土壤區(qū)域,是土壤微生物與植物相互作用的重要場(chǎng)所(Lu et al.,2015)。根際土壤微生物與根系之間存在適應(yīng)性協(xié)同進(jìn)化關(guān)系,并形成植物-微生物之間的互惠關(guān)系(Wang et al.,2016;Bakker et al.,2015;Walters et al.,2018)。植物通過(guò)根系活動(dòng)改變根際土壤的養(yǎng)分含量及其他土壤理化性質(zhì),改變根際微生物群落的組成,使得根際與非根際土微生物群落組成和多樣性上具有明顯的差異(Wang et al.,2016;Bakker et al.,2015)。根際微生物對(duì)植物生長(zhǎng)具有重要的作用,促植物生長(zhǎng)細(xì)菌(PGPR)通過(guò)自身代謝活動(dòng)將土壤營(yíng)養(yǎng)物質(zhì)分解、轉(zhuǎn)化為植物可吸收利用的形態(tài),促進(jìn)植物生長(zhǎng),部分細(xì)菌分泌的激素等物質(zhì)可提高植物的適應(yīng)性(Walters et al.,2018)。根際土壤微生物的群落結(jié)構(gòu)受植物影響,如植物種類(lèi)、生長(zhǎng)階段、健康狀況,另外,土壤理化性質(zhì)對(duì)微生物群落也具有重要的影響,如土壤營(yíng)養(yǎng)狀況、水分和pH、溫度(Lladó et al.,2018;Lu et al.,2015)。
馬鈴薯是中國(guó)農(nóng)業(yè)生產(chǎn)中主栽作物之一,近年來(lái),隨著栽培面積持續(xù)擴(kuò)大,馬鈴薯種植的土壤問(wèn)題日漸突出,緩解或消除馬鈴薯土壤貧瘠等問(wèn)題障礙迫在眉睫(張立超等,2017;王麗紅等,2016)。已有研究表明,多年種植馬鈴薯后會(huì)對(duì)土壤微生物產(chǎn)生抑制作用,使土壤微生物總數(shù)、細(xì)菌數(shù)量減少,真菌數(shù)量增加(Uroz et al.,2016;Hou et al.,2015)。隨著種植年限的增加,細(xì)菌種類(lèi)及數(shù)量不斷減少,特別是芽孢桿菌和放線(xiàn)菌等種類(lèi)的改變,致使土壤微生物群落多樣性水平呈下降趨勢(shì)(孫夢(mèng)媛等,2017)。因此,加深對(duì)土壤微生物群落結(jié)構(gòu)、功能的了解,能夠闡明馬鈴薯種植后土壤微生物的影響機(jī)制。本研究利用Illumina-MiSeq高通量測(cè)序技術(shù)對(duì)馬鈴薯根際與非根際土壤中細(xì)菌的16S rDNA基因V3-V4區(qū)片段和真菌18S rDNA基因V4區(qū)片段進(jìn)行了測(cè)序,研究馬鈴薯根際與非根際土壤微生物群落多樣性及其與土壤養(yǎng)分之間的關(guān)系,為研究馬鈴薯健康種植提供理論數(shù)據(jù)。
1 材料與方法
1.1 樣品采集
供試馬鈴薯為莊薯3號(hào),種植面積為50 m ×50 m。于2018年馬鈴薯盛花期(4月中)采集根際土壤樣本,選擇3塊50 m ×50 m的重復(fù)樣地,每個(gè)樣地選5株長(zhǎng)勢(shì)一致的馬鈴薯植株,采用抖根法(Lu et al.,2015)收集根際土壤,在根系周?chē)?—5 cm的位置采集非根際土,同一樣方內(nèi)的土樣混合均勻作為一個(gè)混合樣,所有土壤樣品立即帶回實(shí)驗(yàn)室,4 ℃冰箱保存。共采集15個(gè)根際樣品和15個(gè)非根際樣品。混勻的土樣一部分風(fēng)干保存,測(cè)定土壤理化性質(zhì),另一部分新鮮土壤用于提取微生物總DNA進(jìn)行高通量測(cè)序。
1.2 土壤理化性質(zhì)
對(duì)于pH值、有機(jī)碳SOC、全氮TN的測(cè)定,分別借助于pH測(cè)量計(jì)、容重法及硝化法進(jìn)行;通過(guò)電導(dǎo)儀測(cè)定電導(dǎo)率 EC,并通過(guò)其估算含鹽量;對(duì)于總磷和速效磷采用鉬銻抗比色法,速效氮采用光度計(jì)法(Zhalnina et al.,2018)。
1.3 土壤總DNA提取
利用 Fast DNA spin kit for soil試劑盒(MP BIO,Inc.,Irvine,CA,USA)進(jìn)行 DNA提取,根據(jù)制造商提供的說(shuō)明進(jìn)行操作。洗脫后總 DNA體積為50 μL。利用1%的瓊脂糖凝膠泳提取總DNA(電泳條帶的單一性),同時(shí)用NanoDrop ND-1000微光分光光度計(jì)測(cè)定其濃度。
1.4 土壤微生物基因組DNA的提取
吸取1 mL根際土壤溶液至2 mL無(wú)菌離心管中,于4 ℃、10000 g離心30 s,棄上清,所得沉淀用于根際土壤微生物基因組DNA提取;對(duì)非根際土,用無(wú)菌藥匙取大約0.2 g土壤(濕重),加入2 mL無(wú)菌離心管中,用于提取基因組DNA。利用OMEGA試劑盒E.Z.N.ATM Mag-Bind Soil DNA Kit試劑盒(OMEGA)提取基因組DNA,利用1%瓊脂糖凝膠檢測(cè) DNA的長(zhǎng)度和完整性,利用NanoDrop檢測(cè)DNA的濃度和純度。
1.5 DNA擴(kuò)增及測(cè)序
采用兩輪法對(duì)細(xì)菌16S rDNA V3-V4區(qū)進(jìn)行擴(kuò)增,第一輪擴(kuò)增利用融合了barcode序列的通用引物進(jìn)行,上游引物 341F:ccctacacgacgctcttccgatctg(barcode)cctacgggnggcwgcag,下游引物 805R:gactggagttcctt ggcacccgagaattccagactachvgggtatctaatcc;真菌 18S rDNA擴(kuò)增,第一輪擴(kuò)增利用融合了barcode 序列的通用引物進(jìn)行,上下游引物序列:NS1:cctacacgacgctctt ccgatctn(barcode)gtagtcatatgcttgtctc,F(xiàn)ung:gactggagtt ccttggcacccgagaattccaattccccgttacccgt tg。反應(yīng)體系30 μL,包含15 μL 2×Taq master Mix(Thermo),引物(10 μmol·L-1)各 1 μL,模板 DNA 20 ng。擴(kuò)增條件:94 ℃3 min,94 ℃ 30 s,45 ℃ 20 s,65 ℃ 30 s,擴(kuò)增 5個(gè)循環(huán);94 ℃ 20 s,55℃ 20 s,72 ℃ 30 s,擴(kuò)增20循環(huán),72 ℃延伸5 min。第二輪擴(kuò)增使用Illumina橋式PCR兼容引物,以第一輪的PCR產(chǎn)物為模板。反應(yīng)體系同上。擴(kuò)增條件:95 ℃ 30 s,95 ℃ 15 s,55 ℃ 15 s,72 ℃ 30 s,擴(kuò)增5個(gè)循環(huán),72 ℃延伸5 min。PCR產(chǎn)物經(jīng)磁珠法回收并定量。將每個(gè)重復(fù)個(gè)體的 PCR產(chǎn)物等量混合,每份混合樣品包含相應(yīng)的全部重復(fù)個(gè)體,將混合樣品送至生工(上海)生物工程有限公司,利用Illumina-MiSeq平臺(tái)進(jìn)行高通量測(cè)序(Uroz et al.,2016;Hou et al.,2015)。
1.6 數(shù)據(jù)分析
測(cè)序原始數(shù)據(jù)去除引物接頭序列、去除各低質(zhì)量堿基(Phred Quality Score=20)后進(jìn)行拼接,舍棄長(zhǎng)度短于200 bp的序列,去除非特異性擴(kuò)增序列及嵌合體后,得到每個(gè)樣本的有效序列數(shù)據(jù)。以97%為劃定閾值,對(duì)16S和18S序列劃分操作分類(lèi)單元(operational taxonomic unit,OTU)。根據(jù)樣本OTUs組成之間的weighted unifrac 距離矩陣,采用主成分分析(PCA)分析微生物群落之間的相似性;Pearson相關(guān)系數(shù)檢驗(yàn)微生物多樣性與土壤養(yǎng)分之間的相關(guān)性;利用Canoco 4.5軟件對(duì)土壤養(yǎng)分和微生物群落多樣性指數(shù)進(jìn)行冗余分析(Reundancy analysis,RDA)(Shi et al.,2016)。
2 結(jié)果分析
2.1 土壤養(yǎng)分
由表 1可知,馬鈴薯根際土壤 pH平均值為5.69,顯著低于非根際(P<0.05);差異性檢驗(yàn)表明,根際土壤電導(dǎo)率、有機(jī)碳、全氮速效氮和速效磷均顯著高于非根際(P<0.05),而根際土壤全磷與非根際差異不顯著(P>0.05)。

表1 馬鈴薯根際與非根際土壤養(yǎng)分Table 1 Soil nutrients of potato in rhizosphere and non-rhizosphere soil
2.2 土壤微生物群落多樣性
由表2可知,馬鈴薯根際土壤細(xì)菌豐富度指數(shù)、Shannon-Wiener 指數(shù)、ACE、Chao 1指數(shù)均顯著高于非根際(P<0.05);而根際土壤細(xì)菌覆蓋度、均勻度和 Simpson指數(shù)與非根際差異不顯著(P>0.05)。由表3可知,馬鈴薯根際土壤真菌豐富度指數(shù)、Shannon-Wiener指數(shù)、ACE、Chao 1指數(shù)均顯著高于非根際(P<0.05);而根際土壤真菌覆蓋度、均勻度和Simpson指數(shù)與非根際差異不顯著(P>0.05)。
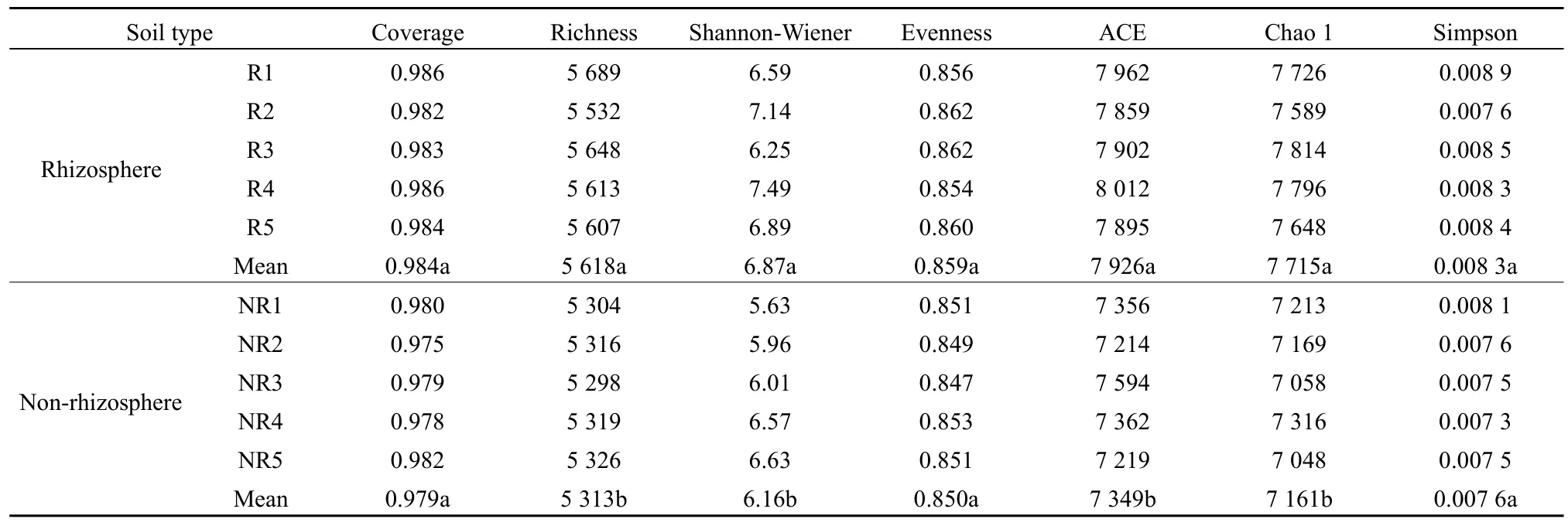
表2 馬鈴薯根際與非根際土壤細(xì)菌多樣性Table 2 Bacterial diversity in rhizosphere and non-rhizosphere soil
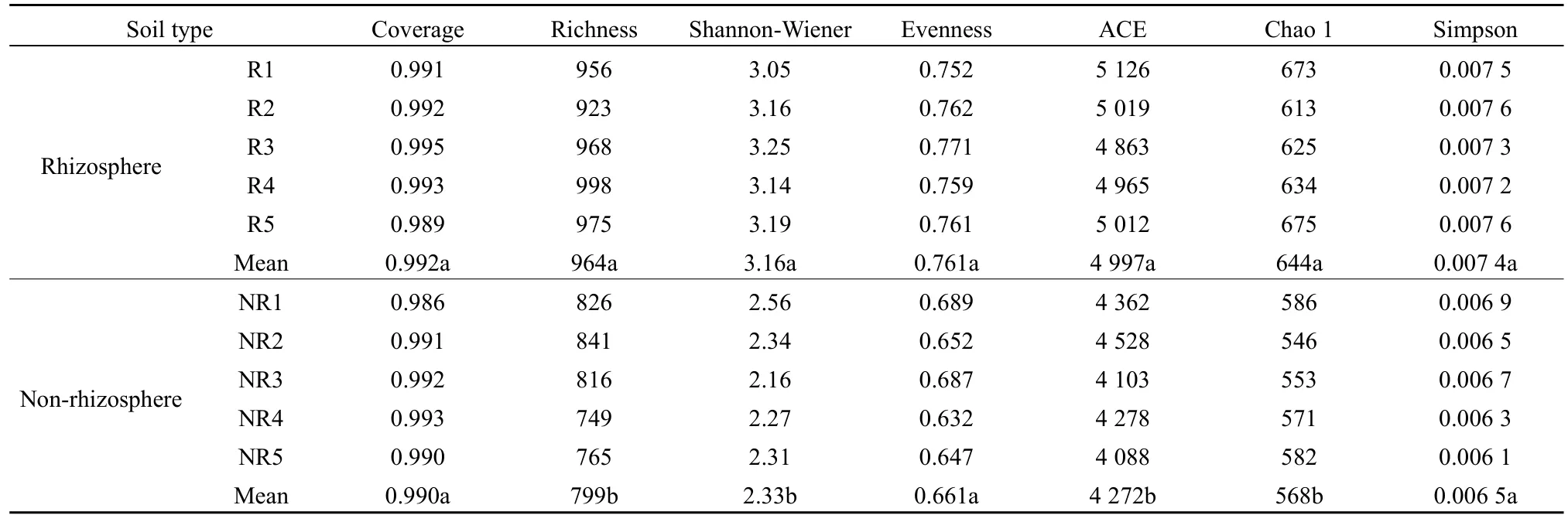
表3 馬鈴薯根際與非根際土壤真菌多樣性Table 3 Fungal diversity in rhizosphere and non-rhizosphere soil
2.3 土壤微生物群落組成
由圖1可知,馬鈴薯根際土壤細(xì)菌群落中,優(yōu)勢(shì)類(lèi)群主要是變形菌門(mén)Proteobacteria(35.98%)、酸桿菌門(mén) Acidobacteria(27.03%)和芽單胞菌門(mén)Gemmatimonadetes(9.14%),還有浮霉菌門(mén)Planctomycetaceae(4.03%)、放線(xiàn)菌門(mén)Actinobacteria(3.02%)、擬桿菌門(mén)Bacteroidetes(1.56%)、后壁菌門(mén) Firmicutes(3.89%)、綠彎菌門(mén) Chloroflexi(6.18%)、疣微菌門(mén) Verrucomicrobia(3.01%)。非根際土壤細(xì)菌群落中,優(yōu)勢(shì)類(lèi)群主要是變形菌門(mén)(46.02%)、酸桿菌門(mén)(23.17%)和芽單胞菌門(mén)(12.06%)。其中根際土壤細(xì)菌酸桿菌門(mén)相對(duì)豐度高于非根際,變形菌門(mén)相對(duì)豐度低于非根際。
馬鈴薯根際土壤真菌群落中,優(yōu)勢(shì)類(lèi)群主要是子囊菌門(mén) Ascomycota(53.03%)和擔(dān)子菌門(mén)Basidiomycota(26.17%),還包括結(jié)合菌門(mén)Zygomycota(3.26%)、壺菌門(mén) Chytridiomycota(0.58%)、新麗鞭毛菌門(mén) Neocallimastigomycota(7.23%)、球囊菌門(mén)Glomeromycota(3.42%)、芽枝菌門(mén) Blastocladiomycota(2.19%)。非根際土壤真菌群落中,優(yōu)勢(shì)類(lèi)群主要是子囊菌門(mén)(50.16%)和擔(dān)子菌門(mén)(30.18%)。主成分分析(PCA,圖2)表明:馬鈴薯根際土壤和非根際土壤比較,細(xì)菌群落、真菌群落都具有很好的相似性,并且根際和非根際的細(xì)菌群落、真菌群落都產(chǎn)生明顯的分離效應(yīng)。

圖1 土壤細(xì)菌(A)和真菌(B)群落組成Fig. 1 Bacterial (A) and fungal (B) community composition in rhizosphere and non-rhizosphere soil
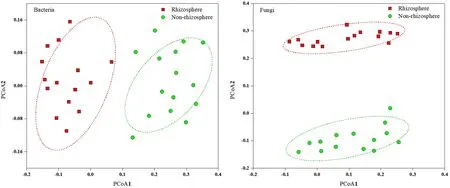
圖2 基于Weighted unifrac距離的土壤微生物群落的主坐標(biāo)分析(PCoA)Fig. 2 Principal coordinate analysis (PCoA) of soil microbial communities based on Weighted unifrac distance
2.4 土壤養(yǎng)分與細(xì)菌群落結(jié)構(gòu)的相關(guān)性
相關(guān)性分析表明(表 4),馬鈴薯土壤細(xì)菌和真菌Coverage、ACE與土壤養(yǎng)分均沒(méi)有顯著相關(guān)性(P>0.05);土壤pH與土壤細(xì)菌和真菌多樣性呈負(fù)相關(guān),土壤電導(dǎo)率和全磷與土壤細(xì)菌和真菌多樣性均沒(méi)有顯著相關(guān)性(P>0.05)。
土壤有機(jī)碳與細(xì)菌 Richness、Shannon呈極顯著正相關(guān)(P<0.01),與細(xì)菌Chao 1和Simpson呈顯著正相關(guān)(P<0.05);土壤有機(jī)碳與真菌Shannon呈極顯著正相關(guān)(P<0.01),與真菌Richness、Chao 1和Simpson呈顯著正相關(guān)(P<0.05)。土壤全氮與細(xì)菌 Richness、Shannon呈極顯著正相關(guān)(P<0.01),與細(xì)菌Chao 1和Simpson呈顯著正相關(guān)(P<0.05);土壤全氮與真菌Shannon呈極顯正相關(guān)(P<0.01),與真菌 Richness、Chao 1 和 Simpson呈顯著的正相關(guān)(P<0.05)。土壤速效氮與Richness、Shannon、Evenness、Chao 1和Simpson呈顯著正相關(guān)(P<0.05),與真菌Shannon、Evenness、Chao 1和Simpson呈顯著正相關(guān)(P<0.05)。土壤速效磷與細(xì)菌Shannon、Chao 1和Simpson呈極顯著正相關(guān)(P<0.01),與 Evenness呈顯著正相關(guān)(P<0.05);土壤速效磷與真菌Shannon、Evenness、Chao 1和Simpson呈顯著正相關(guān)(P<0.05)。
2.5 土壤微生物群落與土壤養(yǎng)分的關(guān)系
馬鈴薯根際土壤細(xì)菌和真菌群落與土壤環(huán)境因子的冗余分析結(jié)果見(jiàn)表 5。蒙特卡羅置換檢驗(yàn)結(jié)果顯示,第一典范軸P值為0.469(F=32.16),所有典范軸的P值為0.902,表明該排序模型的解釋變量(即土壤環(huán)境因子)與響應(yīng)變量(即土壤細(xì)菌和真菌群落結(jié)構(gòu))的變化關(guān)系較為顯著。
由圖3可知,對(duì)于土壤細(xì)菌群落,RDA前兩個(gè)排序軸的特征值分別為0.6498和0.2147,分別解釋了64.98%和21.47%的細(xì)菌多樣性變化。所選的7個(gè)土壤環(huán)境因子共解釋了86%的總特征值,對(duì)馬鈴薯根際土壤細(xì)菌多樣性有顯著影響。其中對(duì)土壤細(xì)菌多樣性影響較大的有SOC和TN,而pH對(duì)土壤細(xì)菌多樣性影響為負(fù)。
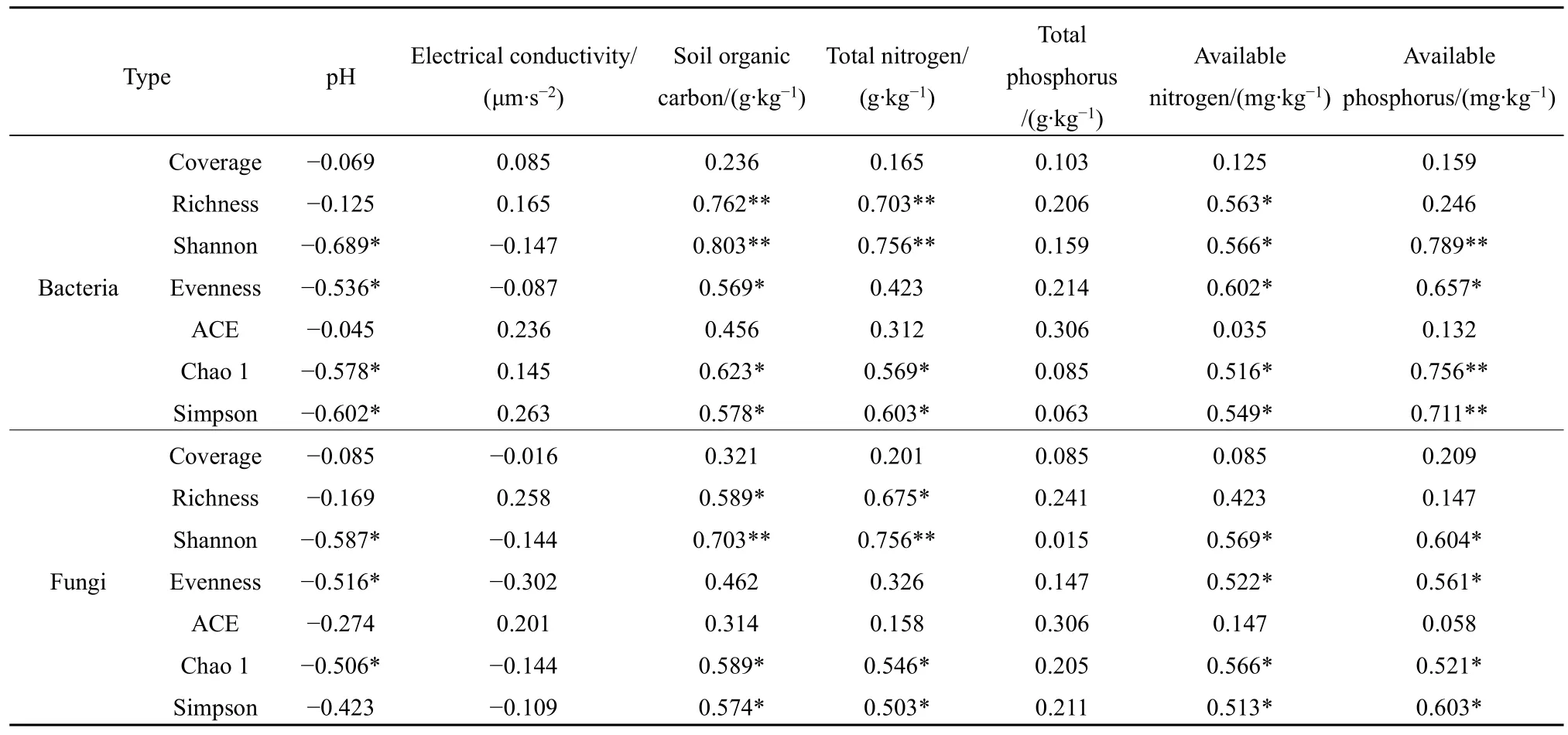
表4 土壤養(yǎng)分與土壤細(xì)菌和真菌多樣性相關(guān)性Table 4 Soil nutrients related to the diversity of soil bacteria and fungi
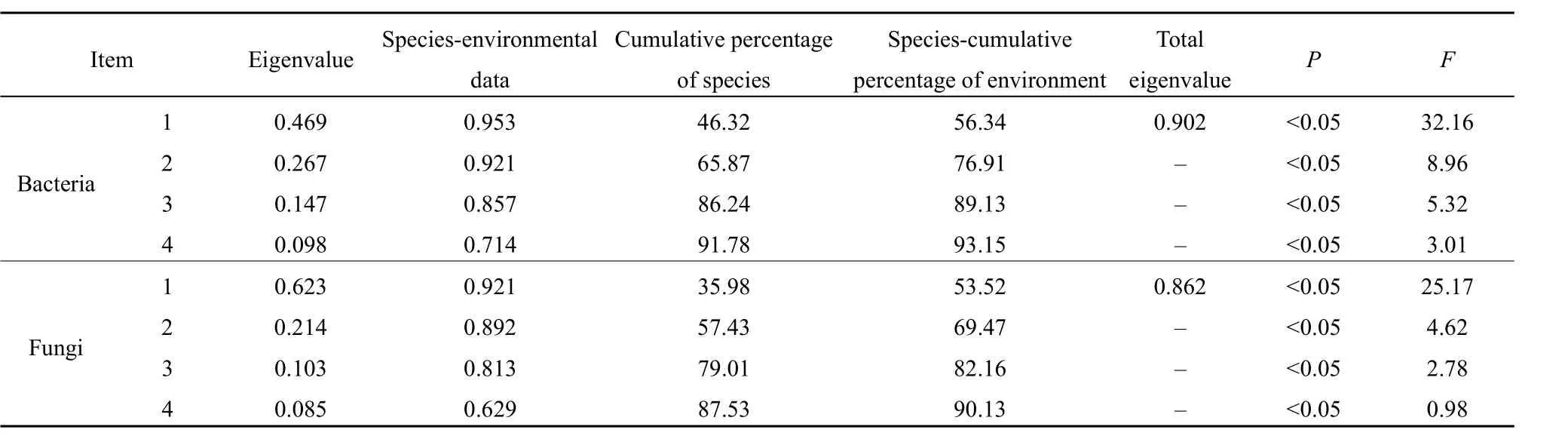
表5 冗余分析結(jié)果Table 5 Redundant analysis results
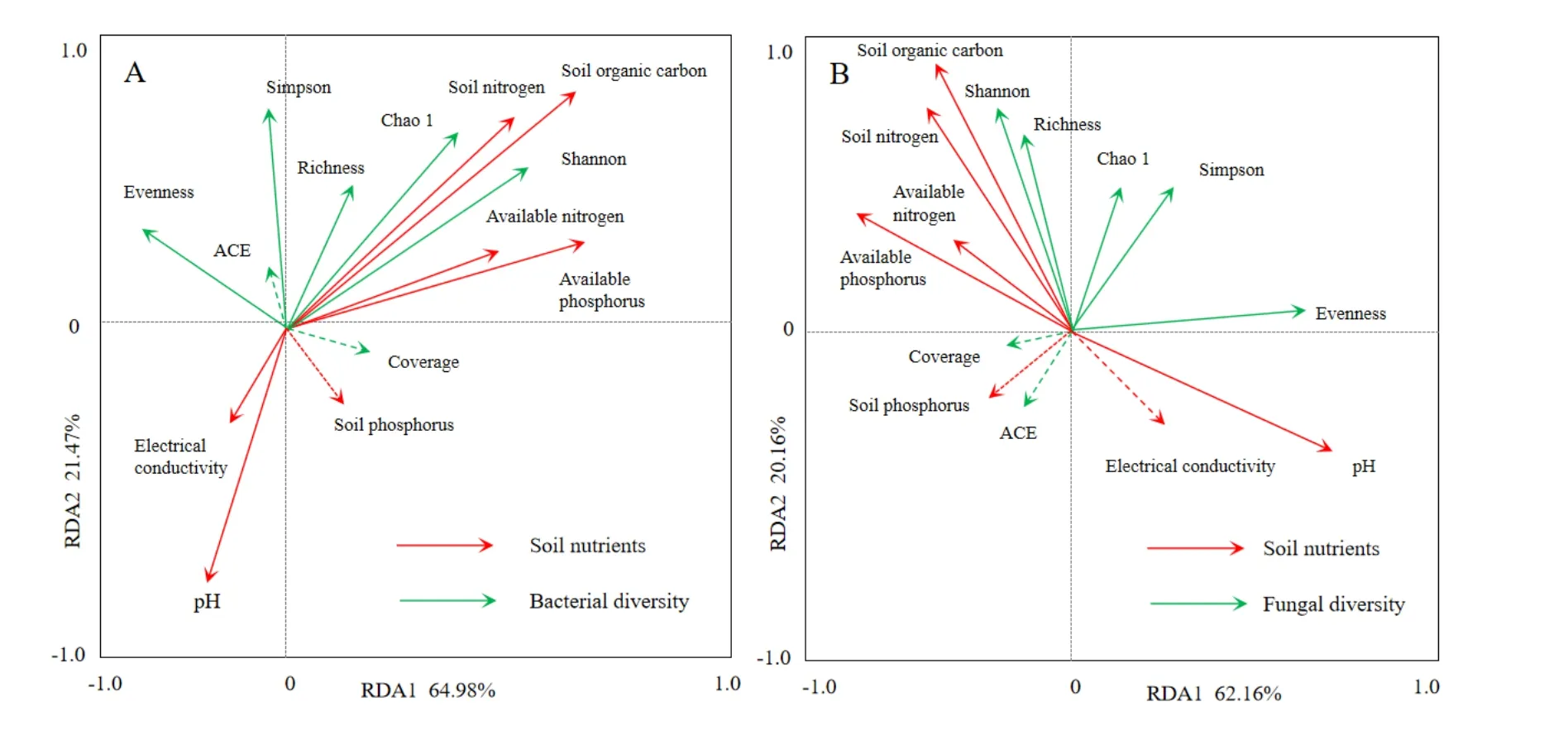
圖3 土壤細(xì)菌(A)和真菌(B)多樣性指數(shù)與環(huán)境因子的關(guān)系Fig. 3 Soil bacterial diversity index(A) and fungal diversity index (B) related to environmental factors
對(duì)于土壤真菌群落,RDA前兩個(gè)排序軸的特征值分別為0.6216和0.2016,分別解釋了62.16%和20.16%的真菌多樣性變化。所選的7個(gè)土壤環(huán)境因子共解釋了82%的總特征值,對(duì)馬鈴薯根際土壤真菌多樣性有顯著影響。其中對(duì)土壤真菌多樣性影響較大的有SOC和TN,而pH對(duì)土壤真菌多樣性影響為負(fù)。
3 討論
本研究中馬鈴薯非根際和根際土壤中細(xì)菌豐富度高,而真菌數(shù)量相對(duì)較少,這與土壤中微生物以細(xì)菌為主一致(Maarastawi et al.,2018;Whitman et al.,2017)。根際細(xì)菌多樣性高于非根際細(xì)菌,而根際真菌多樣性卻低于非根際土壤。根際土壤中營(yíng)養(yǎng)物質(zhì)高于非根際土壤,有利于細(xì)菌的繁殖故豐富度和多樣性較高。相關(guān)性分析也表明細(xì)菌多樣性和組成與SOC、TN呈正相關(guān),與以往研究相一致(Liang et al.,2016;Deng et al.,2015);而土壤真菌多為病原菌,不利于植物生長(zhǎng),可能被植物根系主動(dòng)過(guò)濾,導(dǎo)致根際真菌多樣性與非根際差異不顯著(Taschen et al.,2017)。根際細(xì)菌、真菌群落組成與非根際土壤存在明顯的差異,PCoA結(jié)果也表明非根際土壤與根際微生物群落分離明顯,非根際土壤群落之間,以及根際土壤群落之間各具有較高的相似性。
門(mén)水平上,馬鈴薯根際和非根際土壤細(xì)菌群落中,優(yōu)勢(shì)類(lèi)群主要是變形菌門(mén)、酸桿菌門(mén)和芽單胞菌門(mén),還包括浮霉菌門(mén)、放線(xiàn)菌門(mén)、擬桿菌門(mén)、后壁菌門(mén)、綠彎菌門(mén)、疣微菌門(mén),其中根際土壤細(xì)菌酸桿菌門(mén)相對(duì)豐度高于非根際,變形菌門(mén)相對(duì)豐度低于非根際。根際和非根際土壤真菌群落中,優(yōu)勢(shì)類(lèi)群主要是子囊菌門(mén)和擔(dān)子菌門(mén),還包括結(jié)合菌門(mén)、壺菌門(mén)、新麗鞭毛菌門(mén)、球囊菌門(mén)、芽枝菌門(mén)。根際與非根際菌群的差異,一方面是因?yàn)楦H土壤養(yǎng)分的含量高于非根際土,pH值低于非根際土,利于某些特定類(lèi)群的增殖和豐度的提高(Taschen et al.,2017)。RDA結(jié)果表明,細(xì)菌和真菌群落結(jié)構(gòu)與土壤SOC和TN呈正相關(guān)關(guān)系,而細(xì)菌和真菌群落結(jié)構(gòu)與土壤pH呈負(fù)相關(guān)關(guān)系。總體上,根際土壤細(xì)菌群落多樣性高于非根際,主要是由于根際土壤pH的酸化導(dǎo)致土壤養(yǎng)分含量較高,土壤微生物多樣性較高(Li et al.,2016;Stewart et al.,2017;Zhang et al.,2015)。另一方面,是植物從自身代謝和健康出發(fā),由根系主動(dòng)選擇形成的,gamma變形菌包含許多促植物生長(zhǎng)細(xì)菌(PGPR),如具有固氮作用的芽孢桿菌屬、固氮菌屬,在生產(chǎn)和釋放植物激素和土壤源植物病原菌(如真菌)的生物防治中有重要作用。alpha變形菌包含許多植物共生固氮菌,如Rhizobium、Rickettsia;delta變形菌的一些物種,如Desulfovibrio、Geobacter、Bdellovibrio等,對(duì)土壤硫循環(huán)具有重要作用,對(duì)植物生長(zhǎng)有重要作用,因此在根際土壤中的豐富度明顯高于非根際土壤;alpha和delta變形菌的豐度升高是在根際土壤中變形菌門(mén)的豐度顯著高于非根際土的主要原因(Shen et al.,2015;Mellado-Vázquez et al.,2016)。
4 結(jié)論
馬鈴薯根際土壤pH顯著低于非根際,根際土壤養(yǎng)分、土壤細(xì)菌和真菌群落多樣性(豐富度指數(shù)、Shannon-Wiener指數(shù)、ACE、Chao 1指數(shù))均顯著高于非根際(P<0.05),而根際土壤全磷與非根際差異不顯著(P>0.05),且根際優(yōu)勢(shì)群落相對(duì)豐度較高。土壤微生物群落多樣性和組成在不同生境之間的差異與土壤理化性質(zhì)密切相關(guān),生境不同,根際微生物群落不同。研究發(fā)現(xiàn)馬鈴薯土壤根際和非根際細(xì)菌和真菌群落具有很好的相似性,并且根際和非根際細(xì)菌群落產(chǎn)生明顯的分離效應(yīng)。土壤環(huán)境因子對(duì)馬鈴薯土壤細(xì)菌和真菌多樣性有顯著影響,其中對(duì)土壤細(xì)菌和真菌多樣性影響較大的有 SOC和TN,而pH對(duì)土壤細(xì)菌和真菌多樣性影響為負(fù)。由此可知,在馬鈴薯的種植過(guò)程中,土壤pH值對(duì)根際微生物群落多樣性起著重要的調(diào)節(jié)作用。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語(yǔ)數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫(huà)刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
 生態(tài)環(huán)境學(xué)報(bào)2020年1期
生態(tài)環(huán)境學(xué)報(bào)2020年1期
- 生態(tài)環(huán)境學(xué)報(bào)的其它文章
- 半胱氨酸強(qiáng)化針鐵礦類(lèi)芬頓反應(yīng)降解2, 4-二氯苯酚及影響因素
- 市政污泥與生活垃圾摻燒的重金屬排放特征與風(fēng)險(xiǎn)
- 三峽庫(kù)區(qū)香溪河沉積物重金屬含量分布及風(fēng)險(xiǎn)評(píng)價(jià)
- 廣東紅樹(shù)林沉積物有毒金屬分布及生態(tài)風(fēng)險(xiǎn)評(píng)價(jià)
- 柴油尾氣顆粒對(duì)秀麗隱桿線(xiàn)蟲(chóng)慢性毒性效應(yīng)的初步研究
- 氣象條件對(duì)西安市夏季和冬季近地面大氣環(huán)境污染特征影響