敲減STAT3基因表達對胰腺祖細胞增殖的影響*
2020-03-03 07:27:48李旭艷翟文君馬明君陳振寶孫玉林李永芹
中國病理生理雜志 2020年2期
關鍵詞:檢測
李旭艷, 翟文君, 馬明君, 陳振寶, 孫玉林, 趙 娟, 李永芹
(1嶺南師范學院生命科學與技術學院, 廣東 湛江 524000; 2東北林業大學生命學院, 黑龍江 哈爾濱 150040)
隨著人們生活方式的改變和生活環境的變化,糖尿病和胰腺炎等胰腺相關疾病的發病率呈逐年上升趨勢,嚴重危害人類健康和生活質量。糖尿病患者表現為胰島β細胞受損或胰島素抵抗,胰腺炎患者的胰腺腺泡細胞凋亡或壞死。補充受損或功能異常的胰島β細胞或腺泡細胞將是治療這類疾病的根本途徑。終末分化胰島細胞和胰腺腺泡細胞均起源于胚胎發育早期胰腺干細胞,研究發現,成體胰腺含有1%的胰腺干/祖細胞,激活成體胰腺干/祖細胞增殖分化,獲得終末分化胰腺內、外分泌細胞,是胰腺疾病治療的新希望。
信號轉導及轉錄激活因子3(signal transducer and activator of transcription 3,STAT3)是重要的信號轉導因子,參與干擾素、白細胞介素和生長因子等多種細胞因子的信號轉導過程,是白細胞介素6/Janus 激酶(interleukin-6/Janus kinase,IL-6/JAK)和表皮生長因子受體(epidermal growth factor receptor,EGFR)等酪氨酸激酶信號通路效應分子,STAT3被上游的蛋白激酶磷酸化,形成二聚體,進入細胞核內調控促增殖、抗凋亡、免疫反應等基因表達,調控細胞生長、分化、遷移、凋亡、自噬、免疫以及代謝等重要生命活動。正常生理條件下STAT3低表達或短暫性激活,而在胰腺疾病發生發展過程中STAT3被活化并持續激活。被激活的STAT3促進胰腺癌細胞異常增殖、轉移、生成新生血管[1],炎癥在胰腺癌的進展過程中扮演重要角色,能顯著加速胰腺上皮腫瘤癌變,STAT3作為一種急性期反應因子,參與調控胰腺炎和胰腺纖維化進程[2],另外,STAT3參與糖尿病腎病、糖尿病心肌病和糖尿病視網膜病變的發生和發展的過程[3-5]。阻斷STAT3信號通路,能改善胰腺癌、胰腺炎和糖尿病并發癥癥狀,STAT3信號通路有可能成為治療胰腺疾病的藥物靶點。研究發現,STAT3對腫瘤干細胞、胚胎干細胞和成體干細胞的增殖、自我更新和重編程有重要調控作用。STAT3促進牙髓干細胞增殖、抑制分化[6],促進甲狀腺癌干細胞細胞集落形成[7],維持慢性髓細胞性白血病干細胞的特性[8],STAT3是否調控胰腺干/祖細胞的功能尚未報道。本研究以小鼠胰腺祖細胞為研究對象,沉默STAT3基因表達,檢測STAT3基因對胰腺祖細胞增殖的影響,探究胰腺祖細胞增殖的機制,將為利用胰腺干/祖細胞開展疾病治療提供實驗依據。
材 料 和 方 法
1 細胞
小鼠成體胰腺祖細胞為東北林業大學發育生物學實驗室提供[9]。
2 主要試劑
DMEM/F12培養液、胰蛋白酶、雙抗和L-谷氨酰胺購自HyClone;B-27細胞培養添加劑和胎牛血清(fetal bovine serum,FBS)購自Gibco;細胞培養級別重組人胰島素和表皮生長因子(epidermal growth factor,EGF)購自Becton, Dickinson and Company;細胞轉染脂質體Lipofectamine RNAiMAX購自Invitrogen;總RNA提取試劑Tripure和實時熒光定量PCR試劑RT-qPCR SYRB Green購自Roche;脫氧核糖核酸酶I(deoxyribonuclease I,DNaseI)和核糖核酸酶抑制劑(ribonuclease inhibitor,RI)購自TaKaRa;反轉錄試劑盒、抗β-肌動蛋白(β-actin)抗體(HC201-02)和辣根過氧化物酶標記的II抗(HS201-01)購自北京全式金生物技術有限公司;Cell Counting Kit-8(CCK8)試劑盒購自Dojindo Labratories;細胞周期檢測試劑盒、BCA蛋白濃度測定試劑盒和RIPA蛋白裂解液購自碧云天生物技術研究所;抗STAT3抗體(cst.9139)和抗細胞周期蛋白D2(cyclin D2,CCND2)抗體(cst. 3741)購自Cell Signaling Technology;β-巰基乙醇、過硫酸銨、丙烯酰胺、甲基雙丙烯酰胺和四甲基乙二胺購自Sigma;吐溫20、甲醇等常規生化試劑購自天津天大化學試劑廠。靶向STAT3的小干擾RNA[STAT3small interfering RNA (siRNA),siSTAT3][10]和對照siRNA(negative control,NC)由吉瑪基因股份有限公司合成,序列見表1。qPCR引物由金唯智基因公司合成,序列見表2。

表1 siRNAs序列
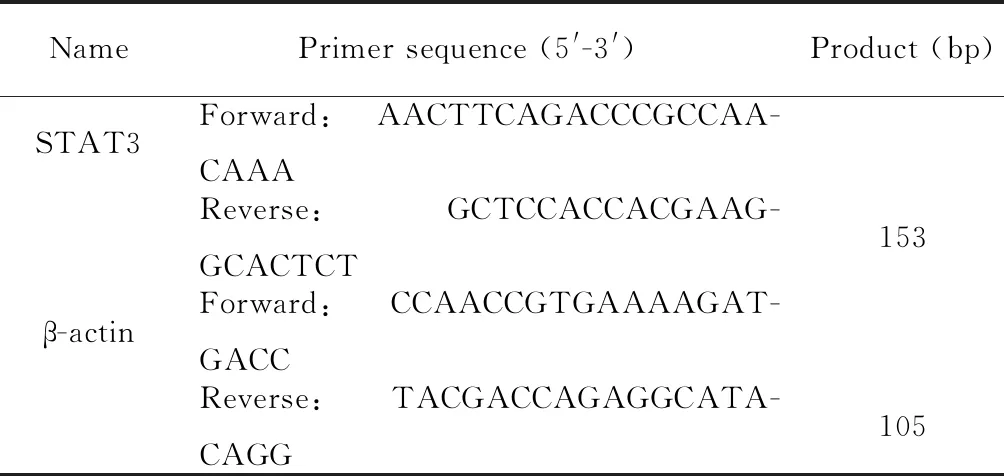
表2 qPCR引物序列
3 主要方法
3.1細胞培養 傳代胰腺祖細胞用10%貼壁培養液過夜培養,次日早更換2%生長培養液,置于37 ℃、5.0% CO2飽和濕度的培養箱中培養,每3 d更換生長培養液。貼壁培養液成分:含10% FBS、1%雙抗和1% L-谷氨酰胺的DMEM/F12培養液。生長培養液成分:含2% FBS、1%雙抗、1% L-谷氨酰胺、2% B-27、50 μmol/L β-巰基乙醇、10 mg/L重組人胰島素和20 μg/L EGF的DMEM/F12培養液。
3.2細胞轉染 取培養8代的胰腺祖細胞,96 孔和12 孔細胞培養板每孔分別接種3×103和1×105個細胞,生長培養液培養24 h后,細胞40%匯合后,每孔轉染50 nmol/L siSTAT3或NC,細胞轉染參照Lipofectamine RNAiMAX說明書操作。對照組和實驗組均設3個復孔。同時轉染陽性對照siRNA,qPCR檢測基因沉默效果, 間接判斷轉染效率。
3.3qPCR檢測各基因mRNA的表達 細胞轉染48 h,提取細胞總RNA,利用隨機引物反轉錄,獲得cDNA,qPCR檢測基因mRNA表達,所得數據通過2-ΔΔCt方法計算,β-actin作為內參照,設置3個重復。
3.4Western blot檢測蛋白的表達 細胞轉染72 h,PBS洗滌,RIPA裂解細胞,收集裂解液,10 000×g離心10 min,取上清,測定蛋白濃度,分裝樣品凍存。樣品進行SDS-PAGE,轉膜,37 ℃封閉1 h,抗β-actin、STAT3和CCND2 抗體4 ℃孵育過夜,TBST洗膜3次,II抗室溫孵育1 h,Tanon 5200發光成像系統采集圖像,β-actin作為內參照。
3.5活細胞計數檢測細胞增殖 24孔板接種胰腺祖細胞,生長至40%匯合,轉染siSTAT3或NC 72 h,胰酶消化為單細胞,收集到離心管定容到1 mL,稀釋后,在光學顯微鏡下用血細胞計數板計數。對照組和實驗組均設3個復孔。
3.6CCK8法檢測細胞活力 96 孔板接種胰腺祖細胞,生長至40%匯合,轉染siSTAT3或NC 72 h,棄去培養液,PBS洗滌細胞,補加110 μL含有10% CCK8的DMEM/F12培養液,置于37 ℃培養箱中培養2 h,通過酶標儀讀取各孔吸光度(A)值。對照組和實驗組均設3個復孔。
3.7流式細胞術檢測細胞周期分布 細胞轉染72 h,收集各組細胞1×106個,PBS洗滌2次,1 100×g離心5 min,70%冷乙醇4 ℃固定2 h,PBS洗2 次,30 mg/L RNAse A 37 ℃孵育30 min,50 mg/L PI避光孵育30 min,冷PBS洗2次,600 μL PBS重懸細胞,Accuri C6流式細胞儀檢測,隨機收集30 000個細胞,采集信號,采用ModFit 4.1流式數據分析軟件進行數據分析。
4 統計學處理
用GraphPad Prism 7軟件進行統計學處理,數據采用平均數±標準誤(mean±SEM)表示,兩組間比較采用獨立樣本t檢驗,以P<0.05為差異有統計學意義。
結 果
1 敲減STAT3基因表達的效果
50 nmol/L siSTAT3或NC轉染胰腺祖細胞48 h和72 h后,分別提取RNA和蛋白,通過qPCR和Western blot檢測siRNA對STAT3基因的沉默效果。與對照組相比,實驗組胰腺祖細胞STAT3的mRNA和蛋白表達水平明顯下調(P<0.05),mRNA的表達水平降低了(49.75±0.03)%,蛋白的表達水平降低了(78.62±0.08)%,見圖1。

Figure 1. The relative expression ofSTAT3gene in the pancrea-tic progenitor cells transfected withSTAT3siRNA. A: the mRNA expression of STAT3 was detected by qPCR after transfection for 48 h; B: the protein expression of STAT3 was determined by Western blot after transfection for 72 h. Mean±SEM.n=3.**P<0.01vsNC group.
圖1 轉染STAT3siRNA后胰腺祖細胞STAT3基因表達水平的變化
2 敲減STAT3基因表達對胰腺祖細胞增殖和活力的抑制作用
細胞轉染后,倒置顯微鏡下觀察,轉染72 h后,采集細胞圖像結果顯示,轉染STAT3siRNA的胰腺祖細胞孔板中細胞密度降低,見圖2A;活細胞計數結果顯示,敲減STAT3基因的表達使細胞數量減少了(45.62±0.03)%(P<0.01),見圖2B。CCK8法檢測結果顯示敲減STAT3基因的表達使胰腺祖細胞活力減少了(43.68±0.05)%(P<0.05),見圖2C。

Figure 2. The effects ofSTAT3silencing on the proliferation and viability of pancreatic progenitor cells. A: the images of cells under the inverted optical microscope after transfection for 72 h (×100); B: the effect ofSTAT3silencing on the proliferation of pancreatic progenitor cells was detected by cell counting after transfection for 72 h; C: the effect ofSTAT3silencing on the viability of pancreatic progenitor cells was measured by CCK8 assay after transfection for 72 h. Mean±SEM.n=3.*P<0.05,**P<0.01vsNC group.
圖2 沉默STAT3對胰腺祖細胞增殖和活力的影響
3 敲減STAT3的表達對胰腺祖細胞細胞周期分布的影響
細胞轉染72 h,經流式細胞術檢測細胞周期分布。與對照組相比,轉染siSTAT3干擾片段的胰腺祖細胞其周期阻滯在G0/G1期,見圖3。
4 敲減STAT3的表達對CCND2蛋白表達的影響
細胞轉染72 h,Western blot 檢測結果可見轉染siSTAT3的胰腺祖細胞中CCND2蛋白表達水平明顯降低(P<0.05),見圖4。
討 論
糖尿病發病率呈逐年上升趨勢,利用胚胎、多能性、成體干細胞誘導生成分泌胰島素的細胞是糖尿病干細胞治療的主要研究方向,激活內源胰腺干/祖細胞生成功能細胞,將是一種更為有效和安全的治療方案,然而成體胰腺干/祖細胞數量少且常處于靜息狀態,因此,探究胰腺干/祖細胞的增殖機制具有重要的科學意義。研究表明,STAT3 在不同類型的細胞和組織中廣泛表達,參與細胞生長和凋亡等功能的調控,本研究利用RNAi技術成功敲減胰腺祖細胞STAT3基因表達,細胞計數和CCK8檢測結果表明,敲減STAT3抑制胰腺祖細胞增殖和細胞活力。
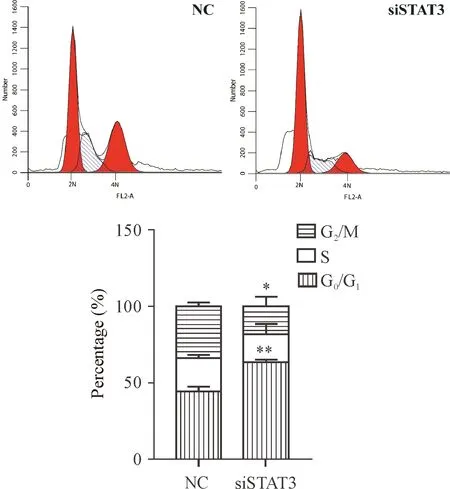
Figure 3. The effects ofSTAT3silencing on the cell cycle distribution of pancreatic progenitor cells were analyzed by flow cytometry after transfection for 72 h. Mean±SEM.n=3.*P<0.05,**P<0.01vsNC group.
圖3 流式細胞術檢測沉默STAT3對胰腺祖細胞細胞周期分布的影響
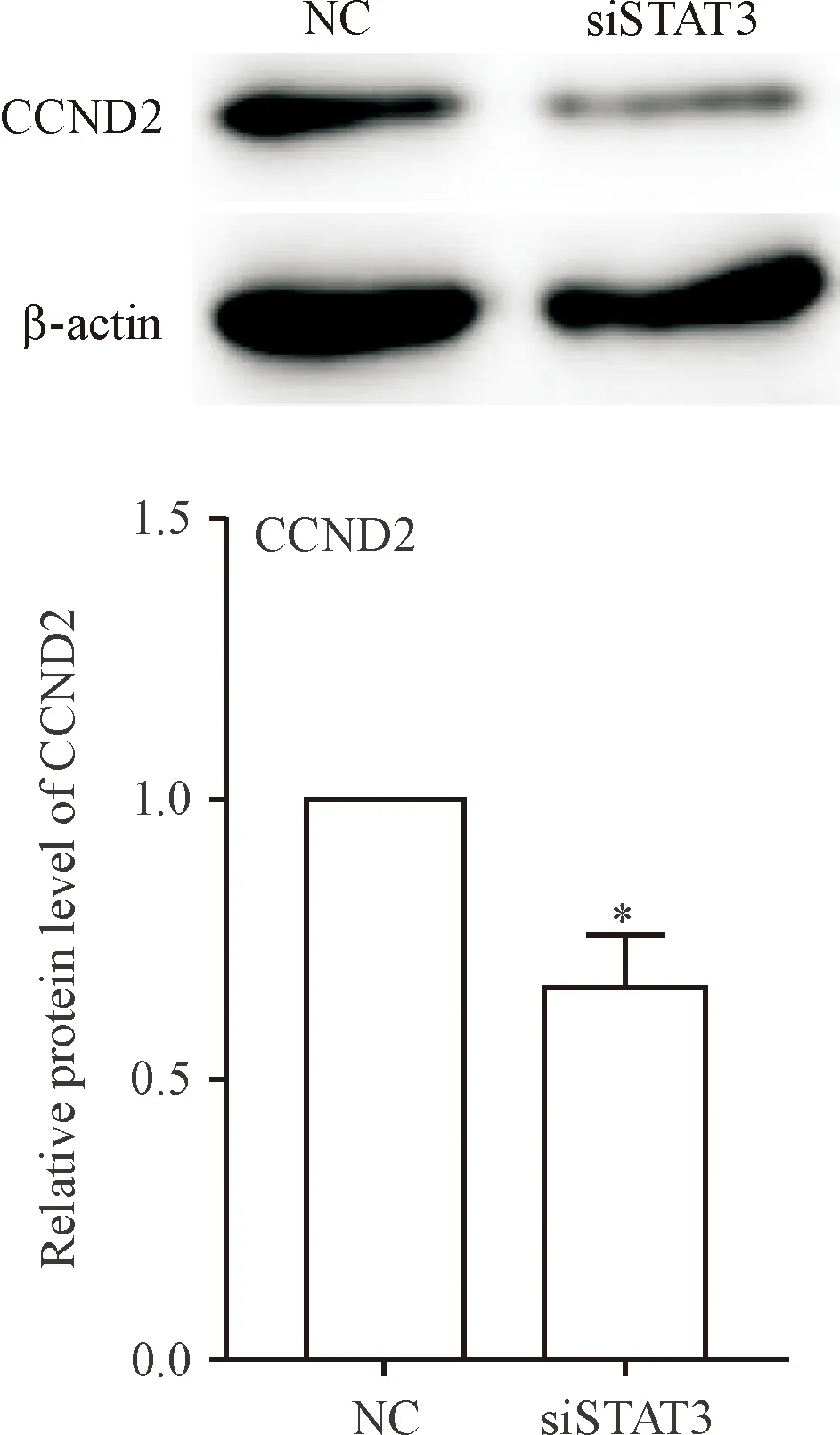
Figure 4. The effects ofSTAT3silencing on the protein expression of CCND2 in the pancreatic progenitor cells after transfection for 72 h. Mean±SEM.n=3.*P<0.05vsNC group.
圖4 沉默STAT3對胰腺祖細胞CCND2表達的影響
STAT3 是轉錄激活因子家族成員之一,在多種信號分子刺激下,STAT3被磷酸化激活,結合于靶基因DNA調控區,調控功能基因表達。STAT3活化后可通過激活其靶基因MYC原癌基因、細胞周期蛋白D1(cyclin D1,CCND1)、CCND2、存活蛋白(survivin)、基質金屬蛋白酶2(matrix metalloproteinase-2,MMP2)、p21、p27等,調控細胞增殖、細胞周期進程、遷移、侵襲和凋亡[11-12]。通過慢病毒介導的短發夾RNA(short hairpin RNA,shRNA)沉默STAT3基因表達,可降低survivin基因表達,抑制成牙骨質細胞分化,誘導細胞凋亡[13]。人工合成STAT3蛋白抗體SBT-100特異性抑制磷酸化STAT3功能,可抑制乳腺癌細胞增殖[14];siRNA沉默STAT3基因表達可降低survivin和CCND1表達,抑制白細胞介素17(interleukin-17,IL-17)對人角質形成細胞增殖的誘導作用[15]。研究表明,STAT3還通過調控下游基因表達影響干細胞增殖、自我更新和分化[16]。已報道,減弱JAK/STAT3信號活性抑制骨髓基質細胞增殖和成骨分化[17]。激活STAT3基因,促進小鼠胚胎干細胞多潛能性調控因子MYC和Bcl-3表達,有利于維持細胞自我更新特性[18-19]。STAT3促進CD24、CD34、CD38、CD44、CD90和CD133等腫瘤干細胞標志性基因表達[20],STAT3與CD44形成二聚物并乙酰化,入核后結合CCND2、MYC和TWIST1基因啟動子,激活細胞周期調控基因表達[21-22]。本研究顯示,在胰腺祖細胞中敲減STAT3基因可降低CCND2蛋白表達,使細胞周期進程阻滯在G0/G1期,抑制胰腺祖細胞細胞增殖。這表明,STAT3基因通過調控細胞周期蛋白CCND2的表達調控胰腺祖細胞增殖。
STAT3磷酸化能被多種途徑激活。EGF、血管內皮生長因子等與相應受體結合,使受體磷酸化,利用其內源性酪氨酸激酶活性直接磷酸化STAT3;IL-6等細胞因子與細胞膜上相應受體結合,募集JAK使受體的酪氨酸殘基磷酸化,募集STAT3并使其磷酸化。 研究表明,EGF是小鼠和人胚胎胰腺祖細胞、小鼠成體胰腺祖細胞維持體外增殖、自我更新和誘導分化的必需因子,用缺乏EGF的培養液培養胰腺祖細胞,使細胞增殖速度減慢[23-24]。EGF與其受體EGFR結合,使受體磷酸化,激活促進細胞增殖和細胞存活的信號通路,如JAK/STAT3、絲裂原活化蛋白激酶/細胞外信號調節激酶(mitogen-activated protein kinase/extracellular signal-regulated kinase,MAPK/ERK)和磷酯酰肌醇3-激酶/蛋白激酶B(phosphatidylinositol 3-kinase/protein kinase B,PI3K/AKT)信號通路。我們推測,敲減STAT3基因表達可能阻斷了上游EGFR/JAK/STAT3信號通路的傳導作用,從而抑制細胞增殖。
STAT3已成為轉錄因子、微小RNA(microRNA,miRNA)、長鏈非編碼RNA(long non-coding RNA,lncRNA)、小分子抑制劑和藥物調控細胞功能的靶點,如抗腫瘤藥CDK5抑制劑Roscovitine對大鼠血管平滑肌細胞增殖的抑制作用,以及lncARSR(lncRNA activated in renal cell carcinoma with sunitinib resis-tance)對肝癌干細胞增殖的促進作用是通過調控STAT3途徑實現的[25-26]。STAT3信號通路在胰腺癌、胰腺炎和糖尿病并發癥中發揮重要功能,STAT3信號通路可能成為胰腺疾病治療的靶點。本研究揭示了STAT3調控胰腺祖細胞增殖的機制,為利用胰腺干/祖細胞開展疾病治療提供了實驗依據。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
