O-N-乙酰葡萄糖胺糖基化水平正調控促血管生成素-2表達從而促進小鼠肝癌中腫瘤新生血管形成
2020-03-09 15:36:22王愛紅王明全
實驗動物與比較醫學 2020年1期
王愛紅, 王明全, 杜 娟
(1. 延安大學醫學院及延安市腫瘤防治研究重點實驗室, 延安 716000;2. 延安大學附屬醫院介入放射科, 延安 716000)
據美國癌癥學會在2018年發表的全球癌癥統計[1],肝癌在2018年將成為世界上第六大最常見的癌癥,也是全球癌癥死亡的第四大原因,每年約有841 000例新病例和782 000例死亡。因此,肝癌在全球病例數量排名第五。原發性肝癌包括肝細胞癌(HCC)(含75%~85%的病例)和肝內膽管細胞癌(包括10%~15%的病例)以及其它罕見類型。HCC的主要發病因素與慢性感染乙型肝炎病毒(HBV)或丙型肝炎病毒(HCV),重度酒精攝入、肥胖、吸煙及2型糖尿病等密切相關[2]。
在HCC患者的癌組織中O-N-乙酰葡萄糖胺(O-GlcNAc)糖基化水平表達上調,且其與腫瘤的發生發展以及肝移植后的腫瘤復發密切相關[3,4]。O- GlcNAc糖基化水平的升高與多種代謝異常相關,如糖尿病、肥胖以及腫瘤相關的代謝[5]。O-GlcNAc糖基化是O-GlcNAc單糖修飾,不存在糖鏈的延伸。O-GlcNAc糖基化也是一種動態的蛋白質修飾過程。O-GlcNAc修飾基團的添加和去除分別是由O-GlcNAc糖基轉移酶(O-GlcNAc transferase,OGT)和N-乙酰氨基葡萄糖苷酶(OGlcNAcase,OGA)來完成[6]。肝癌的形成需要充足的血液供應,新血管的形成對肝癌的進展、侵襲和轉移至關重要[7],因此,血管生成是肝癌研究和治療中的一個重要過程。血管生成素包括血管生成素1(angiopoietin-1,Ang-1)和血管生成素2(angiopoietin-2, Ang-2),Ang-1能夠促進血管的穩定生成, 而Ang-2則作為Ang-1的競爭拮抗劑發揮作用, 降低血管生成的穩定性[8,9]。有研究表明,Ang-2的表達在肝癌和肝硬化患者中升高[10],并且Ang-2高表達與預后不良顯著相關[11]。因此,Ang-2與肝癌的發生發展密切相關。在肝癌的發生發展過程中, O-GlcNAc糖基化水平升高, Ang-2表達水平也明顯上升,但二者之間是否有相互促進或抑制的關系尚不明確。因此,本課題組擬初步探討在肝癌發生發展過程中, O-GlcNAc糖基化水平升高與Ang-2表達上升是否有明確關系。
1 材料與方法
1.1 實驗動物
SPF級雄性C57BL/6小鼠,2周齡,購自第三軍醫大學大坪醫院實驗動物中心[SCXK(渝)2012-0001]。飼養于第四軍醫大學實驗動物中心SPF動物室,光照-黑暗12 h交替,自由飲水進食,室溫維持在20℃左右。
1.2 實驗細胞
人肝癌細胞HepG2,購自上海細胞庫;人臍靜脈內皮細胞(HUVEC),購自協和醫院基礎醫學研究所細胞中心。
1.3 主要試劑、抗體及儀器
二乙基亞硝胺(DEN)購自美國Sigma公司;OGA抑制劑(Thiamet, ThG)購自美國Tim Tec公司;四氧嘧啶(Alloxan) OGT抑制劑購自江蘇恒瑞醫藥公司;Western blotting O-GlcNAc抗體,購自美國Santa Cruz公司;sc-59623和免疫組織化學O-GlcNAc抗體均購自美國Pierce Biotechnology公司; Ang-2抗體(ab155106); CD31抗體(ab28364)購自美國Abcam公司; Transwell小室購自美國Coming-Constar公司。
Mini-protean Tetra System電泳儀, Trans-Blot?SD Cell 轉印系統,Imark酶標儀購自Bio-Rad公司。
1.4 實驗方法
1.4.1 動物模型建立 肝癌小鼠模型的構建及樣品采集: 小鼠適應性喂養3 d后,將小鼠隨機分為正常對照組與模型組,每組20只。于實驗第1日,模型組小鼠腹腔注射給予二乙基亞硝胺(DEN)溶液,給藥劑量為20 mg/kg,對照組小鼠給予等體積生理鹽水。實驗第5周,模型組小鼠的飲水中添加濃度為10 mg/L的DEN溶液,至實驗第9周改為正常飲水。對照組小鼠在實驗過程中正常飲水[12]。每2 d監測一次體質量和觀察精神狀態。于實驗11個月時處死各組小鼠,質量分數4%多聚甲醛溶液固定部分肝臟組織或肝癌組織用于病理觀察,剩余肝臟組織保存于-80℃備用。
1.4.2 免疫組織化學分析肝癌模型小鼠中Ang-2、O-GlcNAc糖基化及CD31的表達水平 將組織切片置于室溫30 min后, 4 ℃丙酮溶液中固定10 min,PBS洗滌2次(每次洗滌5 min)。然后,內源性過氧化物過氧化氫(H2O2) 孵育10 min,37 ℃下正常山羊血清封閉20 min,5% BSA稀釋抗體(Ang-2,1∶500;O-GlcNAc,1∶200,CD31,1∶50),4 ℃過夜。第2日,PBS沖洗3次,每次5 min。滴加生物素標記二抗工作液及辣根酶標記鏈酶卵白素工作液,各37 ℃孵育30 min。最后,滴加DAB顯色液顯色。自來水沖洗,蘇木素復染,封片,用光學顯微鏡拍片。使用Image Pro Plus軟件進行分析。黃色的區域反映靶蛋白的陽性表達。以積分吸光度SUM/Area的比值對Ang-2,O-GlcNAc糖基化的表達進行定量。通過CD31陽性染色確定的血管數量,并在8個視野中計數取微血管密度平均值。
1.4.3 細胞培養和轉染 人HUVEC細胞及HepG2細胞用含10%胎牛血清的DMEM培養, 并放置于37 ℃, 5% CO2培養箱中, 并使用5 mmol/L的Alloxan與ThG分別干預HUVEC細胞及HepG2細胞。以不含Alloxan藥物或ThG培養基常規培養的HUVEC細胞及HepG2細胞作為各自的對照組。
1.4.4 qRT-PCR和Western blotting檢測O-GlcNAc糖基化及Ang-2的mRNA和蛋白水平 qRT-PCR:取手術切除的肝癌組織,采用TRIzol法提取組織總RNA,在37℃,30 min,85℃,5s的條件下進行反轉錄,獲得cDNA,采用 SYBR green實時定量PCR法進行擴增和檢測。
Western blotting:將收集的細胞采用全蛋白提取試劑盒提取蛋白,BCA蛋白含量檢測試劑盒測蛋白濃度。用10% SDS-PAGE凝膠分離蛋白樣品,轉移到PVDF膜上,一抗孵育4 ℃過夜(一抗濃度,O-GlcNAc: 1∶500, Ang-2,1∶2000)。二抗在室溫下孵育1.5 h(濃度均為1∶2 000),ECL顯影后,Bio-RAD儀器掃描,Image Lab進行條帶分析,目的蛋白灰度值/GAPDH灰度值為目的蛋白的相對表達量。
1.4.5 MTT法檢測O-GlcNAc糖基化水平對HepG2細胞增殖的影響 細胞長滿至80%時消化并計數,將每種細胞分別接種200 μL混懸液約3 000個于96孔板,每組細胞做5個復孔,共2塊板,分別培養24h,48h。待細胞貼壁后,把配好的OGT抑制劑及OGA抑制劑分別加入到不同的培養孔中,開始對細胞培養時間計時,等到各個時間點依次收板,避光條件下加入MTT試劑20 μL于每孔中,此時設置調零孔,繼續放入培養箱中孵育4 h后避光取出,去培養基和MTT試劑混合液,加入DMSO 150 μL于每孔中,置于室溫搖床上混勻10 min,用酶標儀490 nm檢測吸光度(A)值。
1.4.6 Transwell檢測O-GlcNAc糖基化水平對HUVEC細胞遷移侵襲能力的影響 HUVEC細胞消化后,置于0.1% BSA的培養基中,并計數,取1×109/L細胞100 μL加入Transwell小室中的上室,并分別加入5 mmol/L的OGT抑制劑及OGA抑制劑,下室中加入400 μL 0.1%小牛血清培養基,37 ℃,5% CO2條件下培養24 h后,去除上層細胞,下層細胞用95%乙醇溶液固定,并用臺盼藍染色。于鏡下計數,以遷移細胞數目表示細胞的遷移能力。
1.5 統計方法
2 結果
2.1 模型小鼠大體表現
在造模過程中,觀察到模型組小鼠飲食量減少、行動緩慢、精神萎靡、且體質量明顯下降。造模11個月時脫頸處死小鼠, 取肝臟拍照(圖1)并稱重。對照組小鼠肝臟表面光滑, 色澤暗紅且有光澤,而使用DEN干預的模型組小鼠的肝臟體積明顯增大,表面粗糙,顯示出許多大小不同的腫瘤, 顏色暗淡成淡黃色, 且在大部分肝臟區域可見。模型組小鼠肝臟重量(2.78±1.25 g)約為正常組小鼠(0.92±0.19 g)的3倍(P<0.05)。模型組最終發生肝癌的小鼠有17只,肝癌發生率為85%。
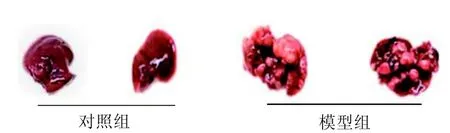
圖 1 各組小鼠肝組織形態
2.2 小鼠肝臟病理形態變化及Ang-2,O-GlcNAc糖基化蛋白表達水平
HE染色結果顯示,對照組肝細胞排列整齊且有規則,肝細胞大小均勻。而模型組小鼠肝細胞變大,大小不一,有炎細胞浸潤,細胞核變大,肝細胞形態結構不清晰且排列紊亂(圖2)。免疫組織化學結果顯示,與對照組中Ang-2蛋白表達水平(0.15±0.05)和O-GlcNAc蛋白表達水平(0.13±0.05)相比,模型組中Ang-2在細胞質中高表達水平為0.36±0.17, O-GlcNAc在細胞核中高表達水平為0.28±0.14, 組間差異具有統計學意義(P<0.01), 即肝癌模型組中, Ang-2與O-GlcNAc的表達均是上調的。與對照組CD31陽性染色的微血管密度(43±16)相比,模型組CD31陽性染色的微血管密度為107±23,其差異具有統計學意義(P<0.01),表明模型組小鼠肝臟組織中血管數量明顯上升。
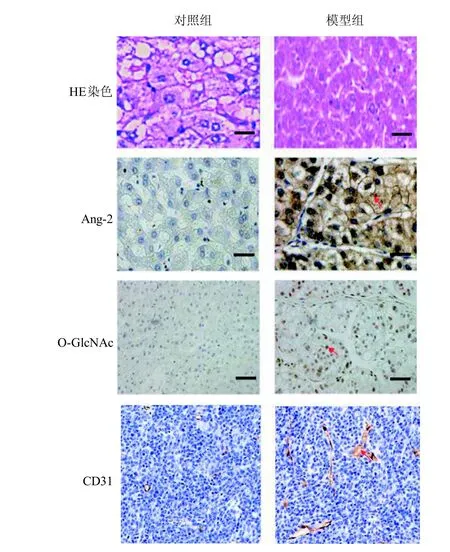
圖 2 小鼠肝癌模型病理變化及免疫組織化學結果 (×400)
2.3 O-GlcNAc糖基化及Ang-2 mRNA表達水平
結果表明,Ang-2表達水平受到O-GlcNAc糖基化表達水平的影響(表1)。
2.4 O-GlcNAc糖基化和Ang-2的蛋白表達水平
如圖3所示,使用OGT抑制劑感染HepG2細胞抑制O-GlcNAc的表達后,Ang-2的表達也隨之降低;而使用OGA抑制劑感染HepG2細胞使O-GlcNAc高表達后,Ang-2的表達水平也明顯上升。以上結果顯示,在肝癌細胞中,OGlcNAc可能是Ang-2的上游蛋白,其表達的變化能引起Ang-2的變化。
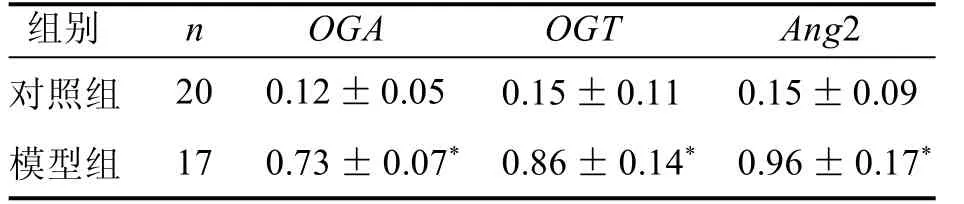
表 1 兩組小鼠OGA、OGT、Ang-2 mRNA相對表達量
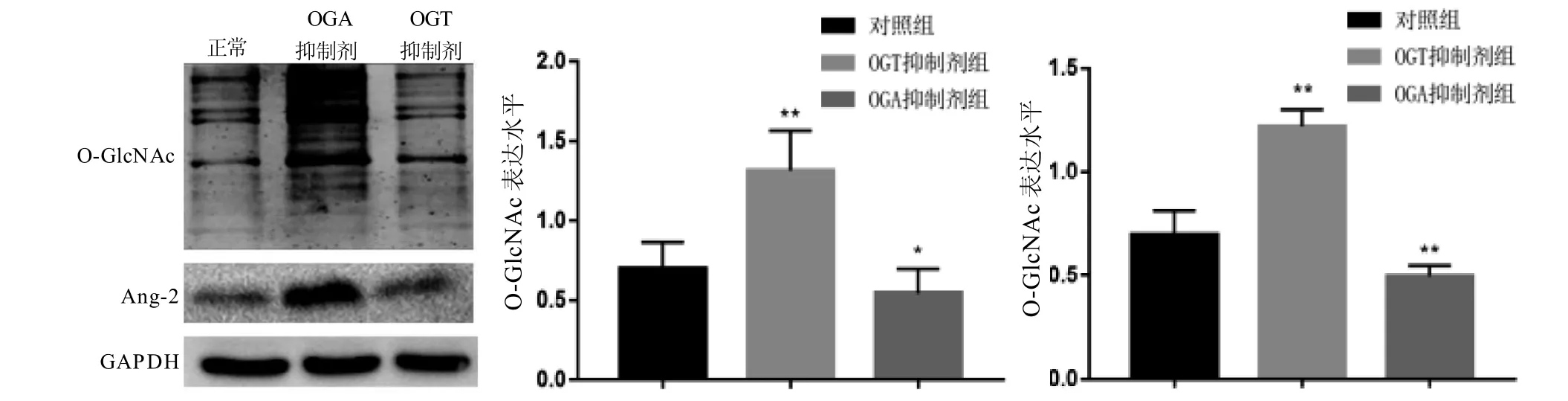
圖 3 HepG2細胞分別添加OGT抑制劑及OGA抑制劑后O-GlcNAc及Ang-2蛋白表達
2.5 O-GlcNAc糖基化水平能促進肝癌細胞增殖
在添加OGT抑制劑24 h后,與對照組相比,細胞的增殖能力明顯受到抑制(P<0.05)。而添加OGA抑制劑24 h后,肝癌細胞增殖能力有所增加(P<0.05)。48 h時, 添加了OGT抑制劑組較24 h對肝癌細胞增殖抑制效果明顯增強(P<0.01),而OGA抑制劑組能夠明顯提高細胞的增殖能力(P<0.01)(圖4)。這些結果說明,O-GlcNAc糖基化水平增高可以促進肝癌細胞的增殖能力。
2.6 O-GlcNAc糖基化水平對HUVEC細胞和HepG2細胞遷移侵襲能力的影響
Transwell結果顯示(圖5~8),經5 mmol/L的Alloxan干預HUVEC細胞和HepG2細胞24 h后,干預組穿膜細胞數與對照組相比明顯減少(P<0.01); 而經5 mmol/L的ThG干預HUVEC細胞和HepG2細胞24 h后,干預組穿膜細胞數較對照組顯著增加(P<0.01)。結果說明, HUVEC細胞及HepG2細胞遷移侵襲能力明顯受到O-GlcNAc糖基化水平影響,且與O-GlcNAc糖基化水平呈正相關。
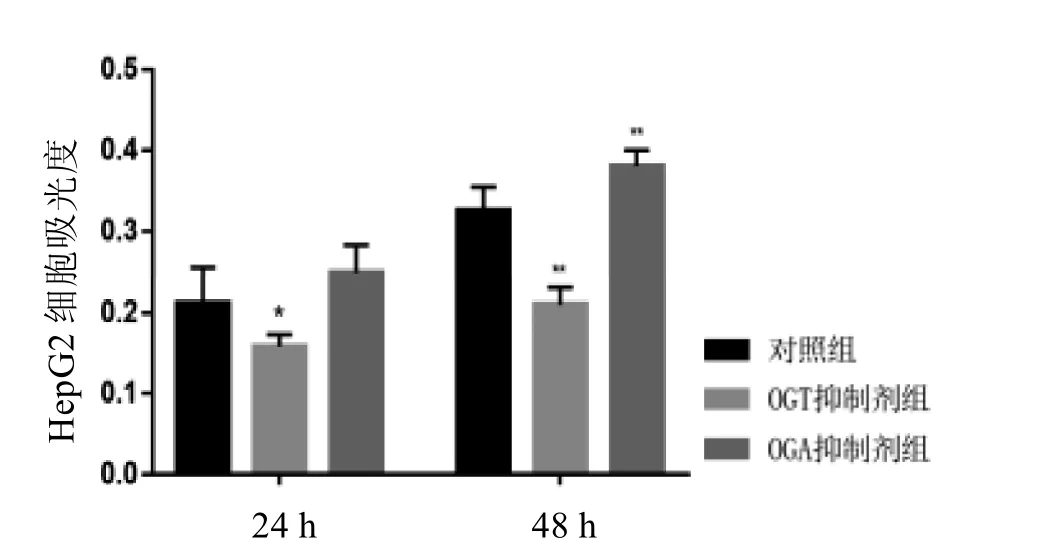
圖 4 O-GlcNAc表達的變化對肝癌細胞HepG2增殖影響
3 討論
圖 5 三組HUVEC細胞遷移侵襲能力比較
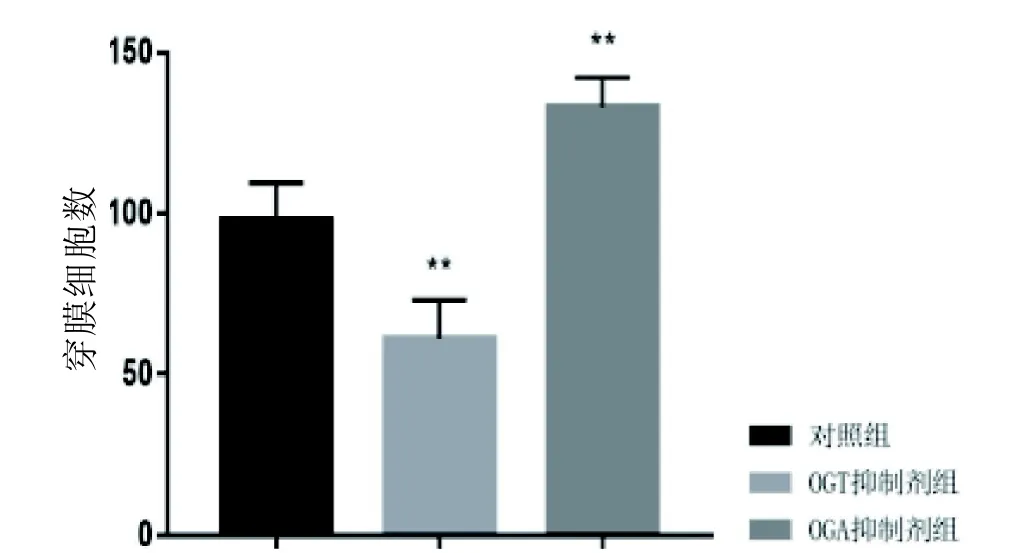
圖 6 O-GlcNAc糖基化水平對HUVEC細胞遷移侵襲能力的影響
HCC是多中心、多階段、多病因以及連續的發展過程[13]。在化學因素誘導肝癌動物模型的實驗中,其中以DEN誘發肝癌模型為最常見,DEN是一種化學致癌物質,由細胞色素P450生物活化(CYP)酶存在于肝臟中,導致DNA化合物通過烷基化機制形成。DEN誘發的肝癌體現了類似于人類HCC的基因表達譜[14],其最經典的方法是“飲水法”建立肝癌動物模型。成功建模后的腫瘤特點兼具肝炎、肝硬化的背景,與人類肝癌形成過程十分類似,為肝癌的研究提供了很好的借鑒方法[15]。本文以DEN誘導的小鼠肝癌模型和人肝癌細胞HepG2作為研究對象,研究肝癌中O-GlcNAc糖基化水平與Ang-2的關系。
圖 7 三組HepG2細胞遷移侵襲能力比較
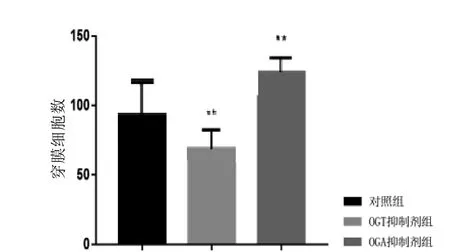
圖 8 O-GlcNAc糖基化水平對HepG2細胞遷移侵襲能力的影響
在HCC患者肝癌組織中,蛋白質O-GlcNAc糖基化可促進肝癌細胞的增殖和轉移。肝癌患者肝臟移植后, 肝癌復發的組織中蛋白質O-GlcNAc糖基化水平顯著高于肝臟移植后肝癌未復發的組織,而肝癌復發的組織中OGA蛋白表達水平顯著低于肝癌未復發的組織,OGT的表達水平在肝癌復發和未復發患者之間沒有差異。Ang-2參與肝癌發生發展過程,其通過破壞血管的穩定性,增強血管內皮生長因子(vascular endothelial growth factor,VEGF)的表達,促使新生血管的生成。Ang-2與腫瘤血管形成的數目以及腫瘤的預后關系密切[16]。有研究[17]表明,Ang-2的表達不僅在肝癌組織上調,且在肝癌患者血清中其濃度也明顯上升,以Ang-2作為靶點的藥物治療可明顯抑制人肝癌細胞的生長。
在本研究中,我們在DEN誘導的動物模型中觀察到,O-GlcNAc糖基化水平與Ang-2在肝癌腫瘤組織中表達均明顯上升,且微血管密度也顯著上升。通過OGA、OGT、Ang2 mRNA的檢測,結果顯示,O-GlcNAc糖基化水平變化影響Ang2 mRNA的表達水平;而且O-GlcNAc糖基化和Ang-2的蛋白表達水平之間也呈現相關關系,當使用OGT抑制劑后,O-GlcNAc糖基化水平明顯下降,HepG2細胞的增殖受到抑制,Ang-2的表達水平也隨之下降;使用OGA抑制劑后,Ang-2的表達水平隨著O-GlcNAc糖基化水平上升也明顯升高,且HepG2細胞的增殖能力明顯增強;最后,我們使用Transwell法檢測發現,HUVEC細胞的遷移侵襲能力明顯受到O-GlcNAc糖基化水平的調控。這些結果說明,在肝癌的發生發展過程中,O-GlcNAc糖基化水平逐漸上升,且Ang-2的表達水平可能受到O-GlcNAc糖基化的正調控,從而促進腫瘤新生血管的生成。本研究提示, 在肝癌進展中, Ang-2受到O-GlcNAc糖基化正調控, 可為未來肝癌的預防及治療提供新的思路。但是,本研究中O-GlcNAc糖基化調控Ang-2的具體機制仍未清楚, 有待繼續深入探討。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
人大建設(2019年12期)2019-05-21 02:55:32
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
癌變·畸變·突變(2015年4期)2015-02-27 06:15:25
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
