新疆雙峰駝納米抗體、重鏈抗體和常規抗體的熱穩定性比較
2020-03-12 01:40:04坤杜孜阿依阿布都沙拉木姜志強劉婭菲熊靜璠娜斯拜阿卜杜瓦哈普李江偉
新疆農業科學 2020年2期
坤杜孜阿依·阿布都沙拉木,姜志強,劉婭菲,熊靜璠,娜斯拜·阿卜杜瓦哈普,李江偉
(新疆大學生命科學與技術學院/省部共建新疆生物資源基因工程重點實驗室,烏魯木齊 830046)
0 引 言
【研究意義】1993年發現在駱駝科動物中存在天然缺失輕鏈的重鏈抗體(Heavy Chain Antibodies, HCAbs)以來[1],但研究者對于HCAbs自身的功能和特性、其在駱駝體液免疫系統中的作用、與疾病的關系等研究較欠缺、鮮有文獻報道。研究駱駝HCAbs的功能及性質,對分析駱駝免疫系統的功能,研究更有效的疾病預防和治療手段,開發和利用這類抗體具有重要意義。【前人研究進展】 駱駝科現存3屬6種動物。在所有這些動物中,除具有由2條重鏈和2條輕鏈組成的IgG1型常規抗體外,還存在天然缺失輕鏈和CH1結構域的IgG2和IgG3型重鏈抗體。在自然界中,缺失輕鏈的HCAbs目前僅見于駱駝科動物和幾種軟骨魚類。當前,大量針對重鏈抗體的研究主要集中在納米抗體的功能特性及應用方面。納米抗體除了具有完全的抗原結合活性外,還具有常規抗體所不具備的諸多優勢[2],包括如組織穿透能力強、穩定性極高、免疫原性低、易與蛋白裂隙中的隱蔽表位結合[3]、對酶活性具有抑制作用[4-6]等。【本研究切入點】盡管納米抗體的研究和應用受到了極大的重視,但對于HCAbs自身的功能和特性、其在駱駝體液免疫系統中的作用、與疾病的關系等研究、鮮有文獻報道。當前對駱駝HCAb的物理化學性質的研究,主要集中在對其可變區即納米抗體的溫度穩定性方面,發現納米抗體普遍具有比常規抗體更高的溫度耐受性[2-6],然而其在體內的天然形式,即重鏈抗體是否也具有與納米抗體一樣的溫度穩定性,重鏈抗體是否比常規抗體溫度穩定性更高,尚缺乏研究。研究比較納米抗體、HCAbs與常規抗體之間在溫度穩定性方面的差異。【擬解決的關鍵問題】 研究HCAbs與常規抗體及納米抗體之間在穩定性的特點,對采用3種不同抗原免疫的新疆雙峰駝,以及免疫血清中的IgG1常規抗體與IgG2和IgG3重鏈抗體及其對應的納米抗體,在不同溫度處理下的抗原結合活性進行比較,分析其溫度穩定性的差異,為研究HCAb的功能特性和駱駝適應極端環境的免疫特點提供參考。
1 材料與方法
1.1 材 料
實驗動物:3只健康新疆雙峰駝(Camelus bactrianus),年齡均在3.5年,雌性2只,雄性1只,由新疆生產建設兵團第十二師提供。
菌種和質粒:大腸桿菌 DH5α、BL21(DE3)和重組質粒 pET30a-spaA-N,pET30a/Lys-23,pET30a/Spa-G26和pET30a/Ali-A4由本室保存。Lys-23、Ali-A4 和Spa-G26為研究組通過噬菌體展示技術從VHH駱駝免疫文庫中篩選獲得的分別結合溶菌酶、蒜氨酸酶和丹毒絲菌表面抗原A(spaA)的納米抗體[7-8]。
主要試劑:溶菌酶(12650-88-3)購自 Sigma-Aldrich公司,Protein A Resin FF(L00464)和Protein G Resin FF(L00664)購自南京金斯瑞公司, HRP conjugated Goat anti-Llama IgG(H&L)購自ImmunoReagents公司,Ni-sephorase 6 FF(17-5318-01)購自GE公司。
1.2 方 法
1.2.1 駱駝免疫
溶菌酶、蒜氨酸酶和spaA 3種抗原分別與弗氏完全佐劑(初免)和不完全佐劑(加強)等體積充分混勻乳化作為免疫原,分別對雙峰駝進行頸部皮下注射,免疫劑量為 600 μg/次,每2周加強1次,共進行5次免疫。每次免疫前和終免7 d后,頸靜脈采血,分離血清,-20℃保存備用。
1.2.2 血清IgG抗體不同亞型的分離和純化
按Hamers-Casterman[1]方法,分離駱駝免疫血清中的IgG1、IgG2和IgG3抗體亞型。簡述如下:將駱駝免疫血清2 mL以PBS稀釋2倍后,加入到 Protein G Resin層析柱中,結合10 min,收集與Protein G未結合的流出液備用,分別以洗脫緩沖液 A(0.15 mol/L NaCl、0.58%乙酸、pH 3.5)和洗脫緩沖液 B(0.1 mol/L 甘氨酸,pH 2.7)先后洗脫獲得 IgG1 及 IgG3組分。將收集的 Protein G 未結合的流出液,加入到 Protein A Resin層析柱中,結合10 min,以洗脫緩沖液 C(0.1 mol/L 甘氨酸,pH 2.0)洗脫獲得 IgG2組分。洗脫的各組分立即中和至 pH 值7.4,在還原條件下對分離的各個亞型 IgG 抗體進行SDS-PAGE 檢測。
1.2.3 納米抗體的表達與純化
提取納米抗體重組質粒,鑒定正確后轉化至BL21(DE3)感受態細菌。選取陽性轉化子誘導表達蛋白。在0.2 mmol/mL IPTG,16℃,150 r/min的條件下過夜誘導表達。誘導后的菌液進行離心處理,超聲后再12 000 r/min離心取上清,使用Ni柱親和層析純化重組納米抗體,15%SDS-PAGE檢測納米抗體蛋白的表達及純化。
1.2.4 ELISA測定免疫血清不同亞型抗體效價
免疫血清、分離純化的IgG各亞型抗體以及納米抗體與抗原的結合采用間接ELISA測定。檢測設立3個平行重復,終數據取平均值。在96孔板上分別包被1 μg/mL溶菌酶、蒜氨酸酶和spaA 3種抗原。BSA封閉后,加入系列稀釋的免疫血清和納米抗體,孵育后加入羊抗美洲駝IgG(H&L)-HRP抗體,稀釋比例為1∶3 000。加入顯色底物TMB,在酶標儀中測定450 nm 吸光值。
1.2.5 免疫血清不同亞型抗體與納米抗體溫度穩定性測定
將純化的各亞型IgG及納米抗體分別在25、37、50、60、70、80和90℃水浴孵育0.5 h,平衡到室溫,在96孔板上包被1 μg/mL各抗原,加入稀釋至5 μg/mL的各亞型IgG及納米抗體,通過ELISA測定其剩余結合活性大小。二抗分別為羊抗美洲駝IgG(H&L)-HRP抗體和鼠抗His-HRP,加入顯色底物TMB,在酶標儀中測定450 nm 吸光值。
采用ELISA測定各抗體在不同溫度處理下剩余抗原結合活性,每組設立3個平行重復孔,以室溫(25℃)處理后的結合值作為100%結合對照,計算每組重復測定樣品的平均吸光度值,計算各抗體的相對剩余結合活性:RT Binding =(OD450heat treated/ OD45025℃treated)×100%。
2 結果與分析
2.1 新疆雙峰駝免疫血清效價的測定
采用ELISA測定溶菌酶、蒜氨酸酶和spaA 3種抗原第5次免疫駱駝的血清抗體效價。在最后1次(5次)免疫后,3種抗原均能激發明顯的抗體反應,抗體效價均在1∶10 000以上。圖1
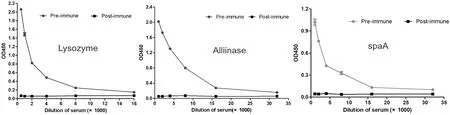
圖1 溶菌酶、蒜氨酸酶和spaA第5次免疫雙峰駝抗體效價
Fig.1 Camel serum titers of immunization with lysome,alliinase and spaA
2.2 血清IgG抗體不同亞型的分離和純化
基于駱駝IgG1、IgG2和IgG3與蛋白G和蛋白A的不同結合條件,通過蛋白G+A進行分離。還原條件下 SDS-PAGE顯示經蛋白G+A純化后,先后洗脫獲得3種不同分子量的 IgG 抗體,分別為43 KD(重鏈)的IgG3,50 KD(重鏈)和25KD(輕鏈)的IgG1和46 KD(重鏈)的IgG2,與Hamers-Casterman[1]報道的大小基本相符。圖2

圖2 還原條件下 SDS-PAGE 分析 protein G 和protein A 純化的雙峰駝免疫血清中 各IgG亞型
Fig.2 SDS-PAGE analysis of different IgG isotype fractions purified by protein G and protein A chromatography from immune cserum of.bactrianus under reduced condition
2.3 納米抗體的表達與純化
保存的pET30a/Lys-23,pET30a/Spa-G26和pET30a/Ali-A4三種質粒轉化BL21(DE3)感受態細菌后,挑取陽性轉化菌,在0.2 mmol/mL IPTG誘導下,三種表達菌均可以表達重組納米抗體Lys-23,Spa-G26和Ali-A4。大量誘導表達后,在Ni柱親和層析中均獲得了較高純度的納米抗體,3種納米抗體表觀分子量在28Kd(Lys-23和Spa-G26)和23Kd(Ali-A4)附近,與預期大小相符。圖3

圖3 SDS-PAGE 分析采用Ni離子親和色譜純化BL21菌表達的3種納米抗體Lys-23,Spa-G26和Ali-A4
Fig.3 SDS-PAGE analysis of purified nanobodies of Lys-23 ,Spa-G26 and Ali-A4 by Ni ion affinity chromatography fromE.coliBL21(DE3)
2.4 免疫血清不同亞型抗體和對應納米抗體溫度穩定性的比較
研究表明,3種納米抗體都表現出較高的溫度穩定性,其中納米抗體Lys-23 和Spa-G26即使在90℃處理下,仍然保留90%的結合活性。而納米抗體Ali-A4穩定性稍差,90℃處理下,可以保留50%的結合活性。對于3種抗原特異的IgG1常規抗體來說,對溫度的敏感性表現出一致性。在60℃溫度處理下,抗原結合活性只有50%或更低,而在更高溫度下處理,抗原結合活性下降很快。在80℃以上,只有10%的結合活性。對于IgG2和IgG3重鏈抗體來說,在60℃以上的溫度處理下,其抗原結合活性比IgG1常規抗體明顯要高,其中,IgG3似乎比IgG2對溫度更穩定。但不同的抗原特異的重鏈抗體,其溫度敏感性具有較多差異。在實驗中,溶菌酶特異的IgG2和IgG3重鏈抗體具有更高的穩定性,而蒜氨酸酶結合的IgG2和IgG3重鏈抗體的穩定性較低。另外,在測定的3個納米抗體溫度穩定性中,2個納米抗體Lys-23 和Spa-G26在80℃處理下,表現出比室溫更高的結合活性,其相對剩余結合活性分別為105%和110%.顯示出這2個納米抗體極高的溫度耐受性。圖4

圖4 不同亞型抗體和對應納米抗體溫度穩定性的比較
Fig.4 Comparison of thermal stability among nanobodies,conventional antibodies and HCAbs
3 討 論
通過親和色譜方法分離和純化得到雙峰駝免疫血清中的IgG1、IgG2和IgG3 3種亞型抗體,采用這些抗體和原核表達純化得到的相應納米抗體,首次全面比較了同一抗原結合的重鏈抗體、常規抗體和納米抗體在溫度穩定性方面的差異。在3種抗體類型中,納米抗體具有最高的溫度穩定性,重鏈抗體與常規抗體相比具有一定的溫度穩定性,而由重鏈和輕鏈組成的常規抗體對較高溫度的耐受性較差。
經典的IgG抗體由2條相同的重鏈(H)和2條相同的輕鏈(L)組成,由H鏈和L鏈的可變區(V)共同構成抗體的抗原結合位區。在B細胞的成熟過程中,通過克隆選擇和VDJ重排以及體細胞突變和親和力成熟等機制,形成了高度多樣性的抗體庫(repertoire)。盡管這個過程在不同脊椎動物有所不同,但最后都組裝形成H2L2結構的雙價分子,顯示出IgG結構在進化中的高度穩定性。HCAbs大約在2 500萬年前才在駱駝科出現,屬于進化上較新的事件[9]。與常規抗體由12個免疫球蛋白折疊域(immunoglobulin fold domains)組成結構不同,HCAbs只包含6個免疫球蛋白折疊域,而納米抗體僅有1個免疫球蛋白折疊域。因此,在應對溫度等變性條件時,理論上具有較少折疊形式的蛋白片段具有更高的耐受性。本研究取得的試驗結果支持該假說。圖5

圖5 常規抗體和重鏈抗體結構域比較
Fig.5 Comparison of immunoglobulin fold domainsbetween conventional and heavy-chain antibodies
以往對駱駝抗體穩定方面的研究主要集中在納米抗體上,很少開展針對HCAbs穩定方面的研究。抗體穩定性除了與免疫球蛋白折疊域數量相關外,還受Fc結構、鉸鏈長度、鏈間和鏈內二硫鍵、可變區CDR序列的影響[10-18].通常情況下,抗體在高于65 ℃溫度下就會喪失變性后的可逆性復性作用[12-13],檢測的駱駝IgG1符合多數抗體的復性特征。而對于駱駝HCAbs,在檢測的溶菌酶和spaA2種抗原特異的IgG2和IgG3重鏈抗體中,其在65℃溫度下的耐受性表現與常規抗體不一樣,而與其對應的納米抗體一致。另外,在試實驗中,IgG3具有比IgG2更高的溫度穩定性趨勢,這可能與2種抗體鉸鏈長短不同有關。IgG3屬于短鉸鏈抗體,只有12個氨基酸,而IgG2擁有由36個氨基酸組成的長鉸鏈。因此,從鉸鏈長度來看,IgG3比IgG2在變性條件下更容易復性。納米抗體溫度穩定性的機制主要有二相穩態和疏水內核等學說[19-22]。對于納米抗體來說,其小分子和單域屬性是其耐受溫度的主要機制[23],然而并不是所有的納米抗體都具有很好的溫度穩定性。在以往研究的多個納米抗體中,大約有1/3在高于65 ℃處理下,不具有可逆的復性活性,最近Kunz P等[15]通過對近70個納米抗體的溫度穩定性研究中也發現有相當數量的納米抗體在65 ℃處理下發生了不可逆的聚集。2個納米抗體Lys-23 和Spa-G26即使在90℃處理下,仍然保留90%的結合活性,具有極高的溫度穩定性。而納米抗體Ali-A4穩定性稍差,在65℃下,大約可以保留70%的結合活性。這些差異可能與這些納米抗體中的二硫鍵有關。Govaert J[18]對二硫鍵在納米抗體中的作用進行的研究表明,鏈內二硫鍵具有穩定納米抗體結構并有助于CDR3發揮剛性結構作用。Lys-23 和Spa-G26都具有一對鏈內二硫鍵,而Ali-A4不具有鏈內二硫鍵。其穩定性符合二硫鍵在納米抗體中的作用。
4 結 論
駱駝抗體中的重鏈抗體、常規抗體和納米抗體在對高溫條件的變性作用下,其可逆性復性作用不盡相同,其中納米抗體由于小分子和單域屬性對溫度處理最耐受,重鏈抗體IgG2和IgG3由于減少的免疫球蛋白折疊域數量,也表現出一定的對高溫變性條件的耐受性,而代表常規抗體的IgG1型抗體表現出與其他哺乳動物相似的對高溫變性條件的敏感性。了解這些抗體的穩定性特征,可以為理解HCAb的功能特性和駱駝適應極端環境的免疫特點提供線索。另外,在納米抗體的應用中,穩定性和可溶性都將影響到納米抗體的工業化生產和在體內的效應。為了延長代謝時間,納米抗體通常以Fc片段連接的形式在體內作用。
