溫度與NaCl處理對甜高粱種子萌發及幼苗抗氧化酶活性的影響
2020-03-13 08:07:28王志恒黃思麒鄒芳楊秀柳魏玉清
中國農業科技導報 2020年9期
關鍵詞:影響
王志恒, 黃思麒, 鄒芳, 楊秀柳, 魏玉清
(北方民族大學生物科學與工程學院, 國家民委生態系統模型及應用重點實驗室, 銀川 750021)
甜高粱[Sorghumbicolor(L.)Moench]又稱糖高粱、甜稈等,具有適應性廣、生長速度快、生物產量高等特點,可廣泛作為飼料、能源和糖料作物,現已成為鹽堿地地區最具有種植潛力的作物之一[1-2]。當前,土壤鹽漬化是限制農業發展的重要環境因子,嚴重威脅作物產量和農村經濟發展[3-4]。據全國第二次土壤普查,中國的鹽漬地分布廣泛,總面積約為3.6×109hm2,其中西北、華北、沿海地區的鹽堿地總面積超過3×108hm2,占全國耕地總面積的10%左右,發展耐鹽植物在鹽堿地的種植成為鹽堿地可持續利用技術研發研究重點之一,對于提升我國耕地農業生產能力、開發耕地面積、保障國家糧食安全具有重要意義[5-6]。
種子萌發階段作為植物生長周期中最重要的階段之一,是自身最脆弱、對外界環境最敏感的時期,極易受到環境因子的影響,而溫度和鹽分均是影響種子萌發的主要環境因素[7]。有研究表明,生長在鹽堿地區的植物經常遇到溫度和鹽分兩種脅迫[8]。目前,關于鹽脅迫對甜高粱種子萌發的影響研究較多,而不同溫度條件下鹽脅迫對甜高粱種子萌發影響的研究未見報道。那么溫度和鹽分及其交互作用對甜高粱種子的萌發有何影響,值得研究。本研究以‘遼甜1號’甜高粱為材料,設置5個溫度梯度下5個NaCl鹽分濃度,通過測定發芽率、種子殘留干重、幼芽幼根長度及幼苗抗氧化物酶活性等多個指標,擬探討不同溫度及NaCl濃度脅迫下甜高粱種子萌發和抗氧化酶活性的變化特征,旨在研究溫度和鹽分及其交互作用對甜高粱種子萌發及幼苗生長的影響,以期為鹽堿地甜高粱的種植提供理論依據和技術指導,從而促進我國鹽堿地甜高粱的產業化發展。
1 材料與方法
1.1 供試材料
供試種子為甜高粱品種‘遼甜1號’,由遼寧省農業科學院創新中心提供。
1.2 試驗設置及方法
萌發溫度設置為15/5、20/10、25/15、30/20與35/25 ℃共5個梯度,光周期為12/12 h(光/暗);將NaCl配制為0、50、100、200、300 mmol·L-1鹽溶液進行梯度處理,共設置25個處理。
取飽滿、色澤大小基本一致的甜高粱種子適量,用10%的次氯酸鈉溶液浸泡20 min消毒,用無菌水沖洗7~8次,直至干凈無味,用濾紙吸干種子表面水分。將甜高粱種子30粒整齊置于墊有雙層濾紙的的培養皿(Φ120 mm)中,每個處理加入10 mL不同濃度的鹽溶液,均設置3個重復。試驗期間每天更換培養基質,以保證培養環境的恒定。置于特定溫度的室內人工智能氣候箱(BIC-800,上海博迅實業有限公司)黑暗條件下進行甜高粱種子萌發試驗。
1.3 指標檢測
試驗過程中,每隔24 h觀察記錄種子萌發情況,以胚根伸出種皮2 mm為萌發標準[8],記錄種子萌發個數,統計萌發率。7 d后結束萌發,從各個處理中隨機選出15株幼苗,用直尺測量胚芽、胚根長度,用電子分析天平(上海精科天美科學儀器公司)測量幼苗干重、種子殘留干重。干重測量前105 ℃烘2 h殺青,80 ℃烘24 h至恒重。按以下公式計算各指標[9-10]。


發芽指數(GI)=∑Gt/t
式中,Gt為t天內發芽的種子數,t為相應的萌發天數。
活力指數=GI×S
式中GI為發芽指數,S為胚根平均長度。
干物質轉運率=
超氧化物歧化酶(superoxide dismutase, SOD)活性測定采用氮藍四唑光還原法[11];過氧化物酶(peroxidase, POD)活性測定采用愈創木酚顯色法[12];過氧化氫酶(catalase, CAT)活性測定采用紫外吸收比色法[13];抗壞血酸過氧化物酶(ascorbate peroxidase, APX)活性測定參考孫云[14]的方法。
1.4 數據處理
采用 Microsoft Excel 2003進行數據統計處理,SPSS Satatistics 21.0進行雙因素方差分析,GraphPad Prism 5.0進行作圖。
2 結果與分析
2.1 不同處理對甜高粱種子發芽進程的影響
不同溫度不同NaCl處理的甜高粱種子發芽結果見圖1,可知溫度和NaCl處理均影響甜高粱種子發芽進程。隨著溫度的升高,甜高粱種子在不同濃度鹽脅迫下初始發芽時間均提前,由種子開始處理后的第3 d提前至第1 d,發芽結束時間由第6 d提前至第3 d;隨著鹽濃度的增加,NaCl處理延后了種子萌發的起始時間,到達發芽高峰期的時間也推遲。在30/20和35/25 ℃條件下,300 mmol·L-1的NaCl處理種子萌發后會出現死苗的現象。由此表明,溫度升高可以加快甜高粱種子的發芽速率,使發芽歷時縮短,而鹽脅迫推遲種子萌發的進程,在高溫和高濃度NaCl處理的交互作用下,會嚴重傷害甜高粱種子的萌發。
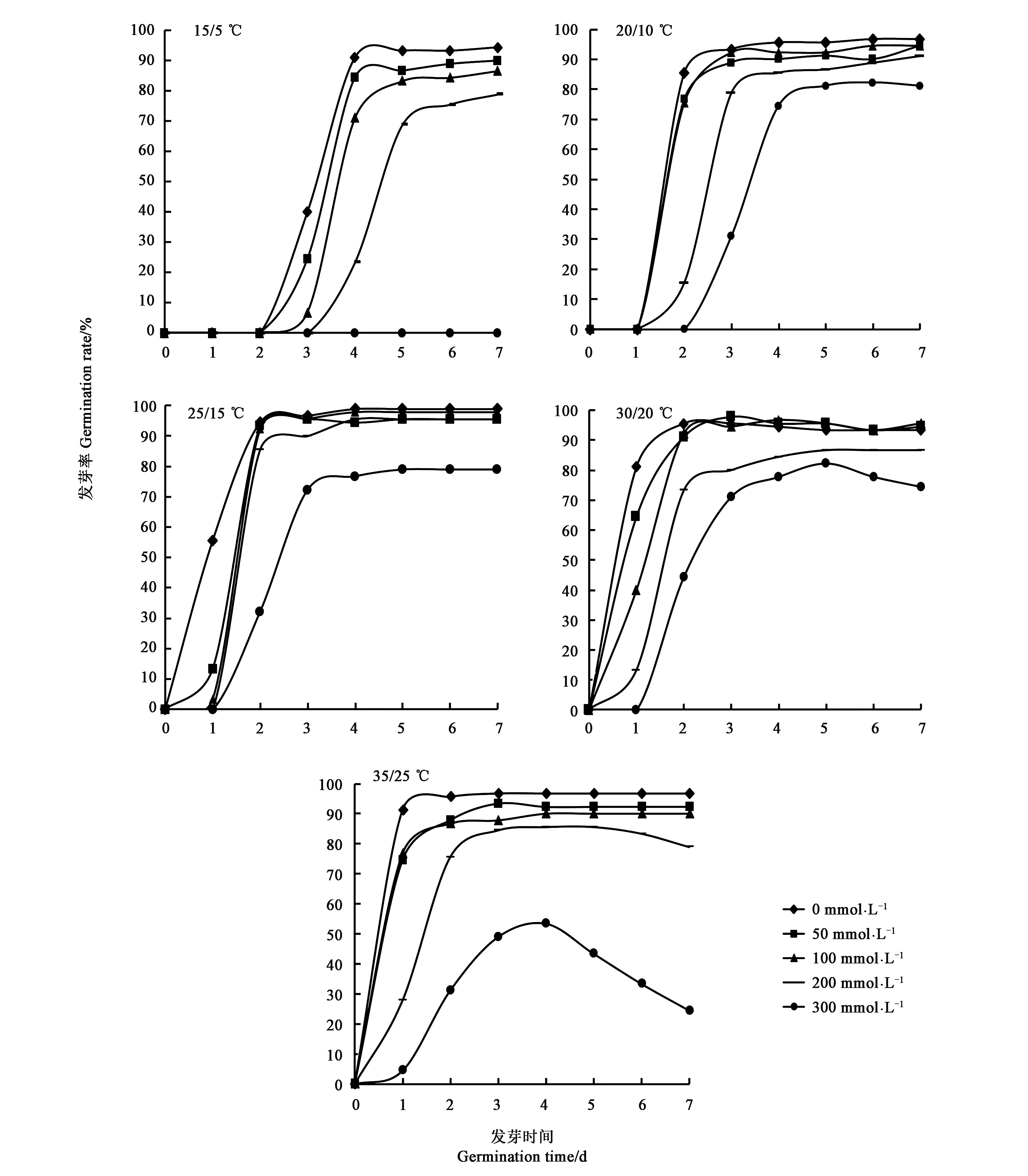
圖1 溫度與NaCl處理對甜高粱種子發芽進程的影響
2.2 不同處理對甜高粱種子萌發指標的影響
不同溫度和不同NaCl處理的甜高粱種子發芽率和發芽勢結果見圖2,可見,當NaCl濃度相同時,種子發芽率和發芽勢在25/15 ℃最高,其次為30/20 ℃,15/5 ℃時發芽率和發芽勢最低,雖然有的處理差異不顯著;當溫度相同時,鹽濃度越高,發芽率和發芽勢越低,15/5 ℃下,當NaCl濃度超過100 mmol·L-1時,發芽率和發芽勢顯著低于0 mmol·L-1處理,20/10、25/15、30/20、35/25 ℃條件下,鹽濃度超過200 mmol·L-1時,發芽率和發芽勢顯著低于0 mmol·L-1處理。由圖2可知,NaCl處理下,種子的發芽指數在35/25 ℃時最高,種子的活力指數在30/20 ℃時最高,二者均在15/5 ℃條件下最低,且顯著低于30/20、35/25 ℃處理;在溫度相同的條件下,隨著NaCl濃度的增加,種子的發芽指數和活力指數逐漸降低。

注:不同英文小寫字母表示相同溫度條件下不同NaCl處理間差異在P<0.05水平具有統計學意義;不同希臘小寫字母表示相同NaCl濃度處理下不同溫度間差異在P<0.05水平具有統計學意義。
溫度(T)、NaCl處理(S)及T×S對甜高粱種子的發芽指標貢獻度的雙因素方差分析結果(表1)表明,T、S及T×S對甜高粱種子的發芽率、發芽勢、發芽指數和活力指數的貢獻度不同,從T、S及T×S所占總平方和的百分比(PP)來看,S(53.53%)對發芽率的貢獻度較高,T×S(29.82%)次之,T(16.66%)最小;T(75.80%)對發芽勢的貢獻度較高,S(18.64%)次之,T×S(5.56%)最小;T(54.92%)對發芽指數的貢獻度較高,S(39.49%)次之,T×S(5.59%)最小;S(49.33%)對活力指數的貢獻度較高,T×S(33.39%)次之,T(16.68%)最小。因此,NaCl處理是影響發芽率和活力指數的主要因子,而溫度是影響發芽勢和發芽指數的主要因子。
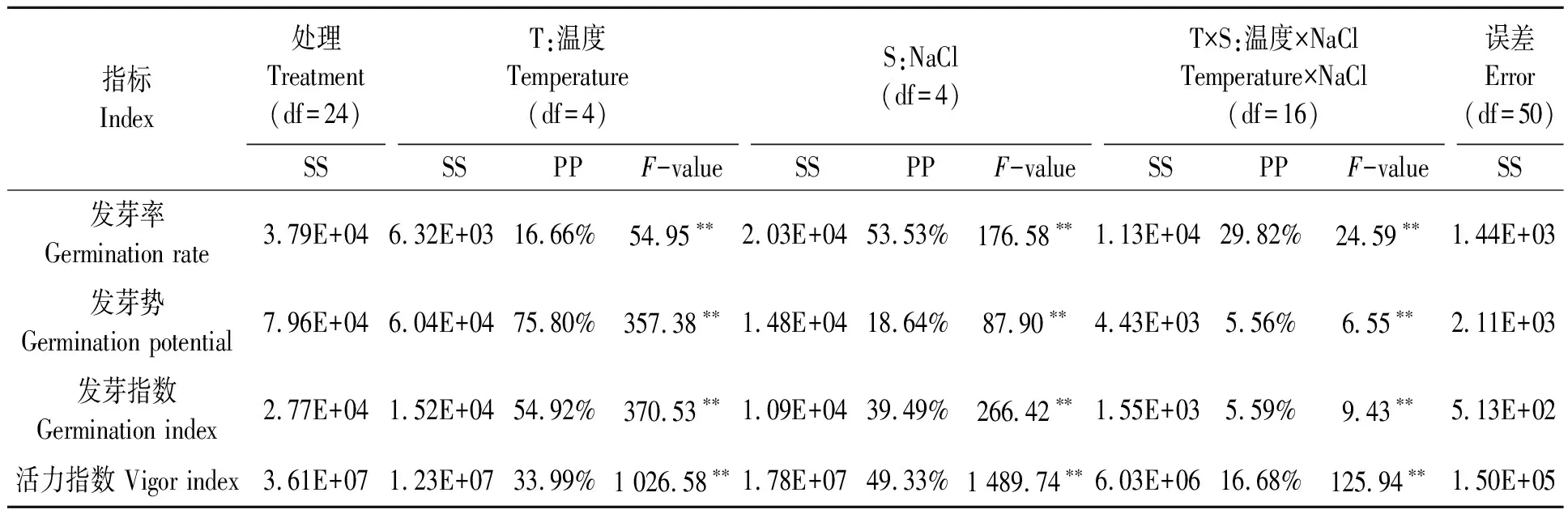
表1 溫度與NaCl處理對甜高粱種子萌發指標影響的雙因素方差分析
2.3 不同處理對甜高粱種子種苗轉化的影響
不同溫度和NaCl處理的甜高粱種子干物質轉移相關指標結果見圖3,當NaCl濃度相同時,芽干重+根干重在25/15 ℃最高,其次為30/20、35/25、20/10 ℃,15/5 ℃時最低;當溫度相同時,隨著鹽濃度升高,芽干重+根干重逐漸減小,當NaCl濃度超過100 mmol·L-1時,其顯著低于0 mmol·L-1處理。當NaCl濃度相同時,種子殘留干重在30/20、35/25 ℃下最低,15/5 ℃時最高;當溫度相同時,隨著NaCl濃度升高,種子殘留干重逐漸增大,當NaCl濃度超過50 mmol·L-1時,種子殘留干重顯著高于0 mmol·L-1處理。當NaCl濃度相同時,干物質轉運率在25/15 ℃、30/20 ℃、35/25 ℃下最高,其次為20/10 ℃,15/5 ℃時最低;當溫度相同時,隨著NaCl濃度升高,干物質轉運率逐漸降低,且不同濃度的NaCl處理均顯著低于0 mmol·L-1處理。當NaCl濃度相同時,總長和芽長在30/20 ℃達到最高,其次為25/15、35/25 ℃;鮮重、幼苗相對含水量和根長在25/15 ℃最高,其次為30/20、35/25 ℃,15/5 ℃時甜高粱早期幼苗的生長情況最差。可見,當溫度相同時,隨著NaCl濃度升高,鮮重、幼苗相對含水量、總長、芽長及根長逐漸減小,當NaCl濃度超過100 mmol·L-1時,均顯著低于0 mmol·L-1處理。
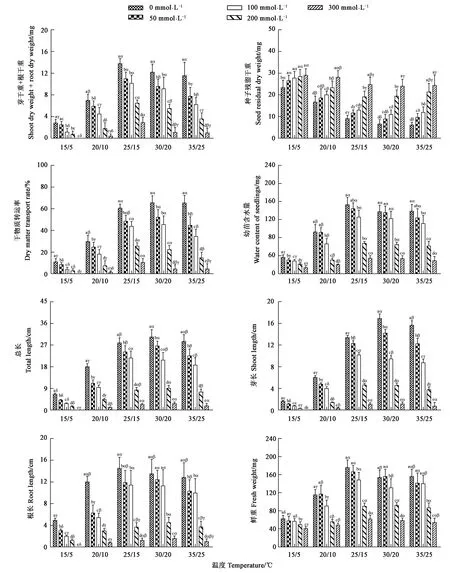
注: 不同英文小寫字母表示相同溫度條件下不同NaCl處理間差異在P<0.05水平具有統計學意義;不同希臘小寫字母表示相同NaCl濃度處理下不同溫度間差異在P<0.05水平具有統計學意義。
雙因素方差分析(表2)表明,T、S及T×S對甜高粱種子的干物質轉移貢獻度不同,從T、S及T×S所占總平方和的百分比(PP)來看,S(47.52%)對芽干重+根干重的貢獻度較高,T(44.80%)次之,T×S(7.69%)最小;T(48.04%)對種子殘留干重的貢獻度較高,S(41.79%)次之,T×S(10.17%)最小;S(45.49%)對干物質轉運率的貢獻度較高,T(43.52%)次之,T×S(10.99%)最小;S(47.52%)對甜高粱早期幼苗生長(鮮重、幼苗含水量、總長、芽長及根長)的貢獻度較高,T次之,T×S最小。因此,NaCl是影響芽干重+根干重、干物質轉運率、及早期幼苗生長主要因素的主要因子,溫度是影響種子殘留干重的主要因子。

表2 溫度與NaCl處理對甜高粱種子種苗轉化影響的雙因素方差分析
2.4 不同處理對甜高粱早期幼苗葉片抗氧化酶活性的影響
SOD、POD、CAT、APX是植物在逆境脅迫下為抵御外界傷害產生的抗氧化酶類。由圖4可知,在15/5和20/10 ℃下,SOD、POD、APX活性均隨著NaCl濃度的增加而逐漸減小,25/15、30/20、35/25 ℃時,3種酶活性在NaCl濃度增加的情況下呈現先增大后減小的趨勢;APX活性在不同溫度條件下均隨著NaCl濃度的增加而逐漸減小。在NaCl濃度相同的情況下,溫度對于4種酶活性的影響各不相同,在0、200 mmol·L-1時,4種酶活性均隨著溫度的升高呈現先增大后減小的趨勢,50、100 mmol·L-1時,4種酶活性均隨著溫度的升高而逐漸增大,300 mmol·L-1時,SOD、POD活性均隨著溫度的升高而逐漸增大,CAT、APX活性均隨著溫度的升高呈現先增大后減小的趨勢。
雙因素方差分析(表3)表明,對甜高粱早期幼苗葉片的抗氧化酶活性貢獻度不同,從T、S及T×S所占總平方和的百分比(PP)來看,S對SOD、POD、CAT活性的貢獻度較高,T次之,T×S最小;T對APX活性的貢獻度較高,S次之,T×S最小。因此NaCl處理是影響SOD、POD、CAT活性的主要因子,溫度是影響APX活性的主要因子。
3 討論
種子萌發過程中,溫度是調控其萌發的重要生態因子之一,同時也是鹽脅迫下種子萌發的重要生態條件[15]。在本研究中,NaCl濃度不超過100 mmol·L-1時,除15/5 ℃外,不同溫度對甜高粱種子的發芽率無顯著影響,說明甜高粱在低濃度的鹽脅迫下能夠適應多種變溫范圍。隨著NaCl濃度的增加,甜高粱在高溫(25/15~35/25 ℃)條件下具有較高的發芽率和發芽勢,15/5 ℃對甜高粱種子的發芽率和發芽勢抑制最強,這是因為在種子萌發過程中,種子內部進行著活躍的物質代謝反應,在一定范圍內,相對較高的溫度對種子萌發具有促進作用,超過某一特定溫度范圍的過高和過低溫度均不利于種子萌發[16]。在300 mmol·L-1濃度的NaCl脅迫下,30/20和35/25 ℃條件對甜高粱已萌發的早期幼苗有致死現象,說明過高的溫度、NaCl脅迫及其交互作用會影響甜高粱種子萌發的一系列生理生化過程,進而影響發芽率和發芽勢,當環境溫度適宜時,則可以減輕一定程度鹽脅迫對甜高粱種子萌發的毒害作用,這與小麥[15]和野大麥[17]的研究結果一致。
植物從種子轉化到幼苗過程中所需的物質及能量是由種子中貯藏的有機物提供,因此在種子萌發過程中總長、芽長、根長、鮮重、種子殘留干重、芽干重+根干重和干物質轉移率能體現出植物種苗轉化狀況,當植物早期幼苗胚芽和胚根生長均衡時,可以表現出更好的生長狀況[9]。本研究發現,在同一NaCl濃度下,除種子殘留干重外,甜高粱種苗轉化過程的總長、芽長、根長、鮮重、芽干重+根干重、干物質轉移率和幼苗含水量隨著溫度的升高呈先增大后減小趨勢,種子殘留干重則隨著溫度的升高呈減小趨勢;在同一溫度下,隨著NaCl濃度的增加,甜高粱種苗轉化過程除種子殘留干重外,其余指標均逐漸減小,種子殘留干重則隨著NaCl濃度的升高而增大;同時根長、鮮重、芽干重+根干重和幼苗含水量在25/15 ℃條件下最高,總長和芽長在30/20 ℃條件下最高,干物質轉運率在25/15和30/20 ℃條件下較高,種子殘留干重在30/20和35/25 ℃條件下較低,說明溫度、NaCl脅迫及其交互作用對種苗轉化的各指標影響不同,這可能是由于種子吸水后,較快地進行代謝活動,種子的水解作用增強,合成作用減弱,種子內部的淀粉和蛋白質水解為可溶性糖和可溶性蛋白等滲透調節物質,當超過種子萌發適宜溫度時,高溫加劇種子的水解和呼吸作用,導致種子內部的淀粉消耗增加,而溫度的升高對于早期幼苗的生長又是一種傷害。因此,在溫度與鹽脅迫的交互作用下,種子萌發時期種苗轉化過程中各指標對于脅迫的響應各有不同[15]。
非生物脅迫下,植物會產生大量活性氧自由基,大量的活性氧會影響蛋白質的合成、破壞膜系統,造成植物細胞嚴重損壞甚至死亡,SOD、POD、CAT和APX是細胞內清除活性氧的主要保護酶,整個保護酶系統防御能力的變化取決于這幾種酶的彼此協調的綜合結果[18]。本研究中,甜高粱早期幼苗葉片的抗氧化物酶活性整體上隨著溫度的升高逐漸增強,在適宜溫度下,酶活性隨著NaCl濃度的增加呈現先升高后下降趨勢,在較低或較高溫度下,酶活性隨著NaCl濃度的增加呈現降低趨勢。這可能是由于當甜高粱種子轉化為早期幼苗后,低溫可以抑制幼苗的一系列生理生化活動,而高溫引起水分蒸發加速,并通過毛細運動增加了鹽濃度,在鹽脅迫的基礎上加劇了對早期幼苗的傷害,而這些酶活性的增強有利于及時清除活性氧自由基,保持平衡,而隨著脅迫程度的進一步增大,抗氧化物酶已不能完全清除存在的活性氧自由基,平衡被打破,因此酶活性降低[19]。因此,在溫度與鹽脅迫交互作用下,脅迫傷害效應疊加,加重甜高粱早期幼苗受損程度,表現為低溫和高鹽脅迫傷害較大,抗氧化物酶活性降低,而高溫和低鹽脅迫提高了抗氧化物酶活性,這與類延寶[20]在小麥上的研究結果一致。
綜上所述,溫度與NaCl處理及其互作均可顯著影響甜高粱種子的萌發及早期幼苗的生長,低濃度NaCl處理下,低溫是抑制其萌發的主要原因,當NaCl濃度較高時,高溫更是加劇了對種子萌發的傷害作用,在30/20 ℃下,大多數指標達到最優狀態,表明該溫度為鹽脅迫下甜高粱種子萌發及早期幼苗生長的最適溫度。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
