蹄葉橐吾萃取物通過NF-κB通路降低細胞炎癥反應*
2020-03-13 03:09:08張春艷劉萬全呂艷欣
世界科學技術-中醫藥現代化 2020年6期
張春艷,劉萬全,呂艷欣,陳 萍,董 靜,王 玉,李 濤
(齊齊哈爾醫學院 黑龍江 161006)
炎癥是由激活的巨噬細胞釋放一系列炎癥因子所產生的一種以防御為主的免疫反應[1-2],但是炎癥因子的持續刺激會對組織產生損害,使機體功能喪失、引發腫瘤,嚴重的甚至導致死亡[3]。因此,開發一種新型、高效、無副作用的天然抗炎藥物具有深遠意義。
蹄 葉 橐 吾(Ligularia fischeri,LF)為 菊 科(compasitae)蹄葉橐吾屬(Ligularia)多年生草本植物,又名腎葉橐吾、馬蹄葉、葫蘆七等[4]。該植物性溫,味辛、苦。其根和根莖收入吉林省藥材標準,名山紫菀,具有溫肺下氣、理氣活血、止痛、止血、利尿、鎮咳祛痰等功效,用于治療慢性支氣管炎、咽喉炎癥、肺癰咳血、跌打損傷等疾病。全草用于治療丹毒性炎癥和關節膿腫,地上部分外敷用于捻挫傷,水煎服可治療痔疾[5-7]。,隨著LF 的藥理活性被逐漸闡明,發現除了其根和根莖具有鎮咳祛痰等藥理活性外,其葉片提取物中的倍半萜類及異蜂斗菜酮等化學成分[8-10]在抗炎免疫,抗腫瘤、抗氧化等方面顯示出潛在的藥用價值[11-13]。目前國內外學者已發現LF 葉片提取物可通過多條信號通路發揮抗炎作用,例如:抑制LPS 刺激RAW264.7 細胞產生一氧化氮和前列腺素-2[11]、通過c-Jun 氨基末端激酶(c-jun N-terminal kinase,JNK)和絲裂原活化蛋白激酶p38(Mitogen-activated protein kinases p38,p38MAPK通路下調炎性蛋白JNK、環氧酶(cyclooxygenase-2,COX-2)的表達,同時降低JNK 的磷酸化水平[12-13]。然而目前對于LF 葉片提取物的抗炎作用機制研究相對較少,深入的機制還有待于進一步挖掘。
細胞核內因子κB(nuclear factor-κB,NF-κB)是一種介導炎癥反應的重要轉錄調控因子,能夠與p38MAPK、JNK 和胞外信號調節激酶(extracellular signal regulated kinase,ERK)等通路共同作用,調節下游靶基因的轉錄和表達。NF-κB 主要由NF-κB 1(p105/p50)、NF-κB 2(p100/p52)、RelA(p65)、RelB 和c-Rel組成,其中p65在調節炎癥反應中發揮了重要的作用[14]。細胞在靜息狀態下,NF-κB 的核定位序列被其阻遏蛋白(inhibitor of NF-κB,IκB)封閉,以無活性的形式存在于細胞漿中[15]。傳統觀點認為,當細胞受到LPS 刺激時,IκB 發生磷酸化,與NF-κB 解離,進而暴露NF-κB 的核定位序列,使NF-κB 被激活,從細胞漿易位至細胞核,介導各種炎性因子的轉錄和表達[16]。除此以外,大量的實驗數據表明,NF-κB的亞基p65可以被細胞內的激酶磷酸化,這種磷酸化修飾可以反式激活NF-κB,從而增強NF-κB 的轉錄活性[15]。NF-κB抑制劑PDTC 能夠抑制NF-κB 的激活,從而阻止NFκB入核,達到抗炎作用[17]。雖然研究已表明EEOLF具有顯著的抗炎活性,但其能否通過NF-κB 信號通路達到抗炎作用尚不明確。本研究以LPS誘導的小鼠巨噬細胞RAW264.7 為炎癥模型,通過此模型,初步探討NF-κB通路介導的EEOLF抗炎作用機制,以期為開發EEOLF天然抗炎藥物提供有力依據。
1 材料和方法
1.1 材料與試劑
1.1.1 細胞
小鼠巨噬細胞RAW264.7購于北京協和醫學院。
1.1.2 藥品
LF為李濤教授在延邊市安圖縣采摘,EEOLF在齊齊哈爾大學制備,制備過程為干燥的LF 葉片8.5 kg,粉碎后,室溫下,每次用95%無水乙醇15 L 浸泡3 天后加熱回流,重復3 次,合并醇提液,減壓濃縮至小體積(約1.0 L),真空干燥,加水1.5 L,得醇提取物用水混懸,用乙酸乙酯萃取3 次,每次用乙酸乙酯1.5 L,合并乙酸乙酯層濃縮至恒重,得率為0.61%[15]。每克相當于10 g生藥,使用時DMSO 溶解后用PBS稀釋,濾過除菌,并分裝儲存。
1.1.3 試劑
FBS胎牛血清(美國Hyclone公司),乙二胺四乙酸(EDTA,美國Sigma 公司),四甲基二乙胺(TEMED,美國Sigma 公司),DMEM 培養基干粉(美國Gibco 公司),NF-κB抗體(美國Cell Signal Technology 公司)、NF-κB磷酸化抗體(美國Cell Signal Technology 公司)、β-actin(美國Cell Signal Technology 公司),山羊抗小鼠lgG/辣根酶標記二抗(美國Cell Signal Technology 公司),LPS(美國Sigma 公司),NF-κB 抑制劑PDTC(上海譜振生物科技有限公司),L-1β及TNF-α ELISA試劑盒(博士德生物科技有限公司)。
1.2 儀器
DY5000X 倒置顯微鏡(重慶澳浦光電技術有限公司),BIO-RAD 680 全自動酶標儀(美國BIO-RAD 公司),超低溫冰箱(美國Thermo Fisher Scientific 公司),3111 型二氧化碳培養箱(美國Thermo Fisher Scientific公司),VS-1300L-U 凈化工作臺(蘇州凈化設備有限公司),Tanon 5200 化學發光成像系統(上海天能科技有限公司)。
1.3 試驗方法
1.3.1 細胞培養及處理
取RAW264.7 巨噬細胞在DMEM 培養基(含10%胎牛血清)培養2 天、處于對數生長期、生長狀態良好的RAW264.7 細胞,經1 × PBS 清洗兩次,每次3 m L,用刮刀刮取細胞,并用細胞培養液吹打混勻,于細胞計數板上計數,加培養液稀釋至濃度為5×104個/mL,接種于96孔板中,每孔100μL,繼續培養,待巨噬細胞處于對數生長期且形成致密融合單層后分組處理。先前研究采用噻唑藍比色法(MTT 法)測得EEOLF 作用于密度為5×104個/mL的RAW264.7細胞24 h后,藥物對細胞的無毒濃度為5 μg·mL-1[12]。本研究取對數期細胞6 瓶,分別進行6 種處理。空白對照組,加入不含血清培養基培養24 h;EEOLF 處理組,加入EEOLF(終濃度為5μg·mL-1)培養24 h;LPS 刺激組,加入LPS(終濃度為0.1 mg·L-1)培養24 h;LPS+PDTC 組,加入PDTC(終濃度為10 ng·μL-1)培養1 h 后,再加入LPS(終濃度為0.1 mg·L-1)培養24 h;LPS + EEOLF 組,加入EEOLF(終濃度為5 μg·mL-1)培養1 h 后,再加入LPS(終濃度為0.1 mg·L-1)培養24 h;LPS + PDTC +EEOLF 組,加入PDTC(終濃度為10 ng·μL-1)培養1 h后,再加入EEOLF(終濃度為5μg·mL-1)培養1 h,最后加入LPS(終濃度為0.1 mg·L-1)培養24 h。
1.3.2 免疫印跡法分析蛋白表達
取RAW264.7 巨噬細胞在DMEM 培養基(含10%胎牛血清)培養2 d、處于對數生長期、生長狀態良好的RAW264.7 細胞,經1 × PBS 清洗兩次,每次3 m L,用刮刀刮取細胞,并用細胞培養液吹打混勻,于細胞計數板上計數,加培養液稀釋至濃度為5×104個/mL,接種于96孔板中,每孔100μL,繼續培養,待巨噬細胞處于對數生長期且形成致密融合單層后按照1.3.2 的方法分組處理。提取細胞核總蛋白,并測定各組的總蛋白濃度,95 ℃將蛋白變性,取30 μg 蛋白樣品進行電泳,將蛋白轉移到硝酸纖維素膜(0.45μm),用脫脂牛奶室溫封閉2 h,TBST 洗3 次。分別加入一抗NF-κB p65、p-p65 和β-actin 抗體,抗體稀釋度為1:1000,4 ℃孵育過夜;TBST 洗3 次,加入辣根酶標記山羊抗小鼠二抗,抗體稀釋度為1∶5000,室溫孵育1 h,TBS 洗滌2 次,再次TBST 洗滌后反應信號經ECL 底物化學發光檢測,結果掃描后,以β-actin 為內參,用ImagePro 分析每個目的蛋白特異灰度值并計算相對灰度值。以上實驗重復3次。
1.3.3 細胞因子檢測
取RAW264.7 巨噬細胞在DMEM 培養基(含10%胎牛血清)培養2 d、處于對數生長期、生長狀態良好的RAW264.7 細胞,經1 × PBS 清洗兩次,每次3 m L,用刮刀刮取細胞,并用細胞培養液吹打混勻,于細胞計數板上計數,加培養液稀釋至濃度為5×104個/mL,接種于96孔板中,每孔100μL,繼續培養,待巨噬細胞處于對數生長期且形成致密融合單層后按照1.3.2 的方法分組處理。用ELISA 試劑盒檢測上清腫瘤壞死因子TNF-α 和白介素IL-1β 分泌情況。將8 pg·mL-1、400 pg·mL-1、200 pg·mL-1、100 pg·mL-1、50 pg·mL-1L、25 pg·mL-1L、12.5 pg·mL-1的標準品各100 uL 依次加入孔中,另取1孔只加細胞培養上清作為陰性對照,每個濃度設置三個復孔;酶標板加上封板膜,37 ℃反應90分鐘;依次加入生物素抗小鼠IL-1β抗體100 uL;酶標板加上封板膜,37 ℃反應60 分鐘;洗滌緩沖液洗滌3 次,每次浸泡1 分鐘左右(每孔洗液至少300 uL);將準備好的ABC 工作液按每孔100 uL 依次加入;酶標板加上封板膜,37 ℃反應30分鐘;洗滌緩沖液洗滌5 次,每次浸泡1-2 min 左右(每孔洗液至少300 uL)。分別加入顯色劑A 和B,輕輕震蕩混勻,37 ℃避光顯色10 min;加終止液,采用酶標儀測量各孔的吸光度A450nm。
1.4 統計學分析
炎癥因子釋放數據采用GraphPad Prism 5 軟件分析,Western blot 數據采用ImagePro 軟件分析,數據以±SD表示,P<0.05為差異具有統計學意義。
2 結果與分析
2.1 RAW264.7巨噬細胞形態學觀察
RAW264.7 巨噬細胞在DMEM 培養基(含10%胎牛血清)培養2天,待巨噬細胞處于對數生長期且形成致密融合單層后分組處理,用以觀察比較EEOLF 及PDTC 作用后細胞的形態變化。各組RAW264.7 細胞如圖1 所示,正常RAW264.7 細胞形態以類圓形為主,邊緣明亮,含有1~2 個細胞核,偶有細長觸角,是一類胞體體積較小的細胞(圖1A)。單獨加入EEOLF 生長24 h 后,細胞無明顯變化,細胞核清楚,狀態良好(圖1B)。加入LPS 誘導24 h 后,細胞模糊不清,細胞內有空泡(圖1C)。用PDTC 或者EEOLF 單獨作用于LPS誘導的RAW264.7 細胞后,細胞內空泡減少(圖1D 和1E),用PDTC 和EEOLF 同 時 作 用 于LPS 誘 導 的RAW264.7 細胞后,細胞核變得清晰,細胞內空泡減少(圖1F),說明EEOLF 和NF-κB 抑制劑PDTC 效果相似。
2.2 EEOLF降低LPS誘導的RAW264.7巨噬細胞NFκB p65表達及其磷酸化
NF-κB p65 在正常情況下位于細胞漿中,當發生炎癥反應時,將會從細胞漿易位至細胞核內[16]。本研究通過構建LPS 誘導的RAW264.7 細胞炎癥模型,Western 法檢測細胞核內NF-κB p65 蛋白表達水平及其磷酸化水平,間接檢測藥物是否可以通過NF-κB 信號通路達到抗炎作用。研究結果顯示,與空白對照組相比,LPS 組NF-κB p65 和p-p65 蛋白表達量顯著升高(###P<0.001),說明LPS誘導的RAW264.7細胞發生了炎癥反應。加入PDTC 或者EEOLF,分別作用于LPS 誘導的RAW264.7 細胞,發現NF-κB p65 和p-p65蛋白表達量明顯低于LPS 組(***P < 0.001),表明EEOLF 與PDTC 一樣,可以降低LPS 誘導的RAW264.7巨噬細胞NF-κB p65 和p-p65 蛋白表達。當PDTC 與EEOLF 共同作用于LPS 誘導的RAW264.7 細胞后,NF-κB p65 和p-p65 蛋白表達量低于PDTC 或EEOLF單獨作用(△△△P < 0.001 或△P < 0.05),說明在NF-κB抑制劑PDTC 與EEOLF 的共同作用下,對細胞核內NF-κB p65 和p-p65 蛋白表達的抑制作用進一步加強(***P<0.001)(圖2)。
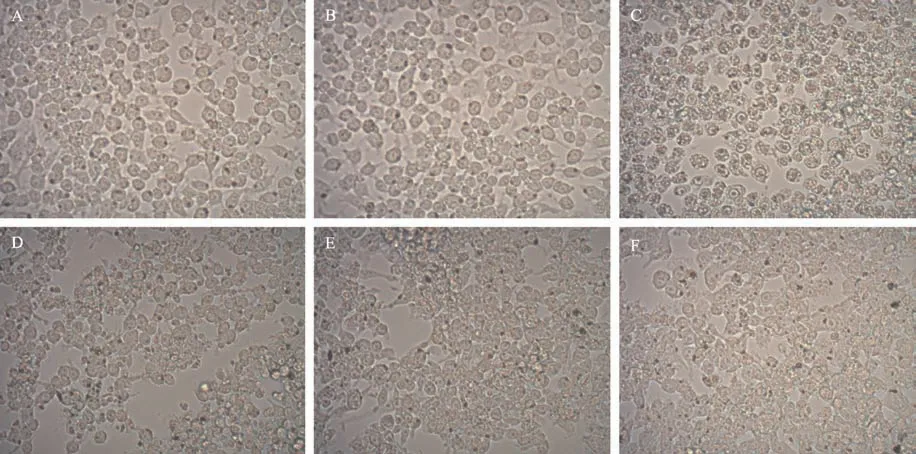
圖1 不同因素處理后的RAW264.7的細胞形態

圖2 EEOLF對RAW264.7細胞NF-κB p65和p-p65蛋白表達的影響
2.3 EEOLF 降低LPS 誘導的RAW264.7 巨噬細胞炎癥因子的分泌
NF-κB 信號轉導通路被激活后,會介導下游各種炎性因子的表達,如:TNF-α、IL-1 等,因此炎性因子分泌量的多少反應了炎癥的嚴重程度。本研究結果顯示,與空白對照組相比,LPS 組TNF-α 和IL-1β 的分泌 量 顯 著 升 高(###P < 0.001),說 明LPS 誘 導 的RAW264.7 細胞發生了炎癥反應。加入PDTC 或者EEOLF,分別作用于LPS 誘導的RAW264.7 細胞后,發現TNF-α 和IL-1β 的分泌量明顯減少(***P < 0.001),表明EEOLF 與PDTC 均能降低炎癥細胞中TNF-α 和IL-1β 的釋放。當PDTC 與EEOLF 共同作用于LPS 誘導的RAW264.7 細胞后,TNF-α 和IL-1β 的分泌量更低(△△△P<0.001),表明PDTC與EEOLF共同作用后對炎癥因子TNF-α和IL-1β釋放的抑制作用進一步加強(***P<0.001)(圖3)。

圖3 不同因素處理后的RAW264.7細胞炎性因子的表達
3 討論
炎癥是一類極其復雜的病理生理過程,是機體對損傷性刺激最原始的保護性反應,在一定程度內起到保護機體的作用,但長期劇烈的炎癥反應會加速疾病進程,使得機體出現中毒性休克,多器官功能衰竭等癥狀[18-19]。
LF 為我國傳統中藥,始載于《神農本草經》,又稱山紫菀,李時珍說:“其根色紫而柔宛故名”。主要分布在東北長白山、內蒙古、朝鮮、日本、前蘇聯等,因其藥理作用廣泛、廉價易采摘等優勢而具有很大的開發價值[20]。近年來研究發現除了其根和根莖具有鎮咳祛痰等藥理活性外,LF地上部分乙醇提取物對降低炎癥組織中的PGE2 的含量、抑制小鼠單核巨噬細胞吞噬功能、抑制組胺、5-羥色胺、緩激膚等炎癥介質的合成或釋放及增強體液免疫功能有關。但具體作用機制尚不十分清楚[21]。本研究以RAW264.7 細胞作為載體,采用LPS 建立體外細胞炎癥模型,探討EEOLF 對LPS 誘導的RAW264.7 細胞功能的調控作用,以豐富EEOLF 的抗炎作用機制,為LF 的臨床應用提供理論基礎。
本研究在顯微鏡下觀察到LPS 刺激RAW264.7 細胞24 h 后細胞邊緣模糊不清,細胞內出現空泡(圖1C)。用PDTC 或者EEOLF 單獨作用于LPS 誘導的RAW264.7細胞后,細胞內空泡減少(圖1D 和1E)。用PDTC 和EEOLF 同時作用于LPS 誘導的RAW264.7 細胞后,細胞核變得清晰,細胞內空泡減少(圖1F),提示EEOLF 與NF-κB 抑制劑PDTC 功能相似,均可使LPS誘導的RAW264.7細胞發生凋亡。
LPS 是革蘭氏陰性菌細胞壁的成分,能夠作為炎癥誘導劑,與細胞膜表面受體(Toll-like receptor 4,TLR4)結合[22-24],刺激巨噬細胞等免疫細胞的活化,引起全身的炎癥反應。LPS誘導的炎癥反應主要是通過激活NF-κB等信號通路實現的,NF-κB 被激活后進入細胞核,與特異性的基因轉錄起始區結合,啟動炎癥基因的轉錄,介導TNF-α、IL-1 等炎癥因子的釋放[15]。炎癥因子的分泌量是檢測炎癥嚴重程度的量化指標,炎癥標志因子TNF-α 在炎癥網絡中具有關鍵作用,是全身炎性反應的始動介質,是炎癥反應發生的經典檢測指標,TNF-α 釋放量的多少直接反應炎癥的輕重[25-26]。而IL-1 一般產生于炎癥早期,具有介導炎癥產生的作用[3]。NF-κB 的活化對于TNF-α、IL-1 等炎癥因子的的表達是必需的[27],因此抑制NF-κB 的活化很可能是治療炎癥的有效方法。
本研究結果顯示,EEOLF 作用于LPS 誘導的RAW264.7 細胞后,可下調NF-κB p65 的蛋白表達及磷酸化水平,同時減少TNF-α、IL-1 等炎癥因子的產生。而使用陽性對照藥PDTC 后,NF-κB p65 的蛋白表達及磷酸化水平同樣受到抑制,相應地,TNF-α、IL-1 等炎癥因子的分泌也受到抑制。進一步揭示了EEOLF 可通過抑制NF-κB 的激活,抑制LPS 誘導的炎癥反應。因此,以NF-κB 信號通路為切入點,研究EEOLF 作用于LPS 誘導的巨噬細胞后該通路的活性,能夠為開發抗炎、抗免疫作用的藥物提供有力的理論基礎,為慢性免疫炎癥性疾病患者帶來福音。
另外,本研究還發現當EEOLF 單獨作用于RAW264.7 細胞后,NF-κB 表達水平升高,這可能是因為藥物作用于細胞后,細胞發生了應激反應。因為當細胞受到各種理化及生物性刺激時,將出現一系列適應代償性反應,這些反應包括一系列非特異性反應,統稱為細胞應激。而研究報道稱NF-κB 是幾種細胞應激信號核轉導因子之一,細胞在受到外界刺激后,可通過經典途徑或旁路途徑激活NF-κB[28]。也就是說當細胞受到藥物刺激后,發生了應激反應,激活了NF-κB,從而導致NF-κB 表達量有所升高,TNF-α 表達量有所升高可能也與NF-κB激活有關[29]。
綜上所述,EEOLF 能減弱LPS 誘導的RAW264.7細胞炎癥,其機制可能與EEOLF 能夠調控NF-κB 通路有關,進而抑制TNF-α 和IL-1β 的釋放。但抗炎是一個綜合而復雜的過程,是多條信號通路共同作用的結果,具體作用機制還需進一步研究。
