芝麻種子萌發動態及其代謝生理變化研究
2020-03-15 03:26:00孫建周紅英樂美旺顏廷獻饒月亮顏小文梁俊超葉艷英
中國農業科技導報 2020年8期
孫建, 周紅英, 樂美旺, 顏廷獻, 饒月亮, 顏小文, 梁俊超, 葉艷英
(江西省農業科學院作物研究所,江西省油料作物生物學重點實驗室,國家油料改良中心南昌分中心,農業農村部長江中下游作物生理生態與耕作重點實驗室,南昌 330200)
芝麻(SesamumindicumL.)是重要優質油料作物,種子含油量高,不飽和脂肪酸占85%左右,富含芝麻素(sesamin)、芝麻林素(sesamolin)、維生素E等天然抗氧化類物質[1-3],是重要的優質食用油源和營養保健食品[4-5]。萌發生理是種子生物學的重要內容之一,在很多植物中開展了大量相關研究,包括種子萌發的逆境生理、營養及化學成分變化和代謝生理等方面。關于芝麻種子萌發過程的相關研究多集中在營養及化學成分[6-7]、芽苗生長與生理[8-9]等方面,關于代謝生理方面的報道不多,韓亞飛等[10]在30 ℃條件下對芝麻種子萌發0~48 h的總糖、粗脂肪、粗蛋白和氨基酸組成等變化進行研究,結果顯示,總糖含量先降后升,粗脂肪含量逐漸下降,粗蛋白含量變化不明顯,不同類型氨基酸占比變化不同。張瑞等[11]研究了35 ℃暗培養條件下芝麻種子萌發 0~72 h 的粗脂肪、粗蛋白、總糖和脂肪酸組成等的變化,結果顯示,粗脂肪含量在36 h后降低幅度較大,粗蛋白含量變化較小,總糖含量不斷增加,脂肪酸組成之間變化幅度較小。張麗霞等[12]研究了芝麻種子在28 ℃條件下萌發0~96 h的脂肪酸組成、氨基酸組成、粗脂肪、粗蛋白等主要成分的動態變化,發現粗脂肪含量在萌發的36 h以前變化較小,粗蛋白含量則呈現先降后升再降的變化,84 h后下降明顯,主要脂肪酸組分的變化不大,17種氨基酸變化各不相同,酸性氨基酸占比隨著種子萌發不斷增加,中性和堿性氨基酸不斷減少。這些研究主要針對芝麻種子萌發過程的營養成分變化,尚未從代謝生理的角度對芝麻種子主要貯藏物質參與代謝的先后順序進行研究。因此,本研究擬檢測芝麻萌發動態的形態學和水分需求變化,確定芝麻種子萌發的關鍵時間點,并進一步測定芝麻種子萌發過程中與代謝相關的可溶性糖、可溶性蛋白、氨基酸、游離脂肪酸及脂肪酶等生理指標變化,明確種子主要貯藏物質參與代謝的先后順序,在萌發動態和代謝生理上豐富芝麻種子生物學相關內容,并為芝麻種子萌發代謝機理的進一步研究提供參考。
1 材料與方法
1.1 試驗材料
供試材料為贛芝9號和贛芝12號,其中贛芝9號為黑芝麻品種,粗脂肪含量為50.5%,粗蛋白含量為19.7%;贛芝12號為白芝麻品種,粗脂肪含量57.1%,粗蛋白含量20.6%。種子均由江西省農業科學院作物研究所芝麻創新團隊提供,選取正常成熟、籽粒飽滿的芝麻種子備用。
1.2 試驗設計
試驗分兩步開展,均在黑暗狀態的25 ℃恒溫培養箱中進行。第一步用于觀察芝麻種子萌發動態,測定吸水率、吸水速率、粗脂肪含量和脂肪酸組成,將精選的干凈種子放置在鋪有3層濾紙的發芽盤中,每盤稱取10 g平鋪于濾紙上,確保種子籽粒之間不重疊,每盤等量地加入充足的蒸餾水以供萌發,分別在0、3、6、9、12、15、18、21、24、30、36、42、48、54、60、66和72 h取樣,每個時間點取樣3盤,即3次重復。每個樣品取出后測定鮮重,并于60 ℃烘箱烘干至恒重,稱量干重用于粗脂肪、脂肪酸組成的測定。根據第一步的發芽動態、吸水率和吸水速率測定結果,確定0、6、12、24、48、72 h共6個時間點進行第二步試驗,將精選的干凈種子放置在鋪有3層濾紙的發芽盒中,發芽盒長11 cm、寬11 cm、高6 cm,每盒放置種子200粒,同樣于25 ℃暗培養,每個時間點取樣3盒,即3次重復,將取樣樣品用蒸餾水沖洗干凈并用濾紙吸干水分備用,用于測定可溶性糖含量等生理指標。
1.3 測定指標與方法
吸水率為種子吸收水分的質量占種子干重的百分比,吸水速率為每g干種子每h吸收的水分質量。粗脂肪含量采取索氏抽提法(GB/T 14772-2008)[11]測定,采取氣相色譜法[11]測定棕櫚酸、硬脂酸、油酸和亞油酸4個主要脂肪酸組成成分。可溶性糖含量采用蒽酮比色法[13]測定,氨基酸含量采用水合茚三酮法[13]測定。可溶性蛋白含量采用BCA法[14]測定。脂肪酶活性通過銅皂-分光光度法測定脂肪酸生成速率并進行計算[15]。游離脂肪酸含量采取銅皂比色法[16]測定。所用試劑均購自蘇州科銘生物技術公司。
1.4 數據分析
Microsoft Excel 2007和SPSS 18.0軟件進行數據整理、相關性分析及作圖。
2 結果與分析
2.1 種子萌發動態及其水分變化
芝麻種子萌發動態見圖1,可見,25 ℃暗培養情況下,12 h芝麻種子實現萌動露白,24 h進入胚根快速伸長階段,48 h胚根和胚芽同步快速伸長生長,72 h子葉開始展開完成種子萌發過程,繼而進入芽期幼苗生長階段。
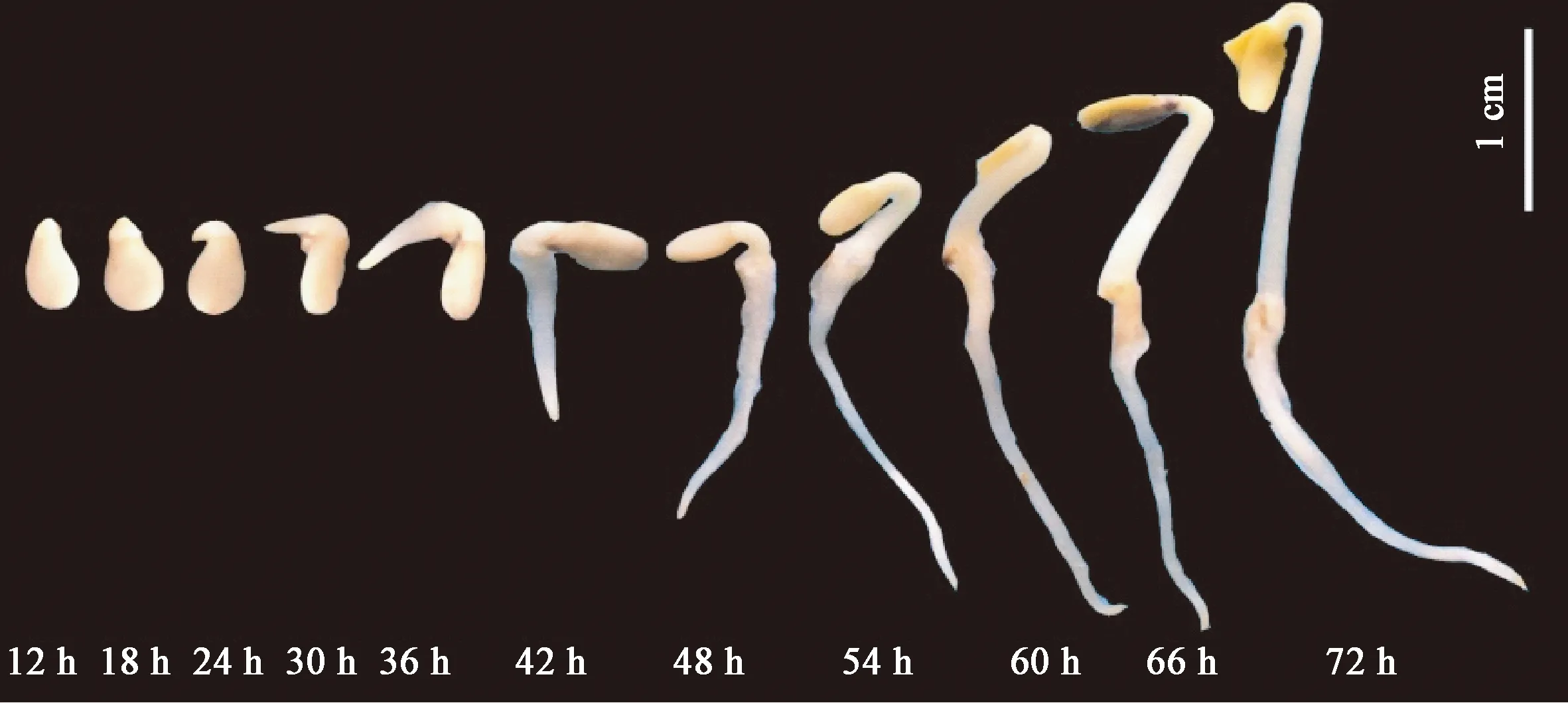
圖1 芝麻種子萌發過程的形態學觀察Fig.1 Germination morphology of sesame seed
芝麻種子萌發過程的吸水率和吸水速率結果(圖2)顯示,種子吸水率呈先急速上升后平穩增加接著再次快速上升的規律,呈現兩個快速吸水期,贛芝9號和贛芝12號3 h吸水率分別達到50.77%和44.19%,并在6 h后進入較為平穩增加階段,24~54 h,吸水率開始進入第二次快速上升階段,贛芝9號和贛芝12號分別迅速增加到162.55%和153.92%,接著增速有所減緩。吸水速率的變化呈現出由高吸水速率(贛芝9號和贛芝12號分別為169.23和147.12 mg·h-1·g-1)快速下降,9 h降到最低,隨后開始平穩上升,至 54 h 達到第二個峰值(贛芝9號和贛芝12號分別為51.51和45.79 mg·h-1·g-1),然后再次下降。兩個品種萌發過程中的吸水率和吸水速率的變化趨勢表現一致,但也存在較小差異,這可能與品種的種皮結構、物質組成和萌發特點等差異有關。
結合種子萌發形態和種子吸水率、吸水速率結果,可以確定25 ℃暗培養條件下,芝麻種子于浸水6 h時吸收的水分超過自身干重,基本滿足對水分的需求,達到種子萌發階段的“吸脹”,此前吸水率急劇上升,吸水速率最高。12 h進入種子萌發的第二階段“萌動”,即“露白”,對水分的需求趨于平穩,吸水率增加較慢,吸水速率在較低水平變化。隨后在24 h開始進入胚根伸長時期, 48 h后胚根和胚芽的下胚軸開始同步進入快速伸長生長時期,對水分的需求開始快速增加。72 h 時子葉基本展開,完成了種子萌發的第三階段“發芽”,隨后進入幼苗生長階段。
鑒于此,本研究下一步開展的代謝生理研究將圍繞干種子(0 h)、吸脹(6 h)、萌動(12 h)、胚根開始伸長(24 h)、胚芽的下胚軸開始伸長(48 h)和基本完成萌發(72 h)共6個時間點進行。
2.2 種子萌發過程的可溶性糖含量變化
可溶性糖是芝麻種子貯藏的主要碳水化合物之一,也是種子萌發過程中脂肪代謝的最終產物之一。圖3顯示,芝麻干種子中貯藏有一定量的可溶性糖,吸脹(6 h)時可溶性糖含量急劇下降,差異達顯著水平。隨著萌發過程的推進,可溶性糖含量呈先降后升的變化趨勢,品種間變化趨勢基本一致。種子吸脹(6 h)到胚芽下胚軸開始伸長生長(48 h),可溶性糖被不斷消耗,含量不斷降低,在完成發芽(72 h)時含量又有所增加,差異達到顯著水平。
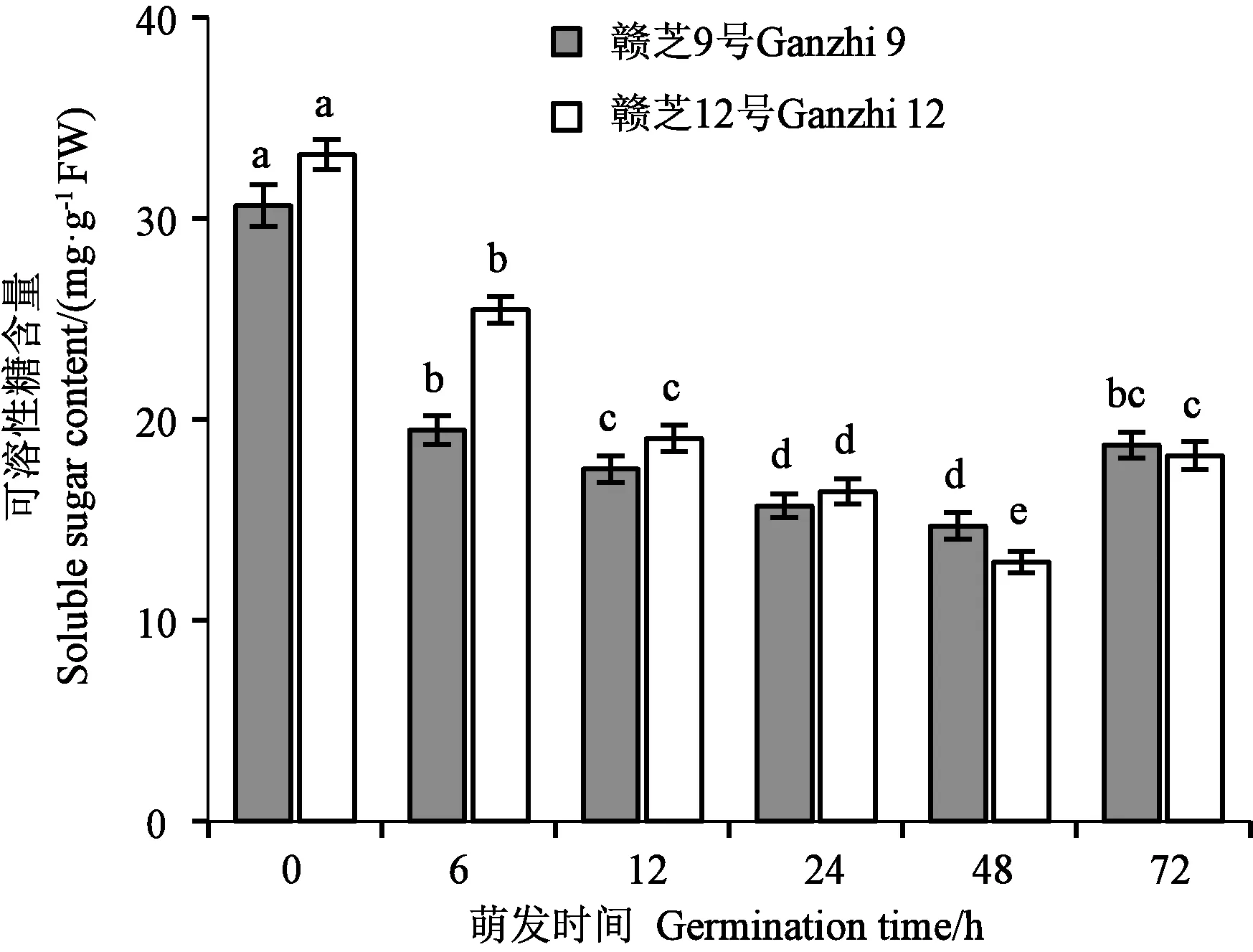
注:不同小寫字母表示同一品種不同萌發時間的差異在P<0.05水平具有統計學意義。Note: Different lowercase letters of the same variety indicate significant difference between different germination times at P<0.05 level.圖3 芝麻種子萌發過程的可溶性糖含量變化Fig.3 Changes of soluble sugar content during germination of sesame seed
2.3 種子萌發過程的蛋白質代謝
蛋白可溶化是貯藏蛋白進入代謝過程的第一步。2個供試品種種子萌發過程中,可溶性蛋白含量均呈先降后升的變化規律(圖4),吸脹(6 h)時可溶性蛋白含量較干種子顯著下降,然后隨著種子的萌發進程,可溶性蛋白含量不斷升高,6~72 h不同萌發時間的可溶性蛋白含量差異均達到顯著水平。
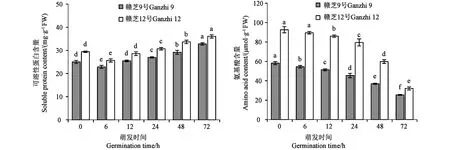
注:不同小寫字母表示同一品種不同萌發時間的差異在P<0.05水平具有統計學意義。Note: Different lowercase letters of the same variety indicate significant difference between different germination times at P<0.05 level.圖4 芝麻種子萌發過程中可溶性蛋白含量和氨基酸含量的變化Fig.4 Changes of soluble protein content and amino acid content during germination of sesame seed
作為蛋白質的代謝產物,氨基酸是參與生命活動的重要因子。芝麻種子萌發過程中,氨基酸含量呈現不斷下降趨勢(圖4),同一品種不同萌發時間的氨基酸含量差異均達到顯著水平。供試品種間的變化趨勢基本一致,但品種間的含量存在較大差異,贛芝12號的氨基酸含量明顯高于相同萌發時間的贛芝9號。
2.4 種子萌發過程的脂肪代謝
萌發過程中種子粗脂肪含量結果(圖5)顯示,從干種子(0 h)、吸脹(6 h)、萌動(12 h)到胚根伸長(24 h),種子的粗脂肪含量沒有顯著變化。胚芽伸長(48 h)后,粗脂肪含量開始顯著下降。在種子萌發過程中,游離脂肪酸含量表現為先急劇下降后不斷升高再快速下降的趨勢。在干種子(0 h)中存在一定含量的游離脂肪酸,在種子吸脹(6 h)時游離脂肪酸急劇下降到最低水平,其后隨著種子萌發進程的推進,游離脂肪酸含量不斷上升,在胚芽伸長(48 h)時達到最高值,然后在發芽(72 h)時再次快速下降。脂肪酶活性結果(圖5)顯示,種子萌發過程中脂肪酶活性不斷增加,品種間總體變化趨勢基本一致,但品種間存在差異。在吸脹(6 h)和萌動(12 h)時兩個供試品種間基本沒有差異,胚根伸長(24 h)后,贛芝12號的脂肪酶活性大幅度增加,而贛芝9號的脂肪酶活性增加較緩,這可能與品種的粗脂肪含量有關,高含油量的贛芝12號較低含油量的贛芝9號在種子萌發的脂肪代謝中需要更高活性的脂肪酶。
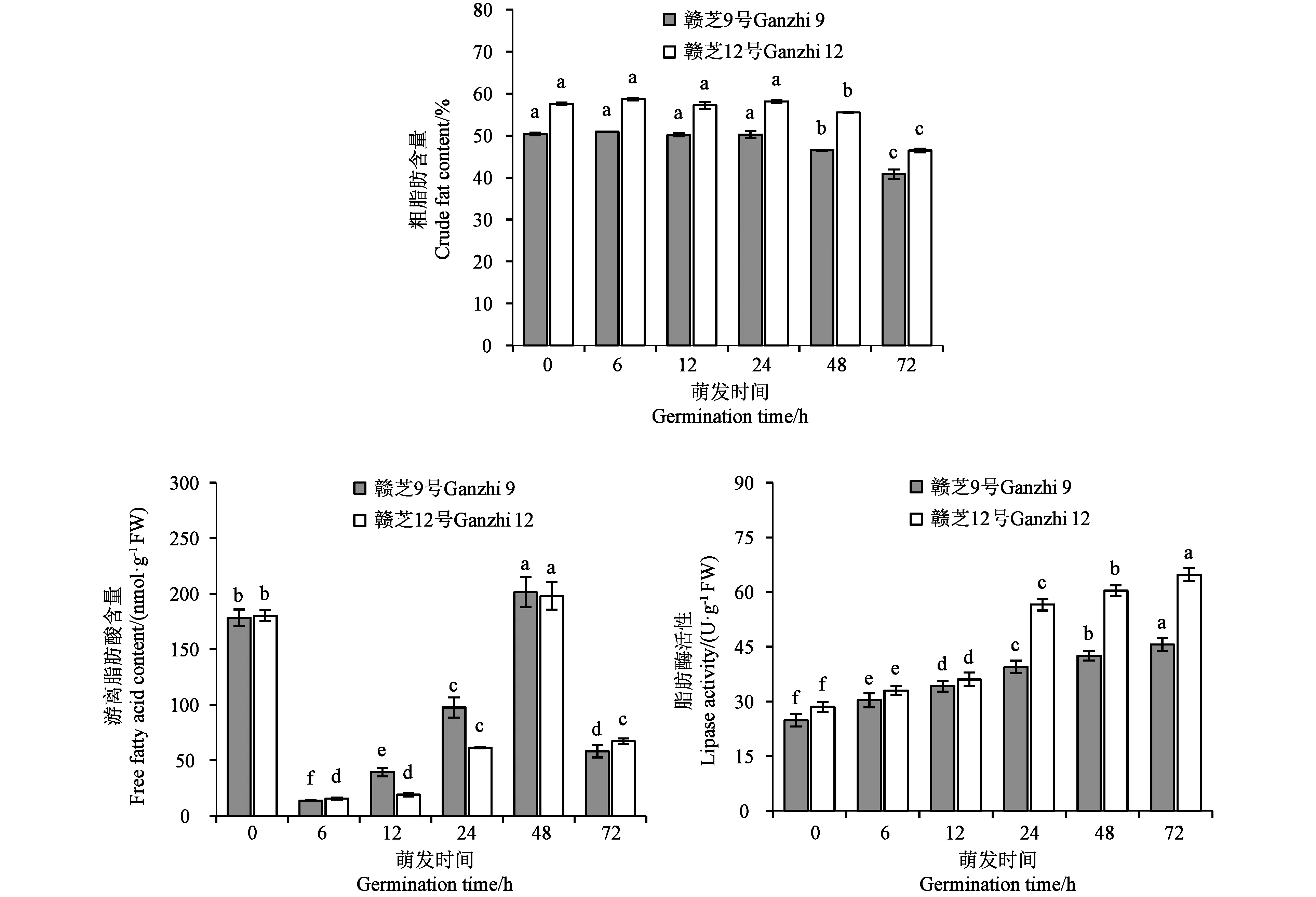
注:不同小寫字母表示同一品種不同萌發時間的差異在P<0.05水平具有統計學意義。Note: Different lowercase letters of the same variety indicate significant difference between different germination times at P<0.05 level.圖5 芝麻種子萌發過程中粗脂肪含量、游離脂肪酸含量和脂肪酶活性的變化Fig.5 Changes of crude fat content, free fatty acid content and lipase activity during germination of sesame
對種子萌發過程中的主要脂肪酸組成進行分析,結果(圖6)顯示,棕櫚酸、硬脂酸、油酸和亞油酸4種主要脂肪酸組成基本保持穩定,變化較小,這與前人的研究結果[11-12]一致。棕櫚酸在種子發芽(72 h)時有所降低,但降低幅度較小,分別為1.56%(贛芝9號)和1.94%(贛芝12號)。相關性分析顯示,油酸與亞油酸之間存在顯著負相關關系(r=-0.522,P<0.01)。
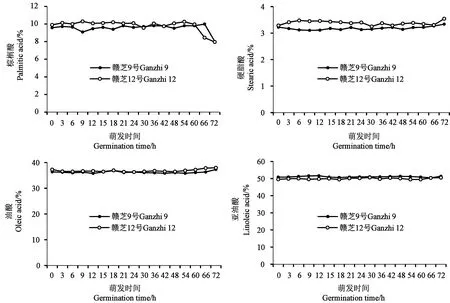
圖6 芝麻種子萌發過程的主要脂肪酸組成變化Fig.6 Changes of main fatty acid composition during germination of sesame seed
2.5 相關分析
對芝麻種子萌發過程中與代謝相關的可溶性糖、可溶性蛋白、氨基酸、粗脂肪、游離脂肪酸和脂肪酶活性共6個生理指標進行相關分析,結果(表1)顯示,可溶性糖含量與脂肪酶活性呈顯著負相關(P<0.05),粗脂肪含量與氨基酸含量呈極顯著正相關(P<0.01),脂肪酶活性與可溶性蛋白呈極顯著正相關(P<0.01)。蛋白質代謝中,可溶性蛋白含量與氨基酸含量呈負相關,與蛋白質代謝生成氨基酸有關。脂肪代謝中,脂肪酶活性與游離脂肪酸含量呈正相關,與粗脂肪含量呈負相關,粗脂肪含量與游離脂肪酸含量正相關,但三者相互間的相關系數均較小,可能與芝麻種子萌發0~72 h內脂肪代謝量較低有關。
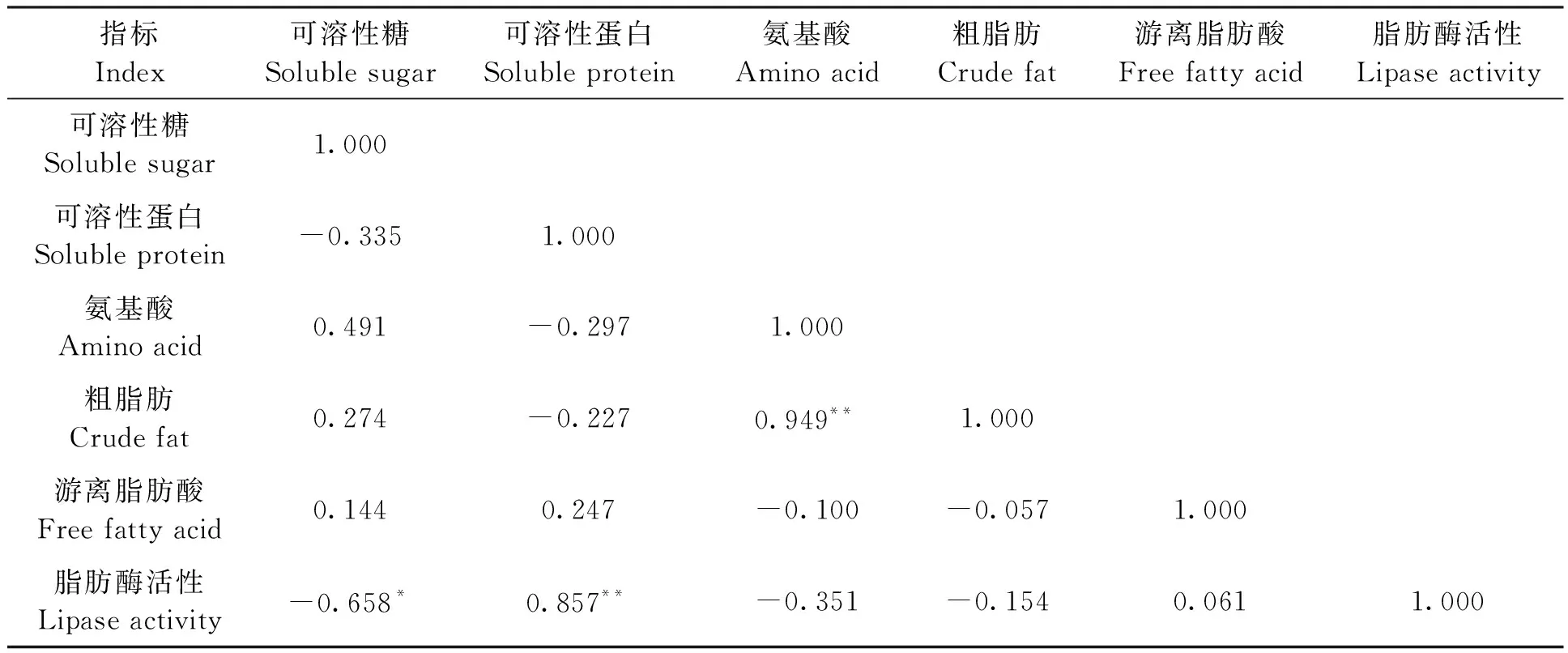
表1 芝麻種子萌發過程代謝生理指標的相關分析Table 1 Correlation analysis of physiological indexes of metabolism during germination of sesame seed
3 討論
種子萌發是植物重要的生命過程之一,可以分為吸脹、萌動、發芽和成苗4個階段,涉及一系列的生理和形態變化,并受環境條件的影響[17]。本研究在25 ℃暗培養條件下,通過形態學、水分吸收和代謝生理指標的檢測,明確了芝麻種子吸脹、萌動、胚根快速伸長、胚芽快速生長和完成發芽的時間點,并對芝麻種子的萌發低謝機理進一步研究。芝麻種子吸脹時間的確定為芝麻生產中浸種處理提供理論參考,即25 ℃時浸種時間不宜超過6 h,溫度較高時可適當縮減浸種時間[10-11]。
碳水化合物(糖)、蛋白質和脂肪是芝麻種子中的主要物質成分,占種子質量的90%以上[18],也是參與芝麻種子萌發代謝的主要物質。芝麻種子中糖類主要是葡萄糖、果糖、蔗糖、車前糖等可溶性糖[18-19]。種子萌發過程中,可溶性糖中的葡萄糖等直接被代謝利用,蔗糖等則被快速分解為小分子物質被利用,為種子萌發和胚生長提供能量[17,20]。本研究中芝麻萌發過程中,可溶性糖含量在0~48 h之間不斷減少,在完成發芽的72 h時則表現為增加,這與韓亞飛等[10]的研究一致,也與其他脂肪類種子的研究結果[20-21]相同。種子萌發72 h時,可溶性糖含量增高,可能是脂肪代謝產生糖類的原因[17]。因此,糖類是芝麻種子萌發過程中最先被代謝利用的物質,為萌發和生長提供能量,而脂肪代謝明顯遲于糖代謝。
蛋白質在種子萌發過程中具有不可替代的作用,其代謝產物氨基酸參與新蛋白合成或為呼吸氧化提供碳架[17,22-23]。種子中的蛋白質主要以貯藏蛋白形式存在,其參與代謝的第一步是轉化為可溶性蛋白。本研究中芝麻種子萌發過程中,吸脹(6 h)時可溶性蛋白較干種子有所降低,隨后隨著萌發進程的推進不斷上升,變化趨勢與花生[24]和刺萼龍葵[25]等的研究結果相似。可能是種子吸脹引起細胞膜損傷導致可溶性蛋白透過膜外滲,從而表現為可溶性蛋白含量降低,隨后貯藏蛋白不斷實現水溶化,導致可溶性蛋白含量增加[20]。
作為種子中高能量貯藏物,脂肪在酶的作用下分解出的脂肪酸轉化為糖供植物利用[20]。本研究結果顯示,芝麻種子萌發過程中粗脂肪含量在24 h前基本沒有變化,48 h后才開始不斷下降。游離脂肪酸含量在吸脹(6 h)時大幅度下降,這可能與吸脹引起的細胞膜損傷導致的游離脂肪酸外滲有關[17,20],隨后不斷上升,于48 h達到最高值后再次下降。隨后脂肪分解,游離脂肪酸含量不斷上升,在48 h之后游離脂肪酸進入代謝過程,其含量再次下降。綜合來看,粗脂肪含量24 h之后開始減少,游離脂肪酸含量48 h達到高峰后快速降低,脂肪酶活性不斷增強,以及可溶性糖含量于72 h再次升高,可以認為芝麻種子萌發過程中脂肪的代謝發生較晚。
綜上所述,在芝麻種子萌發過程中,糖類物質最先參與代謝,其后是蛋白質,最后才是脂肪,這與其他油料作物種子萌發代謝順序基本一致[17,20]。本研究結果闡述了芝麻種子萌發過程中物質代謝先后順序,為萌發過程中相關基因表達及代謝組學等進一步研究提供了理論參考。
