Cordyceps fumosorosea和Beauveria bassiana來(lái)源的堿性蛋白酶在畢赤酵母中的異源表達(dá)與性質(zhì)測(cè)定
2020-03-16 02:06:20任雅馨羅會(huì)穎姚斌王國(guó)增涂濤
中國(guó)農(nóng)業(yè)科技導(dǎo)報(bào) 2020年11期
任雅馨, 羅會(huì)穎, 姚斌 , 王國(guó)增, 涂濤*
(1.中國(guó)農(nóng)業(yè)科學(xué)院飼料研究所, 農(nóng)業(yè)農(nóng)村部飼料生物技術(shù)重點(diǎn)實(shí)驗(yàn)室, 北京 100081; 2.福州大學(xué)生物科學(xué)與工程學(xué)院, 福州 350116)
蛋白酶是特異水解蛋白質(zhì)中肽鍵的一類(lèi)酶的總稱(chēng),其來(lái)源豐富,廣泛存在于動(dòng)物、植物及微生物中[1]。蛋白酶的結(jié)構(gòu)性質(zhì)多樣,種類(lèi)繁多,有多種分類(lèi)方式,根據(jù)其作用的不同pH范圍,可將其分為酸性蛋白酶、中性蛋白酶和堿性蛋白酶[2,3]。堿性蛋白酶主要在堿性環(huán)境中發(fā)揮酶活性,最適pH主要集中在9.0~11.0,最適作用溫度范圍較廣[2-3]。目前已發(fā)現(xiàn)的堿性蛋白酶主要為絲氨酸蛋白酶,催化三聯(lián)體中起核心作用的殘基為絲氨酸[4]。堿性蛋白酶的分子量大小主要分布在15~34 kD,少部分可達(dá)45甚至82 kD,也有分子量為8 kD的堿性蛋白酶[5-6]。其等電點(diǎn)一般較高,大多分布在pH 8.0~9.0[7]。盡管堿性蛋白酶的來(lái)源廣泛,但微生物來(lái)源的堿性蛋白酶以其周期短、成本低、限制因素少等優(yōu)點(diǎn)成為目前生產(chǎn)工藝最為成熟的一類(lèi)堿性蛋白酶。目前,能產(chǎn)堿性蛋白酶的微生物有很多,大部分是芽孢桿菌,如枯草桿菌(Bacillussubtilis)、短小芽孢桿菌(Bacilluspumilus)、地衣芽孢桿菌(Bacilluslicheniformis)等;有關(guān)真菌來(lái)源的堿性蛋白的報(bào)道也在逐年增多,以酵母和絲狀真菌為主,如棕曲霉(Aspergillusochraceus)、嗜熱圓酵母(Torulathermophila)等;部分放線(xiàn)菌也可以分泌堿性蛋白酶,如灰色鏈霉菌(Streptomycesgriseus)、費(fèi)氏鏈霉菌(Streptomycesfradiae)等[8]。
作為應(yīng)用最為廣泛的工業(yè)酶制劑之一,蛋白酶約占酶制劑總銷(xiāo)售量的60%,其中堿性蛋白酶的占比近40%[2-3],發(fā)揮著巨大的工業(yè)應(yīng)用價(jià)值。洗滌劑中添加堿性蛋白酶能提高洗滌劑的清潔能力,加快對(duì)蛋白類(lèi)污漬的分解,作為堿性蛋白酶最主要的應(yīng)用市場(chǎng),洗滌劑等相關(guān)工業(yè)蛋白酶需求量洗滌劑等相關(guān)工業(yè)的堿性蛋白酶需求量約占其全球需求總量的2/3[2-3]。在食品行業(yè)中,堿性蛋白酶被應(yīng)用于大豆蛋白的水解,以產(chǎn)生高營(yíng)養(yǎng)價(jià)值的水解產(chǎn)物[9];此外,乳制品及肉制品加工也需要堿性蛋白酶的作用[10-11]。在飼料行業(yè)中,部分堿性蛋白酶具有水解角蛋白的能力,可以降解包括羽毛在內(nèi)的富含角蛋白卻難以降解的蛋白資源,應(yīng)用于飼料的生產(chǎn)中[12]。在制革過(guò)程中,可以利用堿性蛋白酶處理皮革原料,不僅可以有效的避免環(huán)境污染和安全問(wèn)題,而且可以大大提高脫毛速度,增加皮革面積,保留皮革原本的顏色,達(dá)到更好的脫毛效果[13]。在醫(yī)藥領(lǐng)域,堿性蛋白酶也發(fā)揮著一定的作用,其被對(duì)于燒傷、傷口化膿、血栓溶解具有一定的療效[14-15]。此外,堿性蛋白酶在污水處理、廢棄銀回收、絲綢脫膠等方面也有所應(yīng)用。巨大的應(yīng)用前景使得針對(duì)堿性蛋白酶的研究十分必要。
直接利用野生菌株發(fā)酵生產(chǎn)堿性蛋白酶存在產(chǎn)酶水平低、性質(zhì)不穩(wěn)定、分離純化麻煩等缺點(diǎn)。隨著基因工程技術(shù)的發(fā)展,建立了許多優(yōu)質(zhì)的表達(dá)系統(tǒng)以實(shí)現(xiàn)目標(biāo)蛋白的高效表達(dá)。目前,已成功構(gòu)建的蛋白表達(dá)系統(tǒng)有大腸桿菌表達(dá)系統(tǒng)、酵母表達(dá)系統(tǒng)、枯草芽孢桿菌表達(dá)系統(tǒng)、昆蟲(chóng)表達(dá)系統(tǒng)和哺乳動(dòng)物表達(dá)系統(tǒng)等。其中,畢赤酵母表達(dá)系統(tǒng)以其表達(dá)局限性小、產(chǎn)量高、可控性強(qiáng)、產(chǎn)品投入少、產(chǎn)物易分離及純化效果好等特點(diǎn)成為備受青睞的表達(dá)系統(tǒng),尤其是對(duì)于真菌來(lái)源的目的蛋白,其具有完善的翻譯后修飾功能。目前,已有堿性蛋白酶實(shí)現(xiàn)了在畢赤酵母中的表達(dá),如來(lái)源于A(yíng)spergillusoryzae的Alp[16]及來(lái)源于A(yíng)spergillussojae的AP[17]等。本研究對(duì)來(lái)源于真菌Cordycepsfumosorosea和Beauveriabassiana的堿性蛋白酶基因pa1及pa2成功實(shí)現(xiàn)了在畢赤酵母GS115中的高效表達(dá),對(duì)重組蛋白的酶學(xué)性質(zhì)進(jìn)行了系統(tǒng)的研究,以期挖掘具有潛在工業(yè)應(yīng)用前景的優(yōu)質(zhì)堿性蛋白酶。
1 材料與方法
1.1 材料
1.1.1菌株與質(zhì)粒 質(zhì)粒pEASY-T3和菌株EscherichiacoliTrans-T1購(gòu)自北京全式金公司,分別用作克隆載體和宿主;質(zhì)粒pPIC9和畢赤酵母PichiapastorisGS115 為本實(shí)驗(yàn)室保存,分別用作表達(dá)載體和宿主。
1.1.2培養(yǎng)基 LB培養(yǎng)基:酵母浸提物5 g·L-1、胰蛋白胨10 g·L-1、NaCl 10 g·L-1,固體培養(yǎng)基添加15 g·L-1瓊脂粉。
MD固體培養(yǎng)基:葡萄糖20 g·L-1、瓊脂糖20 g·L-1、YNB 13.4 g·L-1、生物素0.004 g·L-1。
YPD培養(yǎng)基:葡萄糖20 g·L-1,蛋白胨20 g·L-1,酵母提取物10 g·L-1。
BMGY培養(yǎng)基:酵母提取物10 g·L-1、蛋白胨20 g·L-1、1%甘油(體積分?jǐn)?shù))、YNB 13.4 g·L-1、生物素0.004 g·L-1。
BMMY培養(yǎng)基:酵母提取物10 g·L-1、蛋白胨20 g·L-1、0.5%無(wú)水甲醇(體積分?jǐn)?shù))、YNB 13.4 g·L-1、生物素0.004 g·L-1。
牛奶雙層固體培養(yǎng)基:上層包含酵母提取物10 g·L-1、蛋白胨20 g·L-1、0.5%無(wú)水甲醇(體積分?jǐn)?shù))、YNB 13.4 g·L-1、生物素0.004 g·L-1、瓊脂粉20 g·L-1;下層包含脫脂奶粉80 g·L-1、瓊脂粉20 g·L-1。
1.1.3試劑 底物酪蛋白(casein sodium salt from bovine milk)購(gòu)自Sigma公司;福林酚試劑購(gòu)自上海荔達(dá)生物科技有限公司;質(zhì)粒小提中量試劑盒、蛋白分子量Marker(M221)購(gòu)自天根生化科技有限公司;DNA純化試劑盒購(gòu)自O(shè)MEGA公司;限制性?xún)?nèi)切酶EcoRⅠ、NotⅠ、BglⅡ購(gòu)自TaKaRa公司;Assembly Kit試劑盒2×FastPfuPCR SuperMix購(gòu)自北京全式金公司;2×TaqPCR StarMix、核酸分子量Maker(MD113)購(gòu)自GenStar康潤(rùn)生物公司;其他試劑均為國(guó)產(chǎn)分析純。
1.1.4儀器 純水儀(美國(guó)Millipore)、立式高速冷凍離心機(jī)與臺(tái)式高速冷凍離心機(jī)(日本HIMAC)、移液器(德國(guó)Eppendorf)、凝膠成像系統(tǒng),PCR儀與電轉(zhuǎn)儀(美國(guó)Bio-Rad)、膜包(德國(guó)Vivascience)、AKTA avant蛋白純化儀與陽(yáng)離子交換柱HiTrap SP HP 5 mL(美國(guó)GE)、酶標(biāo)儀(美國(guó)Thermo),其他均為國(guó)產(chǎn)儀器。
1.2 方法
1.2.1菌株產(chǎn)酶能力的鑒定 蛋白酶的酶活(U)定義為:標(biāo)準(zhǔn)條件下(pH 8.5,60 ℃,反應(yīng)20 min),每分鐘釋放1 μg酪氨酸所需的酶量為一個(gè)酶活單位。
電擊轉(zhuǎn)化后獲得攜帶堿性蛋白酶基因的轉(zhuǎn)化子,轉(zhuǎn)接到牛奶雙層平板上,30 ℃條件下培養(yǎng)72 h,根據(jù)轉(zhuǎn)化子周?chē)欠癯霈F(xiàn)牛奶水解圈及出圈大小,判斷轉(zhuǎn)化子是否表達(dá)蛋白酶活性,并衡量其活性高低。
1.2.2堿性蛋白酶的酶活測(cè)定 本文以福林酚法測(cè)定蛋白酶活性,以酪蛋白為底物:500 μL底物(1% ,質(zhì)量體積分?jǐn)?shù))和500 μL適當(dāng)稀釋倍數(shù)的酶液,在60 ℃反應(yīng)20 min后加入1 mL 0.4 mol·L-1三氯乙酸溶液終止反應(yīng)。將終止后的反應(yīng)體系12 000 r·min-1離心3 min后,取上清進(jìn)行顯色。顯色反應(yīng)體系:2.5 mL 0.4 mol·L-1碳酸鈉溶液,500 μL上清和 500 μL 福林酚試劑,40 ℃反應(yīng)20 min,待其冷卻至室溫后讀取其波長(zhǎng)680 nm的吸光值,計(jì)算酶活力。所有反應(yīng)均設(shè)有三個(gè)平行和空白對(duì)照。
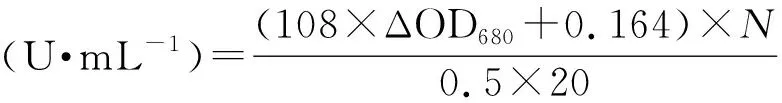
式中,ΔOD680=實(shí)驗(yàn)組OD680-空白對(duì)照OD680;N為配液的稀釋倍數(shù)。
1.2.3編碼堿性蛋白酶基因的獲取及其表達(dá)載體的構(gòu)建與鑒定 本研究中異源表達(dá)的兩條堿性蛋白酶基因分別來(lái)源于真菌Cordycepsfumosorosea和Beauveriabassiana,根據(jù)Genbank中公布的蛋白序列(ANV82734.1和AHA43398.1),基于畢赤酵母的偏好性對(duì)其二者進(jìn)行密碼子優(yōu)化,并送至深圳華大基因有限公司進(jìn)行合成,分別命名為pa1和pa2。
合成獲得的基因通過(guò)同源重組的方法連接在pUC57載體上。首先,通過(guò)PCR方法借助特異性引物向目的基因中引入同源區(qū)段(pa1-F:GGCTGAAGCTTACGTAGAATTCGCTCCAGTTGTT-GAACCAGCTCCT,pa1-R:CTAAGGCGAATTAAT-TCGCGGCCGCTTAAGTAGCACCGTTAAAAGCCAA-GTAGTTGACA;pa2-F:GGCTGAAGCTTACGTA-GAATTCGCTCCAGTTGTTGAACCAGCTCCA,pa2-R:CTAAGGCGAATTAATTCGCGGCCGCTTAAGT-AGCACCGTTGAAAGCCAAGTAGTTAAC,下劃線(xiàn)標(biāo)注為與載體pPIC9同源區(qū)段)。PCR反應(yīng)體系:2×FastPfuPCR SuperMix 25 μL,pUC57-X1 μL,引物 各1 μL;ddH2O 22 μL(X表示基因編號(hào))。PCR反應(yīng)條件:94 ℃ 5 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 90 s,30個(gè)循環(huán);72 ℃ 10 min。同時(shí),利用限制性?xún)?nèi)切酶EcoRⅠ、NotⅠ對(duì)表達(dá)載體pPIC9進(jìn)行酶切,并回收PCR產(chǎn)物和酶切產(chǎn)物。回收后的表達(dá)載體pPIC9和攜帶同源區(qū)段的目的片段通過(guò)Assembly Kit試劑盒進(jìn)行連接,連接產(chǎn)物轉(zhuǎn)化E.coliTrans-T1感受態(tài),轉(zhuǎn)化菌液涂布于LB平板(含100 μg·mL-1氨芐青霉素)。 37 ℃培養(yǎng)箱過(guò)夜培養(yǎng)后,挑取轉(zhuǎn)化子進(jìn)行菌落PCR驗(yàn)證,反應(yīng)體系:2×TaqPCR StarMix 10 μL,引物各1 μL,ddH2O 8 μL,挑取轉(zhuǎn)化子加入上述反應(yīng)體系中。反應(yīng)條件同上。將陽(yáng)性轉(zhuǎn)化子送華大基因測(cè)序,正確的重組質(zhì)粒命名為pPIC9-pa1、pPIC9-pa2。
1.2.4重組堿性蛋白酶表達(dá)載體在畢赤酵母中的表達(dá)及純化 通過(guò)質(zhì)粒小提中量試劑盒提取重組pPIC9-pa1、pPIC9-pa2質(zhì)粒,利用限制性?xún)?nèi)切酶BglⅡ?qū)ζ溥M(jìn)行線(xiàn)性化處理,利用DNA純化試劑盒進(jìn)行純化。取20 μL純化后的線(xiàn)性化產(chǎn)物加入80 μL畢赤酵母感受態(tài)進(jìn)行電擊,設(shè)置Fungi檔位為PIC模式。電擊完畢后,立即加入200 μL預(yù)冷的1 mol·L-1山梨醇,懸浮菌液并轉(zhuǎn)移至滅菌的EP管中,30 ℃靜置復(fù)蘇30~60 min。將菌液涂布在MD平板上,30 ℃培養(yǎng)48 h。隨后將MD平板上生長(zhǎng)的畢赤酵母單菌落轉(zhuǎn)接到牛奶雙層平板上,30 ℃培養(yǎng)72 h,若轉(zhuǎn)化子表達(dá)蛋白酶酶活,則可水解牛奶雙層平板中所含的牛奶,形成明顯的水解圈。通過(guò)水解圈出現(xiàn)與否及其大小判斷各陽(yáng)性轉(zhuǎn)化子的酶活,挑選最早出圈且水解圈最大的轉(zhuǎn)化子進(jìn)行搖瓶發(fā)酵。
將選中的重組酵母菌株接種到30 mL YPD液體培養(yǎng)基中,30 ℃、200 r·min-1活化培養(yǎng)48 h。隨后按1%的接種量將其轉(zhuǎn)接到300 mL的BMGY培養(yǎng)基中,30 ℃、200 r·min-1培養(yǎng)48 h,后將菌體轉(zhuǎn)移到200 mL的BMMY培養(yǎng)基中,30 ℃、200 r·min-1誘導(dǎo)培養(yǎng)48 h,期間每隔24 h補(bǔ)加0.5%甲醇。發(fā)酵過(guò)程結(jié)束后,將菌液12 000 r·min-1離心10 min,收集上清,即為粗酶液。用截留分子量5 kD的膜包對(duì)粗酶液進(jìn)行濃縮,并通過(guò)透析(截留分子量:3 kD)將粗酶液置換到pH 6.5的10 mmol·L-1磷酸氫二鈉-檸檬酸緩沖液(buffer A)中實(shí)現(xiàn)脫鹽。脫鹽后的酶液通過(guò)陽(yáng)離子交換柱(HiTrap SP HP 5 mL)進(jìn)行純化,利用buffer B (在buffer A基礎(chǔ)上添加1.0 mol·L-1NaCl,并調(diào)節(jié)為pH 6.5)進(jìn)行梯度(0~1.0 mol·L-1NaCl)洗脫,收集洗脫下來(lái)的具有酶活力的組份,利用SDS-PAGE驗(yàn)證其蛋白的純度及分子量大小,并送目的條帶至中國(guó)農(nóng)業(yè)科學(xué)院蜜蜂研究所進(jìn)行液相色譜-電噴霧離子化-質(zhì)譜聯(lián)用(liquid chromatography-electrospray ionization-tandem mass spectrometry,LC-ESI-MS)鑒定。蛋白濃度通過(guò)Easy Protein Quantitative Kit蛋白定量試劑盒進(jìn)行定量。
1.2.5堿性蛋白酶的酶學(xué)性質(zhì)研究 酶學(xué)性質(zhì)研究所用酶液均達(dá)到電泳純。緩沖體系:乳酸-乳酸鈉緩沖體系(50 mmol·L-1,pH 3.0~5.0),磷酸鹽緩沖體系(200 mmol·L-1,pH5.0~8.0),硼砂-氫氧化鈉緩沖體系(100 mmol·L-1,pH 8.0~12.0)。
①最適pH及pH穩(wěn)定性。利用pH 5.0~12.0的緩沖液將純化后的PA1、PA2酶液稀釋適當(dāng)倍數(shù),在60 ℃下,與處于相應(yīng)pH的底物反應(yīng)20 min,測(cè)定其活性,以探究PA1、PA2的最適pH。
將純化后的PA1、PA2酶液在37 ℃、pH 3.0~12.0條件下預(yù)處理1 h后進(jìn)行適當(dāng)稀釋?zhuān)?0 ℃、pH 8.5條件下測(cè)定剩余酶活,以探究PA1、PA2的pH穩(wěn)定性。
②最適溫度及熱穩(wěn)定性。將純化后的PA1、PA2酶液用100 mmol·L-1,pH 8.5的硼砂-氫氧化鈉緩沖體系稀釋適當(dāng)倍數(shù),測(cè)定二者在不同溫度下(20~80 ℃)的酶活,以探究其最適溫度。
將純化后的PA1、PA2用100 mmol·L-1、pH 8.5的硼砂-氫氧化鈉緩沖體系稀釋到50 μg·mL-1,分別在50、55、60和65 ℃下進(jìn)行熱處理,處理時(shí)間分別為2、5、10、20、30和60 min,處理后的樣品置于冰上以終止酶的熱損失,冷卻后稀釋適當(dāng)倍數(shù),在60 ℃,pH 8.5條件下測(cè)定剩余酶活,以探究其熱穩(wěn)定性,以未處理酶液所測(cè)的活性為100%。
③對(duì)不同金屬離子及化學(xué)試劑的反應(yīng)。在酶促反應(yīng)體系中加入不同金屬離子及化學(xué)試劑(Na+、K+、Cu2+、Ca2+、Mg2+、Zn2+、Mn2+、Co2+、Ni2+、Fe3+、Pb2+、Cr3+、EDTA、β-巰基乙醇、SDS),終濃度為5 mmol·L-1,60 ℃、pH 8.5(100 mmol·L-1硼砂-氫氧化鈉緩沖體系)條件下反應(yīng)20 min,以探究不同金屬離子及化學(xué)試劑對(duì)酶活性的影響。以未添加金屬離子或化學(xué)試劑的反應(yīng)體系所測(cè)酶活為100%。
④對(duì)不同蛋白酶抑制劑的反應(yīng)。在酶促反應(yīng)體系中分別加入蛋白酶抑制劑E-64(半胱氨酸蛋白酶抑制劑)、phenylmethylsulfonyl fluoride(PMSF,絲氨酸蛋白酶抑制劑)、ethylenediaminetetraacetic acid(EDTA, 金屬蛋白酶抑制劑)及pepstatin A(天冬氨酸蛋白酶抑制劑),終濃度為1 mmol·L-1,60 ℃、pH 8.5(100 mmol·L-1硼砂-氫氧化鈉緩沖體系)條件下反應(yīng)20 min,以探究不同蛋白酶抑制劑對(duì)酶活性的影響。以未添加抑制劑的反應(yīng)體系所測(cè)酶活為100%。
⑤動(dòng)力學(xué)參數(shù)。分別配制濃度為0.5、0.8、1.0、1.3、1.5、2.0、2.5、5.0、8.0、10.0 mg·mL-1的酪蛋白為底物,對(duì)純化后的PA1、PA2酶液適當(dāng)稀釋與不同濃度的底物在60 ℃、pH 8.5條件下反應(yīng)10 min,測(cè)定酶活力。利用GraphPad Prism version 5.01 (La Jolla, CA)軟件計(jì)算Km及Vmax,再根據(jù)酶的理論分子量計(jì)算kcat值。
1.2.6序列分析及結(jié)構(gòu)建模 利用在線(xiàn)軟件Signal P 4.0 (http://www.cbs.dtu.dk/services/SignalP/)預(yù)測(cè)其信號(hào)肽,Motif Scan(https://myhits.isb-sib.ch/cgi-bin/motif_scan/)預(yù)測(cè)其N(xiāo)端前肽;利用NetNGlyc 1.0 Server預(yù)測(cè)N糖基化位點(diǎn);利用Vector NTI Advance 10.0 software分析PA1、PA2的蛋白理論分子量、等電點(diǎn);SWISS-MODEL (https://www.swissmodel.expasy.org/)進(jìn)行三級(jí)結(jié)構(gòu)的建模及其活性位點(diǎn)分析。
2 結(jié)果與分析
2.1 堿性蛋白酶PA1及PA2序列分析
來(lái)源于真菌Cordycepsfumosorosea的堿性蛋白酶PA1全長(zhǎng)380個(gè)氨基酸,預(yù)測(cè)其N(xiāo)端1~15個(gè)氨基酸為信號(hào)肽。根據(jù)畢赤酵母偏好性對(duì)去除信號(hào)肽外的365個(gè)氨基酸進(jìn)行密碼子優(yōu)化,獲得全長(zhǎng)為1 098 bp的基因序列(包括終止密碼子),提交至NCBI數(shù)據(jù)庫(kù),序列號(hào)為MK622909。胞外蛋白酶通常由三部分組成,信號(hào)肽、N端前肽及催化功能區(qū),少部分蛋白酶前體還具有C端前肽或C端延伸區(qū)。酶原表達(dá)至胞外后,需加工去除N端前肽(或C端前肽)才可成為有活性的成熟酶[18],預(yù)測(cè)PA1的16~100位氨基酸構(gòu)成其N(xiāo)端前肽。N-糖基化預(yù)測(cè)結(jié)果顯示PA1含有1個(gè)潛在的N-糖基化修飾位點(diǎn)Asn270。PA1成熟蛋白理論蛋白分子量為28.6 kD,等電點(diǎn)為8.82。
來(lái)源于真菌Beauveriabassiana的堿性蛋白酶PA2全長(zhǎng)為379個(gè)氨基酸,經(jīng)過(guò)序列分析,其與PA1的一致性為80.0%,前1~15位氨基酸為信號(hào)肽,對(duì)除去信號(hào)肽外的364個(gè)氨基酸進(jìn)行密碼子優(yōu)化,獲得全長(zhǎng)為1 095 bp的基因序列(包括終止密碼子),優(yōu)化后的序列提交至NCBI數(shù)據(jù)庫(kù),序列號(hào)為MK622910。PA2蛋白的16~99位氨基酸構(gòu)成其N(xiāo)端前肽,加工去除N端前肽后的成熟蛋白的理論蛋白分子量大小為28.2 kD,等電點(diǎn)為9.10。不含潛在的N-糖基化修飾位點(diǎn)。
2.2 堿性蛋白酶PA1及PA2結(jié)構(gòu)分析
PA1與已有晶體結(jié)構(gòu)的Lecanicilliumpsalliotae來(lái)源的堿性蛋白酶Ver122(PDB登錄號(hào):3F7M)一致性為82.44%。以3F7M為模板對(duì)PA1進(jìn)行同源建模(圖1)。這是一個(gè)典型的枯草桿菌類(lèi)蛋白酶的結(jié)構(gòu),由兩層α-螺旋結(jié)構(gòu)以及夾在中間的一層β-折疊結(jié)構(gòu)構(gòu)成。第五個(gè)β-折疊和第四個(gè)α-螺旋之間以及第六個(gè)β-折疊和第五個(gè)α-螺旋之間的兩段肽段形成底物結(jié)合區(qū)域,底物結(jié)合區(qū)域向內(nèi)存在三個(gè)催化位點(diǎn),分為別Asp 141、His171和Ser326。PA1成熟蛋白中共有5個(gè)Cys,其中Cys136-Cys225和Cys280-Cys351分別形成兩對(duì)二硫鍵,Cys175并未形成。此外,在PA1成熟蛋白中還存在一個(gè)保守的Ca2+結(jié)合位點(diǎn),殘基Pro277、Val279、Asp302與Ca2+形成穩(wěn)定的作用網(wǎng)絡(luò)。
與PA1一樣, PA2與已有晶體結(jié)構(gòu)的Lecanicilliumpsalliotae來(lái)源的堿性蛋白酶Ver122(PDB登錄號(hào):3F7M)的一致性最高,為79.21%。以3F7M為模板對(duì)PA1進(jìn)行同源建模(圖1)。根據(jù)同源建模結(jié)果,PA2的蛋白結(jié)構(gòu)與PA1相似,都是典型的枯草桿菌類(lèi)蛋白酶結(jié)構(gòu),底物結(jié)合位置、催化三聯(lián)體、二硫鍵及鈣離子結(jié)合位點(diǎn)的分布均十分相似。
2.3 堿性蛋白酶PA1及PA2的表達(dá)與純化
重組表達(dá)載體pPIC9-pa1及pPIC9-pa2在畢赤酵母GS115中成功實(shí)現(xiàn)異源表達(dá),根據(jù)牛奶水解圈的大小及出圈先后,從36個(gè)轉(zhuǎn)化子中篩選出酶活較高的一株進(jìn)行搖瓶發(fā)酵。搖瓶發(fā)酵獲得的粗酶液經(jīng)過(guò)截留分子量為5 kD膜包濃縮后,進(jìn)一步通過(guò)利用陽(yáng)離子交換柱進(jìn)行純化,純化后的蛋白分子量大小及其純度通過(guò)SDS-PAGE電泳的驗(yàn)證(圖2)。在25~35 kD處分別有單一的蛋白條帶,這與PA1和PA2的理論分子量大小相符。在N糖基化預(yù)測(cè)時(shí)表明蛋白PA1僅存在一個(gè)潛在糖基化位點(diǎn),PA2沒(méi)有潛在糖基化位點(diǎn),說(shuō)明該蛋白在酵母表達(dá)時(shí)的N糖基化程度低或者無(wú)N糖基化修飾。
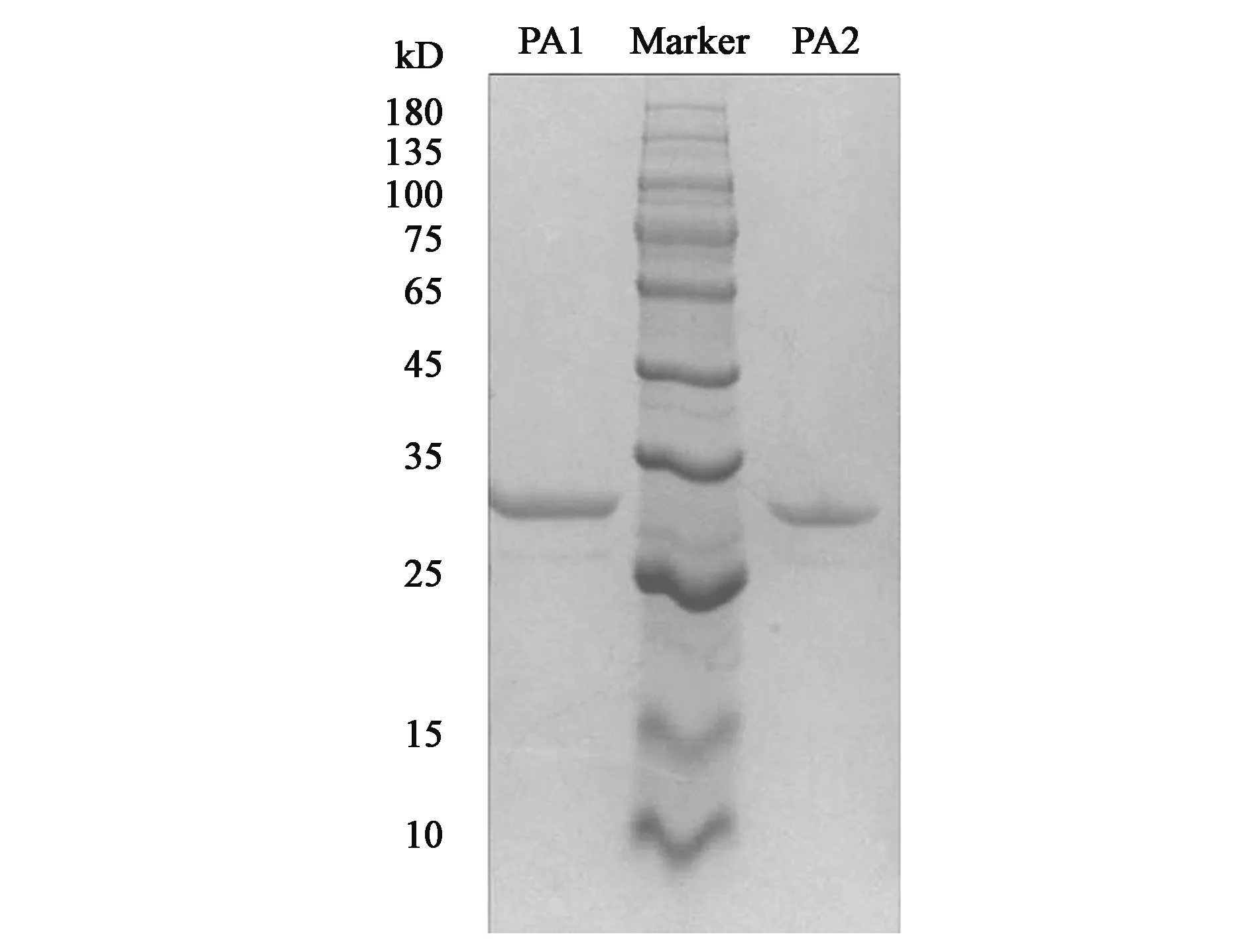
圖2 PA1及PA2純化產(chǎn)物SDS-PAGEFig.2 SDS-PAGE analysis of purified PA1 and PA2
2.4 堿性蛋白酶PA1及PA2的酶活測(cè)定
對(duì)于挑選獲得的PA1及PA2轉(zhuǎn)化子進(jìn)行搖瓶發(fā)酵,測(cè)定結(jié)果顯示,在60 ℃、pH 8.5條件下,PA1的粗酶液酶活為(218.4±2.7)U·mL-1,根據(jù)誘導(dǎo)時(shí)間,其平均生產(chǎn)力可以換算為4.55 U·mL-1·h-1。同樣條件下,PA2的粗酶液酶活為(246.5±3.5) U·mL-1,其平均生產(chǎn)力可以換算為5.14 U·mL-1·h-1。以酪蛋白為底物,純化后的PA1及PA2比活分別為(2 705.3±31.9)和(2 242±39.5)U·mg-1。
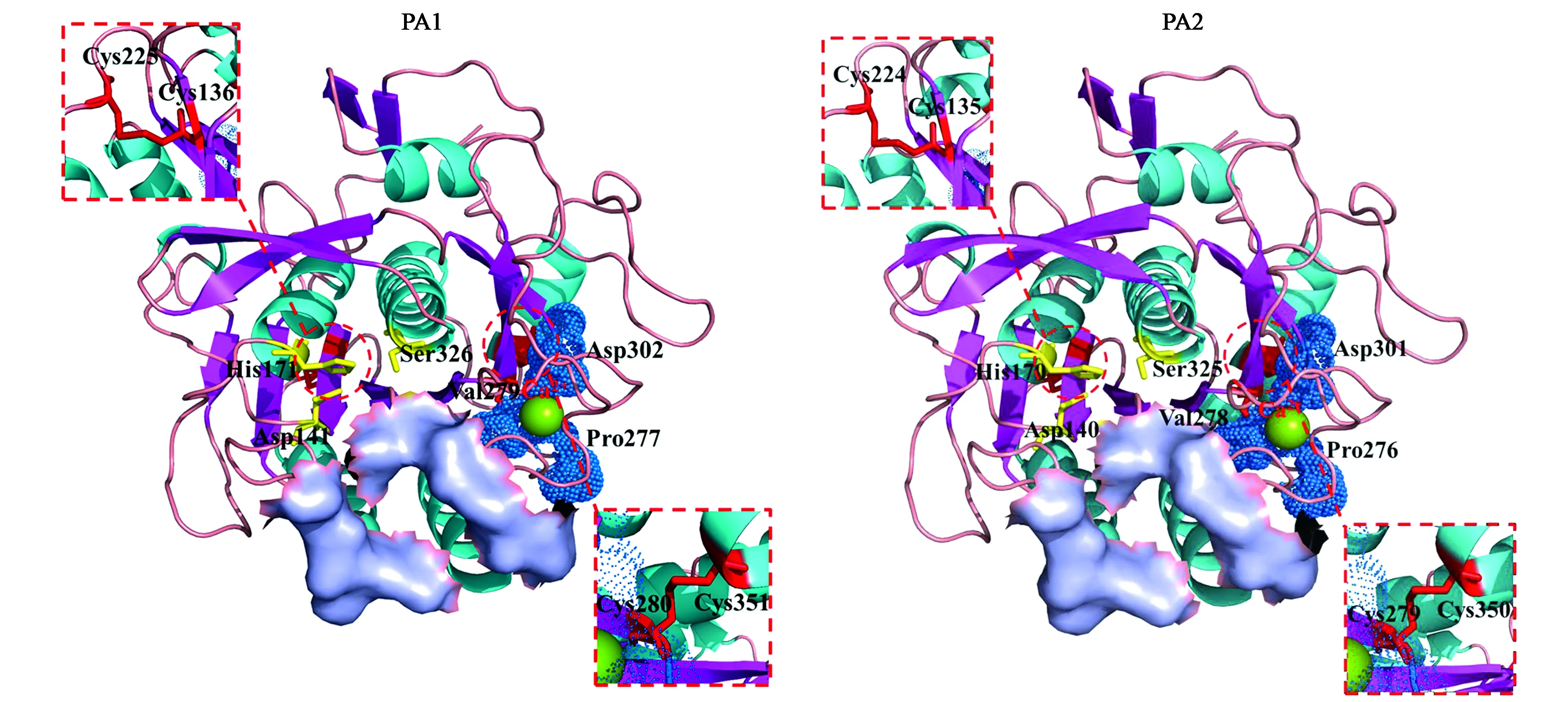
注:黃色標(biāo)注的氨基酸為催化三聯(lián)體;綠色球體為Ca2+;藍(lán)色球體為與Ca2+結(jié)合的氨基酸;紫色標(biāo)注部分為底物結(jié)合位置;紅色標(biāo)注部分為二硫鍵。Note: Yellow labeled amino acids are catalytic triads; green spheres are Ca2+ atom; blue spheres are the amino acids bound to Ca2+; purple surfaces are the substrate binding position; the sites marked in red are disulfide bonds.圖1 PA1及PA2三級(jí)結(jié)構(gòu)Fig.1 Putative modeled structure of PA1 and PA2
2.5 pH對(duì)堿性蛋白酶PA1及PA2的影響
純化后的PA1最適pH為8.5(圖3),整體嗜堿,在pH 7~10之間酶活均保持在50%以上,其中pH 8.0~9.5條件下,更是保持在90%以上。當(dāng)pH上升至10.5后,酶活明顯降低,pH 12.0條件下,基本檢測(cè)不到蛋白酶活性。盡管屬于堿性蛋白酶,PA1在偏酸條件下仍能檢測(cè)到一定活性,pH 6.0時(shí)仍保持30%以上的活性。與PA1相同,純化后PA2的最適pH也為8.5(圖3),整體嗜堿,但相較于PA1,PA2的整體作用范圍稍向酸偏移,在pH 10.0條件下,僅保持30%左右的酶活,較PA1低了30%左右,而在pH 6.0條件下,PA2保持40%以上的活性,較PA1高近10%。
PA1和PA2都具有良好的pH穩(wěn)定性(圖3),在pH 4.0~11.0條件下處理1 h后,PA1仍能保持80%以上的剩余酶活,而PA2的剩余酶活可以保持85%以上,這充分說(shuō)明PA1和PA2的pH穩(wěn)定性范圍十分寬泛。
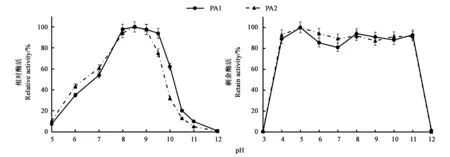
圖3 PA1及PA2最適pH和pH穩(wěn)定性Fig.3 Optimum pH and pH stabilities of PA1 and PA2
2.6 溫度對(duì)堿性蛋白酶PA1及PA2的影響
PA1的最適溫度為60 ℃(圖4),最適溫度以下,酶活性隨溫度變化趨勢(shì)較為平緩,反應(yīng)溫度在40 ℃時(shí),仍保持40%以上的酶活。當(dāng)反應(yīng)溫度高于60 ℃后,酶活性隨溫度變化趨勢(shì)相對(duì)陡峭,盡管65 ℃條件下,仍能保持近50%的酶活,但當(dāng)溫度達(dá)到70 ℃時(shí),僅保持10%左右的活性。與PA1相同,PA2的最適溫度也為60 ℃,酶活性隨溫度變化的整體趨勢(shì)與PA1相似。在目前已報(bào)道的真菌來(lái)源的堿性蛋白酶中, PA1及PA2屬于真菌來(lái)源的堿性蛋白酶中的高溫酶。
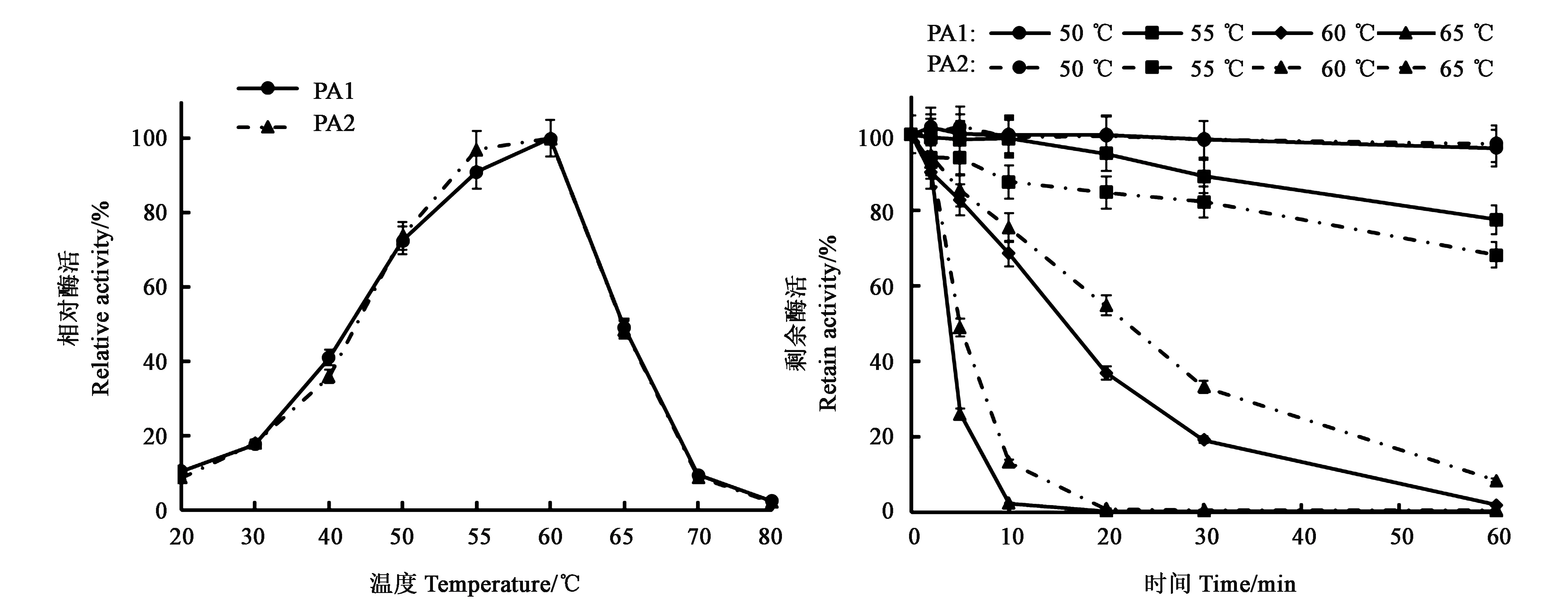
圖4 PA1及PA2最適溫度和熱穩(wěn)定性Fig.4 Optimum temperature and thermostability of PA1 and PA2
PA1的溫度穩(wěn)定性也較好(圖4),在50 ℃條件下高度穩(wěn)定,處理1 h幾乎沒(méi)有酶活損失;55 ℃處理1 h,仍保持近80%的剩余酶活;在60 ℃的條件下處理10 min后還有70%左右的剩余酶活,30 min時(shí)就僅剩20%左右的酶活;當(dāng)處理溫度達(dá)到65 ℃時(shí),酶活降低比較明顯,處理10 min后,幾乎檢測(cè)不到活性。PA2的溫度穩(wěn)定性也較好,在50 ℃條件處理1 h幾乎沒(méi)有酶活損失;與PA1相比,在相對(duì)低溫的條件下,PA2的穩(wěn)定性略低,55 ℃條件下處理1 h,PA2的剩余酶活比PA1低10%左右。但在相對(duì)高溫的條件下,PA2的穩(wěn)定性?xún)?yōu)于PA1,PA2在65 ℃條件下處理10 min后,仍能檢測(cè)到13%左右的活性。
2.7 金屬離子和化學(xué)試劑對(duì)堿性蛋白酶PA1及PA2的影響
從表1可以看出,Ni2+和Mn2+對(duì)PA1及PA2有明顯的促進(jìn)作用,Zn2+、Co2+和Cu2+對(duì)PA1及PA2的抑制作用明顯,其中Cu2+的抑制效果最為嚴(yán)重,終濃度5 mmol·L-1Cu2+存在的情況下PA1和PA2幾乎損失90%左右的酶活。其他金屬離子對(duì)PA1及PA2活性也有程度不同的作用。EDTA對(duì)PA1及PA2的酶活影響幾乎沒(méi)有影響,在表面活性劑SDS存在的情況下,PA1及PA2的酶活性仍能保持60%,還原劑β-巰基乙醇存在時(shí),PA1及PA2的酶活性仍能保持近90%,這二者作為常用的工業(yè)添加劑,PA1及PA2在其存在的情況下仍能保持不低的活性,這表明PA1及PA2具有一定的工業(yè)應(yīng)用潛力。
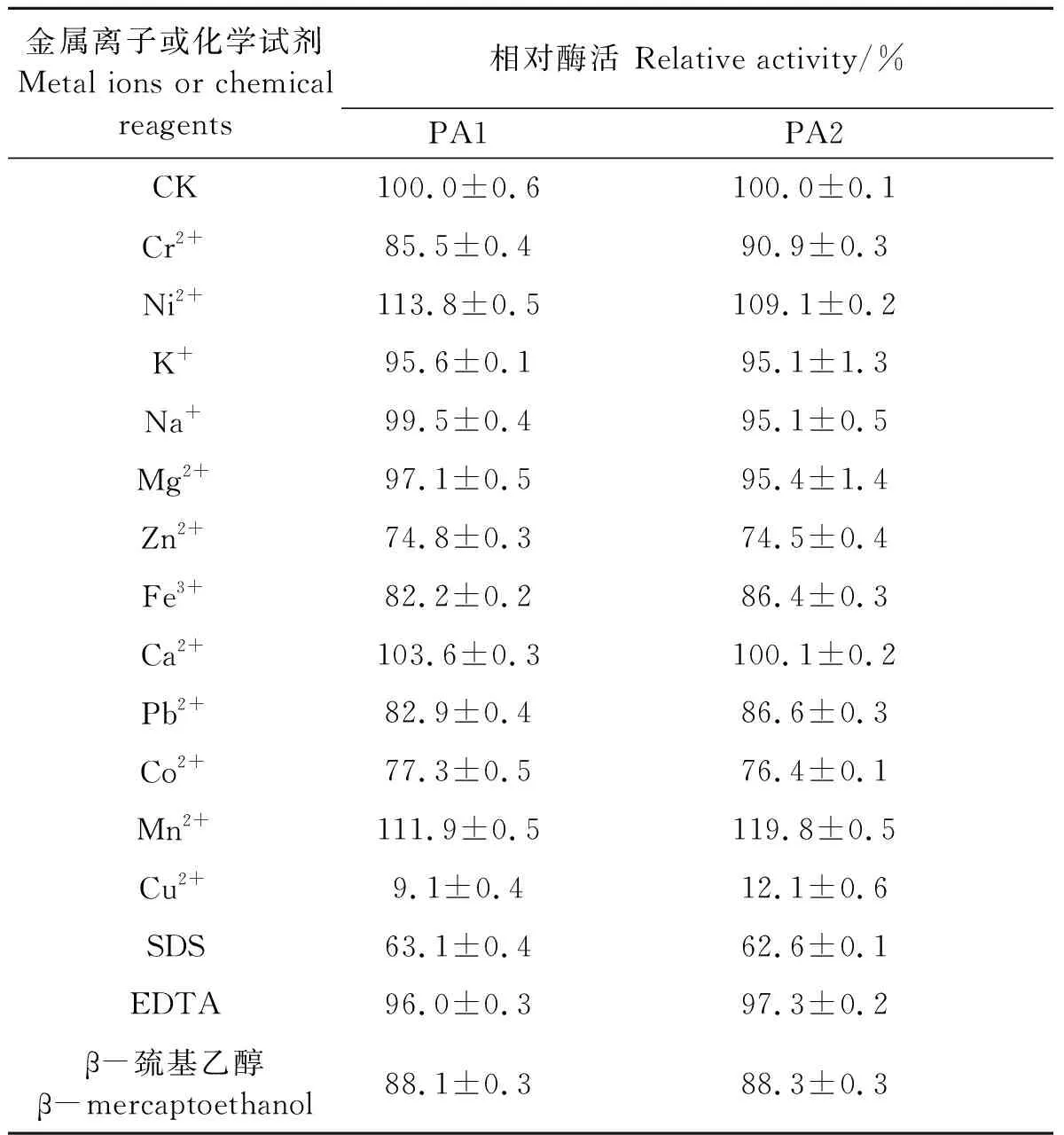
表1 金屬離子及化學(xué)試劑對(duì)PA1及PA2活力的影響Table 1 Effect of metal ions and chemical reagents on the activity of PA1 and PA2
2.8 不同蛋白酶抑制劑對(duì)堿性蛋白酶PA1及PA2的作用
如圖5所示,半胱氨酸蛋白酶抑制劑E-64、金屬蛋白酶抑制劑 EDTA及天冬氨酸蛋白酶抑制劑 Pepstatin A對(duì)PA1及PA2幾乎沒(méi)有抑制作用,而在絲氨酸蛋白酶抑制劑PMSF存在的情況下,PA1及PA2酶活性基本完全喪失,該結(jié)果進(jìn)一步證明PA1及PA2屬于絲氨酸蛋白酶。
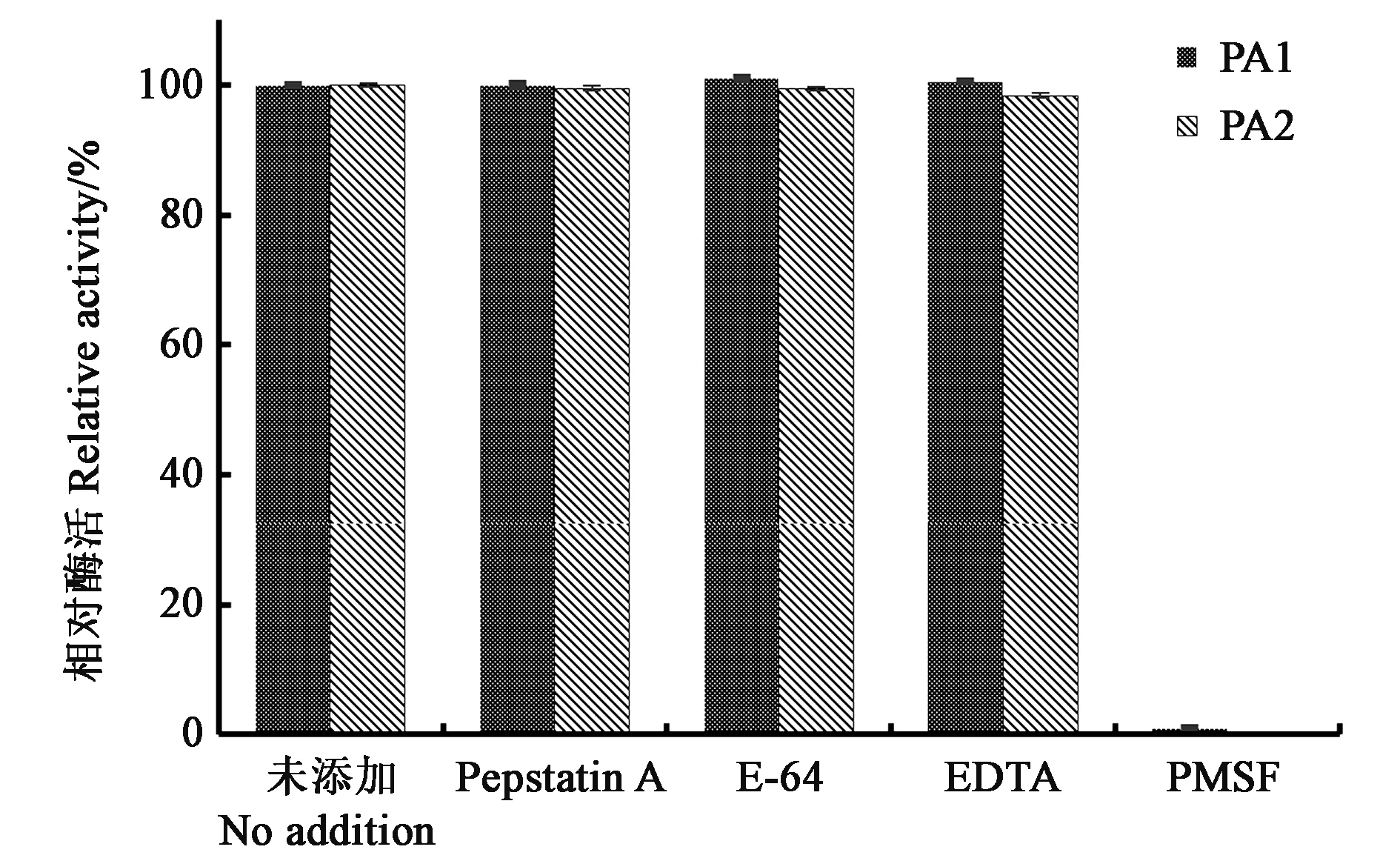
圖5 蛋白酶抑制劑對(duì)PA1及PA2活力的影響Fig.5 Effect of protease inhibitors on the activity of PA1 and PA2
2.9 堿性蛋白酶PA1及PA2的動(dòng)力學(xué)常數(shù)
以酪蛋白為底物,在60 ℃、pH 8.5的條件下,測(cè)得堿性蛋白酶PA1的Km值為(3.46±0.23) mg·mL-1,Vmax值為(4 283.0±56.4) μmol·min-1·mg-1,進(jìn)一步計(jì)算得,其kcat值為2 027.8 s-1,催化效率kcat/Km為585.6 mL·s-1·mg-1。相同條件下測(cè)定PA2的Km值為(4.39±0.58) mg·mL-1,Vmax值為(4 596.0±68.9) μmol ·min-1·mg-1,進(jìn)一步計(jì)算得,其kcat值為2 150 s-1,催化效率kcat/Km為490.3 mL·s-1·mg-1。
3 討論
本文以Parengyodontiumalbum來(lái)源的優(yōu)質(zhì)堿性蛋白酶Proteinase K的蛋白序列出發(fā),通過(guò)序列和進(jìn)化分析,獲得真菌Cordycepsfumosorosea和Beauveriabassiana來(lái)源的堿性蛋白酶基因pa1及pa2。其中pa1尚無(wú)功能驗(yàn)證相關(guān)報(bào)道,對(duì)于pa2,2014年,Borgi等[19]曾通過(guò)原酶純化手段獲得對(duì)其所表達(dá)蛋白,并確定為堿性蛋白酶,但尚未發(fā)現(xiàn)對(duì)其二者進(jìn)行異源表達(dá)及性質(zhì)研究的報(bào)道。目前,已報(bào)道的用于堿性蛋白酶異源表達(dá)的宿主菌有大腸桿菌、枯草芽孢桿菌、畢赤酵母、解脂耶氏酵母,釀酒酵母及魯氏酵母。本文利用畢赤酵母表達(dá)系統(tǒng)對(duì)pa1及pa2進(jìn)行異源表達(dá),按照畢赤酵母偏好性進(jìn)行密碼子優(yōu)化,去除了其二者的天然信號(hào)肽,與pPIC9表達(dá)載體連接,采用載體上存在α-factor信號(hào)肽引導(dǎo)實(shí)現(xiàn)跨膜轉(zhuǎn)移。重組菌株P(guān).pastorisGS115 PA1及P.pastorisGS115 PA2在搖瓶發(fā)酵水平上,PA1平均生產(chǎn)力可以換算為4.55 U·mL-1·h-1,PA2平均生產(chǎn)力可以換算為5.14 U·mL-1·h-1。二者的表達(dá)水平比大多數(shù)報(bào)道的同類(lèi)型重組菌株要高,如來(lái)源于A(yíng)spergillusoryzae的Alp在P.pastorisGS115中實(shí)現(xiàn)異源表達(dá),平均生產(chǎn)力為0.4 U·mL-1·h-1[16];來(lái)源于BacillusstearothermophilusFI的F1在P.pastorisGS115中實(shí)現(xiàn)異源表達(dá),平均生產(chǎn)力為0.6 U·mL-1·h-1[20];來(lái)源于BacilluscereusYSQ08的aprA在P.pastorisX33中實(shí)現(xiàn)異源表達(dá)[21],平均生產(chǎn)力為1.7 U·mL-1·h-1。2018年,Ke等[17]實(shí)現(xiàn)了Aspergillussojae來(lái)源的堿性蛋白酶Ap在PichiapastorisKM71中的高效表達(dá),72 h發(fā)酵后測(cè)得酶活為400.4 U·mL-1,平均生產(chǎn)力約為5.6 U·mL-1·h-1,相較于本文所報(bào)道的PA1及PA2略高,但分析誘導(dǎo)過(guò)程,其所添加的甲醇量為本實(shí)驗(yàn)的兩倍。因此,后續(xù)工作中可通過(guò)優(yōu)化P.pastorisGS115 PA1及P.pastorisGS115 PA2的發(fā)酵工藝參數(shù),進(jìn)一步提高其表達(dá)水平。
真菌來(lái)源的堿性蛋白酶的最適溫度一般較低,多數(shù)處于35到50 ℃,如來(lái)源于A(yíng)spergillusfumigatus的TKU003的最適溫度為40 ℃[22],來(lái)源于A(yíng)spergillusustus的NIOCC#20的最適溫度為45 ℃[23]。本研究中PA1及PA2的最適溫度均為60 ℃,與近年來(lái)報(bào)道的一些真菌來(lái)源的高溫堿性蛋白酶相一致,如來(lái)源于Streptomycesambofaciens.的DP2[24]和來(lái)源Engyodontiumalbum的堿性蛋白酶[25],因此認(rèn)為PA1及PA2屬于高溫堿性蛋白酶。除最適溫度較高以外,PA1及PA2也具有較好的熱穩(wěn)定性,在50 ℃條件下處理1 h幾乎沒(méi)有酶活損失,60 ℃環(huán)境中,PA1及PA2處理30 min后仍具有一定活性。多數(shù)堿性蛋白酶的熱穩(wěn)定性較差,如來(lái)源于A(yíng)spergillusclavatus的堿性蛋白酶在50 ℃條件下處理18 min,剩余50%的活性[26],來(lái)源于Beauveriabassiana的堿性蛋白酶在50 ℃條件下處理5 min,僅剩余40%左右的活性[27]。從結(jié)構(gòu)角度分析,PA1及PA2的蛋白結(jié)構(gòu)中還存在一個(gè)Ca2+結(jié)合位點(diǎn),Ca2+對(duì)大多數(shù)堿性蛋白酶的熱穩(wěn)定性有增強(qiáng)作用[28-29]。
已報(bào)道的真菌來(lái)源的堿性蛋白酶最適pH從7.5到11均有分布,如來(lái)源于Beauveriabassiana的BBP的最適pH為7.5[30],來(lái)源于Engyodontiumalbum的堿性蛋白酶最適pH為11.0[25]。PA1及PA2的最適pH為8.5,與早先報(bào)道的最適pH分布在8.0~9.5的真菌來(lái)源的堿性蛋白酶相一致,如Aspergillusparasiticus[31]、Aspergillusfumigatus[32]及Aspergillusclavatus[26]來(lái)源的堿性蛋白酶,符合堿性蛋白酶的pH活性范圍。此外,PA1及PA2還具有極好的pH穩(wěn)定性,大部分堿性蛋白酶在中性及堿性環(huán)境中穩(wěn)定,如來(lái)源于A(yíng)spergillusclavatusES1的堿性蛋白酶蛋白酶在4 ℃下處理1 h,僅在pH 8.0~9.0的環(huán)境中保持穩(wěn)定,pH 6及pH 10環(huán)境中酶活損失50%左右[33];來(lái)源于A(yíng)spergillusnidulansHA-10的堿性蛋白酶在37 ℃條件下處理1 h,其活性穩(wěn)定范圍為pH 6.0~10.0,在pH 4環(huán)境中僅保持30%左右的活性[34]。目前,也有一些真菌來(lái)源的堿性蛋白酶在酸性環(huán)境中保持一定的穩(wěn)定性,如Fusariumsp. BLB來(lái)源的堿性蛋白酶在37 ℃ 1 h的處理?xiàng)l件下,在pH 3.0~9.0的環(huán)境中保持穩(wěn)定,盡管在酸性環(huán)境中具有良好的穩(wěn)定性,但其在堿性條件下的穩(wěn)定較差,pH 11.0的環(huán)境中酶活幾乎完全損失[35]。PA1及PA2在37 ℃ 1 h的處理?xiàng)l件下,在pH 4.0~11.0的環(huán)境中均保持80%以上的活性,呈現(xiàn)出良好的穩(wěn)定性。
綜上所述,來(lái)源于真菌Cordycepsfumosorosea和Beauveriabassiana的堿性蛋白酶基因pa1和pa2與pPIC9表達(dá)載體連接,經(jīng)轉(zhuǎn)化后成功實(shí)現(xiàn)在畢赤酵母中的高效表達(dá)。對(duì)純化后的PA1及PA2的酶學(xué)性質(zhì)進(jìn)行測(cè)定,確定其二者的最適pH均為8.5,最適溫度均為60 ℃,都具有良好的pH穩(wěn)定性和溫度穩(wěn)定性,對(duì)于表面活性劑和還原劑也具有一定的耐受性,表明二者具有一定的工業(yè)應(yīng)用潛力。在后續(xù)工作中,可對(duì)二者的應(yīng)用價(jià)值展開(kāi)進(jìn)一步研究,如可將其應(yīng)用于加酶洗滌劑、羽毛降解、皮革制造、肉質(zhì)嫩化等領(lǐng)域,分析比較其各自在實(shí)際生產(chǎn)中的優(yōu)劣,更好地發(fā)揮其本身價(jià)值,豐富堿性蛋白酶資源,為工業(yè)生產(chǎn)提供更多選擇。
